 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
VALIDATION OF RADIOACTIVE METHODS IN THE QUALITY CONTROL OF DNA RESTRICTION ENZYMES Elder Pupo, Enrique Perez, Luis E Trujillo, Frank Miranda, Ernