 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Acetylsalicylic acid: its effects on a highly expressed phosphatase from Solanum cardiophyllum Humberto Lopez Delgado and Guillermo Carrillo-Castaneda Colegio de Postgraduados. IRGP. Montecillo, Mex. 56230. Mexico.
Code Number:BA96075
Size of Files:
Text: 25.5K
Graphics: Line drawings (gif) - 6.4K
Photographs (jpg) - 55.6K
Abstract A pH 6 phosphatase activity was identified in extracts from Solanum cardiophyllum plantlets cultured in vitro in the absence of acetylsalicylic acid (ASA) using p-nitrophenylphosphate as substrate. This activity was inhibited by both phosphate and fluoride but not by ASA added to the enzymatic reaction mixtures. There was, however, a marked difference found between the development of the plantlets grown on the ASA-containing (10^-5 M) (MBA) medium, which grew poorly on it and those which, after being exposed to ASA in a first period, were cultured in a subsequent period on the ASA-free (MB) medium. In these not only the original stems turned into vigorous plantlets during the second culture period, but also many new stems emerged. In a similar way, the activity expressed in the plantlets that had grown in the presence of ASA, whose average activity was 27 % lower than the activity of the plantlets developed on the ASA-free medium, differed from that expressed in the plantlets developed first on the MBA medium and in a consecutive period on the ASA-free medium, being the average activity detected 94 % higher than the activity of the plantlets grown on the ASA- free medium.
Key words: phosphatase activity, signal transduction, plantlets
Resumen Una actividad de fosfatasa pH 6 fue identificada en extractos de plantulas de Solanum cardiophyllum cultivadas in vitro, en ausencia de acido acetilsalicilico (ASA), usando como substrato p-nitrofenilfosfato. Esta actividad, que fue altamente expresada en la plantula, fue inhibida por fosfato y fluoruro pero no por ASA agregados a las mezclas de reaccion enzimaticas. Sin embargo, una marcada diferencia fue encontrada entre las plantulas desarrolladas en el medio suplementado con ASA 10^-5 M (MBA), las cuales dificilmente crecieron y las plantulas que se cultivaron primero en el medio MBA y posteriormente en el medio sin ASA (MB). En estas, el tallo original crecio vigorosamente en el segundo medio y varios tallos nuevos emergieron. De manera paralela, la actividad de fosfatasa expresada en las plantulas desarrolladas en presencia de ASA, que fue en promedio 27 % menor que la actividad expresada en las plantulas desarrolladas en el medio MB, fue diferente de la actividad expresada en las plantulas desarrolladas en forma consecutiva en el medio MBA y posteriormente en el MB, la cual fue en promedio 94 % mayor a la observada en las plantulas cultivadas en el medio MB.
Palabras claves: actividad de fosfatasas, transduccion de senal, plantula, PR-proteinas
Introduction The plant facing a pathogen attack induces a broad-spectrum resistance. This includes the localized accumulation of salicylic acid (SA) followed by its systemic accumulation. A set of genes, the so-called pathogenesis-related genes are then switched on and their products in turn act as molecular signals promoting the coordinate expression of the systemic acquired resistance (SAR) genes (1). These confer a nonspecific resistance (2). The concentration of SA in these plants was correlated with the level of induction of both SAR genes and resistance (3-6).
Reversible protein phosphorylation functions as a major mechanism for the control of biological processes in eukaryotic cells (7, 8). In plants, however, little information is available (9, 10). Recently, it has been demonstrated (11) that SA is probably not the movile inducing signal for the SAR response in plants, and that it is not the primary systemic signal (12).
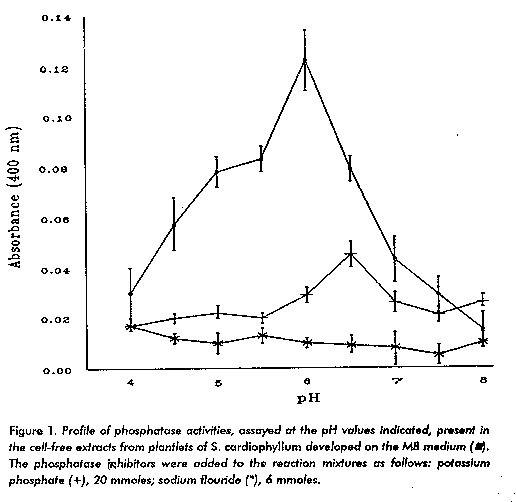
The artificial application to the plant of either SA, or ASA, its exposure to UV light or ozone (13) increases the endogenous concentration of SA to levels that account for the induction of the pathogenesis related proteins (14) and SAR response (1, 15). Some protein products of the SAR genes have activity against pathogens and therefore they are related to plant resistance (16). A rather broad spectrum of SA induced effects on biological activities of cells are known (17-24). In a previous report, the inhibitory effect of ASA on the in vitro development of S. cardiophyllum (Solanaceae) vitroplants was assessed (25). Despite the existing information, the generality of the role of SA in biological signaling is still under debate (11, 12) and the role of phosphatases in this particular case is still in the field of speculation. In view of the fact that in other systems, phosphatases are known to be involved in the control of biological processes, the experiments presented in this preliminary paper were performed to define whether or not a highly expressed phosphatase in the plant is interfered by ASA, and if that were the case, to determine the relationship between the patterns of plant growth and enzyme activity exhibited by S. cardiophyllum plantlets. Materials and Methods Culture of Axilary Buds Axilary buds of the stem from plantlets of S. cardiophyllum were cultured at 26-28 C and under a photoperiod of 16 h during periods of 30 days as follows: on the solid MBA medium, which was the Murashige and Skoog medium (26) modified as previously described (25) containing 10^-5 M ASA (condition A); on the MBA solid medium first and a subsequent period on the MB solid medium, which was the ASA- free MBA medium (condition B) and on the MB solid medium (condition C). Phosphatase Assays Plantlets (350 mg wet weight) developed from the axilary buds were ground with 2 mL of distilled water in a frozen mortar, the debris were spun down at 4 000 xg at 4 C during 10 min and the clear supernatant was immediately used for the phosphatase assays. The amount of protein in the extracts was assessed in accordance with Lowry, et al. (27). For the determination of the phosphatase activities (orthophosphoric- monoester phosphohydrolase; EC 3.1.3.2) present in the extracts, in 1 mL of the final volume, the reaction mixture contained: 0.5 mL of buffer solution (potassium acetate: acetic acid 10^-4 pH 4.0, 4.5; potassium citrate: hydrochloric acid 10^-4 M pH 5.0, 5.5, 6.0; potassium citrate: potassium hydroxide 10^-4 M pH 6.5, 7.0; tris-HCl (hidroxymethyl aminomethane): hydrochloric acid 10^-4 M pH 7.5, 8.0), 0.4 mL distilled water and 0.1 mL of a freshly prepared solution of p-nitrophenylphosphate (10 moles mL-1). Immediately after preparation the reaction mixtures were prewarmed 10 min at 30 C and afterwards 6 uL of the enzymatic extract was added (9 ug of protein approximately). The reaction mixtures were incubated at 30 C during 10 min and the reaction stopped with 3 mL of NaOH 0.1 N. The amount of p-ni- trophenol produced was determined at 400 nm. As inhibitors of the phosphatase activities 6 moles of NaF or 20 moles of potassium phosphate were used in the reaction mixture, using solutions with their pH adjusted to the pH of the buffer of the reaction mixture (28). Every enzymatic determination was performed in duplicate. Results Identification of a Highly Expressed Phosphatase Activity in S. cardiophyllum The identification of a highly expressed phosphatase activity in the enzymatic extracts of S. cardiophyllum was made screening the range of pH between 4 and 8 and using plantlets developed under the condition C. In the series of three independent experiments, the maximal phosphatase activity detected was at pH 6 (Figure 1). The phosphatase activities assayed in the presence of potassium phosphate or NaF showed that the pH 6 phosphatase activity was strongly inhibited by any of these compounds.
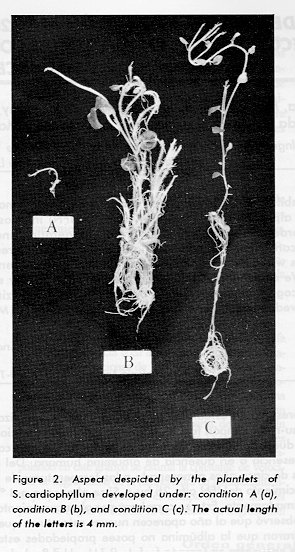
The expression of the pH 6 phosphatase activity was determined in the plantlets developed under the three experimental conditions indicated. The average enzymatic activity determined in the extracts of plantlets developed under the experimental condition C was considered as 100 %. In the extracts of plantlets developed under the condition A, the data of almost all determinations along the three series of experiments performed were the lowest observed, being the average activity 27 % lower than condition C (Table 1). Contrasting with the activity of condition A, in all the assays performed with the extracts of condition B the highest activities were detected with an average activity of 94.42 % above condition C. According to Tukey's test, two groups of means were formed. No difference between condition A and condition C mean was found; however, condition B mean differed significantly from any other condition. The analysis of variance for condition and experiment showed a highly significant effect of the condition. ASA interfered with the development of the plantlets grown under condition A (Figure 2a); however, when these plantlets were transferred to and cultured on the ASA-free medium, not only the original stems turned into vigorous plantlets but also new stems emerged (Figure 2b). The aspect of the plantlets grown on the ASA-free medium is also shown (Figure 2c).
--------------------------------------------------------------
Table 1. Determination of the pH 6 phosphatase activity in
cell-free extracts from plantlets of S.cardiophyllum
cultured during periods of 30 days on:MB medium plus 10^-5 M
ASA (condition A), MB medium plus 10^-5M ASA and a
subsequent period on MB medium (condition B), and on the
MB medium (condition C). In each experiment the assays
were performed in duplicate and the data represent the average
and percentage. Within conditions, values in the last column
followed by the same letter are not significantly different
(after Tukey's test, a = 0.05).
--------------------------------------------------------------
Plantlets mumols of p-nitrophenol produced/mg protein/hour
developed
under: Experiment 1 Experiment 2 Experiment 3 Average
--------------------------------------------------------------
Condition A 66.66 86.74 105.36 86.25 b
(74.06) (69.39) (75.31) (72.90)
Condition B 170.00 360.00 160.00 230.00 a
(188.88) (288.00) (114.36) (194.42)
Condition C 90.00 125.00 139.90 118.30 b
(100.00) (100.00) (100.00) (100.00)
--------------------------------------------------------------
The activity of the pH 6 phosphatase was assayed in the presence of 10^-3 and 10^-5 M ASA added to the reaction mixture and as is shown in Table 2, the activities detected in the extracts from plantlets grown on the ASA-free medium (condition C) were not interfered by ASA. However, in those plantlets that had been developed under condition B an average reduction of 26.68 % of the activity was observed when 10^-3 M ASA was present in the reaction mixture, in comparison with the same kind of extracts simultaneously assayed without adding ASA to the reaction mixture.
-------------------------------------------------------------- Table 2. Determination of the pH 6 phosphatase activity in the cell-free extracts of S. cardiophyllum, adding ASA to the reaction mixtures. The data, presented in percentages, are the results of three independent experiments performing each determination sixfold. -------------------------------------------------------------- Plantlets developed Concentration of ASA (M) in the mixture under 10^-3 10^-5 0 -------------------------------------------------------------- Condition B 73.32 81.32 100.00 Condition C 103.00 115.38 100.00 -------------------------------------------------------------- Discussion A phosphatase activity which is highly expressed at pH 6 in S. cardiophyllum plantlets cultured in vitro using p-nitrophenylphosphate and crude extracts was clearly identified. These experimental conditions allowed the obtainment of preliminary information in order to have a general view of the phenomenon. Gellatly, et al. (29) purified an acid phosphatase from potato tuber, which performs its maximal activity at pH 5.8 and has significant phosphotyrosine phosphatase activity. Several reports about kinases and phosphatases in plants exist (6, 29-35).
ASA by itself did not interfere with the phosphatase activity. In the series of 12 assays performed in three independent experiments, it was demonstrated that the pH 6 phosphatase activity was not interfered when 10^-5 or even 10^-3 M ASA was present in the enzymatic reaction mixture, using the extracts from plantlets that had not been exposed to ASA. These results indicated that the ASA solution by itself had not interfered with the enzymatic activity. When the plantlets had been grown in the presence of ASA, the enzymatic activity assayed under the same experimental condition suffered an average decrease of 26.68 % (Table 2) compared with the same extracts simultaneously assayed without adding ASA to the reaction mixture. The magnitude of this inhibition was particularly high taking into account that the enzymatic activity shown by the extracts from plantlets developed under the experimental condition B was 94.42 % above the average activity displayed by the extracts of condition C. These results suggest: 1) That SA and the presence of another element whose expression might be induced in the development of the cells exposed to ASA, was required in the cell to allow its interaction and to interfere with the activity of the pH 6 acid phosphatase. Chen, et al. (36) have identified in tobacco a soluble SA-binding protein (SABP) which in its native form might be a homotetramer of a 57 kDa polypeptide. They indicated that SABP in association with SA might perform altered activities. 2) That the cell growing under condition A, in order to overcome the pH 6 phosphatase activity shortage, over-produces it, and the activity determined under this condition represents that activity displayed by the free enzyme molecules. During the second period of incubation of condition B, the plantlets were developed on the ASA-free medium. Under these conditions, the concentration of endogenous SA of the cell might have fallen short enough to cause dissociation of the ternary complex. The high activity detected in this kind of extract might account for the sum of the active pH 6 phosphatase plus the enzyme molecules that reassumed their activity once dissociated (3). That the effect of ASA added at the moment of the assay on the phosphatase activity, from extracts of plantlets developed under condition B, could be explained by the remains of the ASA-induced factor present in those extracts, which allowed the formation of the ternary complex while losing the activity of a proportion of the enzyme molecules. SA interacts with specific target proteins to adjust enzymatic activities. Chen, et al. (37) have demonstrated the specific inhibition of certain catalase isozymes by SA in tobacco leaves.
The effects of ASA on plantlet development were parallel to their pH 6 phosphatase expression. The highest pH 6 phosphatase activity detected was in the plantlets developed under the experimental condition B. These cultures showed the production of several well developed stems from a single bud. This has a potential utilization in plant micropropagation. Preliminary results (unpublished data) have shown that following this procedure S. tuberosum cv Alpha stems can be obtained. Their tuber production in the greenhouse was interesting because they developed numerous aerial tubers. Contrasting with the phosphatase activity of condition B, in almost all the assays performed with the extracts of condition A, the lowest phosphatase activity was detected. The plantlets cultured under condition A showed the lowest development as well. The data of each experiment is presented (Table 1) to show the extent of the variation of the pH 6 phosphatase activity. However, the highest activity was always the expression found in the plantlets of condition B. It has been demostrated that the condition B mean differs significantly from the other conditions and the highly significant effect of the condition. Many tubers of S. cardiophyllum were utilized in the developent of this work and, in general, a single plantlet was used to prepare every enzymatic extract. We believe that the source of this variation is the natural variability of this wild species. Work in progress deals with the full characterization of the homologous enzyme in S. tuberosum which, being an important crop, suggests the potential practical applications of these procedures. Acknowledgements We thank Dr. Alfredo Carballo Quiroz and Dr. Manuel V. Ortega for a critical reading of the manuscript. References 1 Ward ER, Uknes SJ, Williams SC, Dincher SS, Wiederhold DL, Alexander DC, Ahl-Goy P, Metraux JP, Ryals JA. Coordinate gene activity in response to agents that induce systemic acquired resistance. Plant Cell 1991;3:1085-1094. 2 Ross AF. Systemic acquired resistance induced by localized virus infections in plants. Virology 1961;14:340-358. 3 Enyedi AJ, Yalpany N, Silverman P, Raskin I. Localization, conjugation and function of salicylic acid in tobacco during the hypersensitive reaction to tobacco mosaic virus. Proc Natl Acad Sci USA 1992;89:2480-2484. 4 Malamy J, Carr JP, Klessig DF, Raskin I. Salicylic acid: A likely endogenous signal in the resistance response of tobacco to viral infection. Science 1990; 250:1002-1004. 5 Metraux JP, Signer H, Ryals J, Ward E, Wyss-Benz M, Gaudin J, Raschdorf K, Schmid E, Blum W, Inveradi B. Increase in salicylic acid at the onset of the systemic acquired resistance in cucumber. Science 1990;250:1004-1006. 6 Uknes S, Winter A, Delaney T, Vernooij B, Morse A, Friedreich L, Nye G, Potter S, Ward E, Ryals J. Biological induction of systemic acquired resistance in Arabidopsis. Mol Plant-Microbe Interact 1993;6:692-698. 7 Budde RJA, Chollet R. Regulation of enzyme activity in plants by reversible phosphorylation. Physiol Plant 1988; 72:435-439. 8 Cohen SP. The structure and regulation of protein phosphatases. Annu Rev Biochem 1989;58:453-508. 9 Chen Z, Klessig DF. Identification of a soluble salicylic acid-binding protein that may function in signal transduction in the plant disease-reistance response. Proc Natl Acad Sci USA 1991;88:8179-8183. 10 Trewavas A, Gilroy S. Signal transduction in plant cells. Trends in Genet 1991;7:356-361. 11 Vernooij B, Friedrich L, Morse A, Reist R, Kilditz-Jawhar R, Ward E, Uknes S, Kessmann H, Ryals J. Salicylic acid is not the translocated signal responsible for inducing systemic acquired resistance but is required in signal transduction. Plant Cell 1994;6:959-965. 12 Rasmussen JB, Hammerschmidt R, Zook MN. Systemic induction of salicylic acid accumulation in cucumber after inoculation with Pseudomonas syrigae pv syringae. Plant Physiol 1991;97:1342-1347. 13 Yalpany N, Enyedi AJ, Leon J, Raskin I. Ultraviolet light and ozone stimulate accumulation of salicylic acid, pathogenesis- related proteins and virus resistance in tobacco. Planta 1994; 193:372-376. 14 Jung JC, Fritig B, Hahne G. Sunflower Helianthus annus L. pathogenesis-related proteins. Induction by aspirin (acetylsalicylic acid) and characterization. Plant Physiol 1993;101:873-880. 15 White RF Acetylsalicylic acid (aspirin) induces resistance to tobacco mosaic virus in tobacco. Virology 1979; 99:410-412. 16 Mauch F, Mauch-Mani B, Boyer T. Antifungal hydrolases in pea tissue. II. Inhibition of fungal growth by combinations of chitinase and beta-1,3- glucanase. Plant Physiol 1988;88:936- 942. 17 Cohen SP, Levy SB, Foulds JF, Rosner JL. Salicylate induction of antibiotic resistance in Escherichia coli: Activation of the mar-operon and a mar- independent pathway. J Bacteriol 1993; 175:7856-7862. 18 Jain A, Srivastava HS. Effect of salicylic acid on nitrate reductase activity in maize seedlings. Physiol Plant 1981; 51:339-342. 19 Jones AM. Surprising signals in plant cells. Science 1994;263:183-184. 20 Kim SR, Kim Y, An G. Identification of methyl jasmonate and salicylic acid response elements from nopaline synthase (nos) promoter. Plant Physiol 1993;103:97-103. 21 Kopp E, Ghosh S. Inhibition of NF-kB by sodium salicylate and aspirin. Science 1994;265:956-959. 22 Leslie CA, Romani RJ. Salicylic acid: a new inhibitor of ethylene biosynthesis. Plant Cell Rep 1986;5:144-146. 23 Murphy TM, Raskin I, Enyedi AJ. Plasma membrane effects of salicylic acid treatment on cultured rose cells. Envir Exp Bot 1993;33:267-272. 24 Raskin I. Role of salicylic acid in plants. Ann Rev Plant Physiol Plant Mol Biol 1992;43:439-463. 25 Lopez DH, Lopez PMC, Villalobos AVM, Larque-Saavedra A. Efecto del acido salicilico en el crecimiento de yemas de Solanum cardiophyllum cultivadas in vitro. Agrociencia serie Fitociencia 1990;1:145-159. 26 Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 1962;15:473-497. 27 Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem 1951;193:265-275. 28 Carrillo-Castaneda G, Ortega MV. Effect of inorganic phosphate upon Salmonella typhimurium phosphatase activities: non-repressible alkaline phosphatase and non- inhibited acid phosphatase. Biochim Biophys Acta 1967; 146:535-543. 29 Gellatly KS, Moorhead GBG, Duff SMG, Lefebvre DD, Plaxton WC. Purification and characteriation of potato tuber acid phosphatase having significant phosphotyrosine phosphatase activity. Plant Physiol 1994;106:223-232. 30 Arino J, Perez-Callejon E, Cunillera N, Camps M, Posas F, Ferrer A. Protein phosphatases in higher plants: Multiplicity of type 2A phosphatases in Arabidopsis thaliana. Plant Molec Biol 1993;21:475-485. 31 Casamayor A, Perez-Callejon E, Pujol G, Arino J, Ferrer A. Molecular characterization of a fourth isoform of the catalitic subunit of protein phosphatase 2A from Arabidopsis thaliana. Plant Molec Biol 1994;26:523- 528. 32 Drobak BJ. Plant phosphoinositides and intracellular signaling. Plant Physiol 1993;102:705-709. 33 Schlatter U, Wagner E, Greppin H, Bonzon M. Adenylate kinase in tobacco cell cultures. I. Separation and localization of different activities. Plant Physiol Biochem 1993;31:815-825. 34 Schlattner U, Wagner E, Greppin H, Bonzon M. Adenilate kinase in tobacco cell cultures. II. Variability and regulation of isoform activity patterns in different cell lines. J Plant Physiol 1994;144:400-409. 35 Smith MJH, Dawkins PD. Salicylate and enzymes. J Pharm Pharmac 1971; 23:729-744. 36 Chen Z, Silva H, Klessig DF. Purification and characterization of a soluble salicylic acid-binding protein from tobacco. Proc Natl Acad Sci USA 1993a; 88:8179-8184. 37 Chen Z, Silva H, Klessig DF. Active oxygen species in the induction of plant systemic acquired resistance by salicylic acid. Science 1993b;262:1883-1886. Copyright 1996 Elfinos Scientiae The following images related to this document are available:Photo images[ba96075b.jpg]Line drawing images[ba96075a.gif] |
| |||||||||

{kind=link}
{kind=link}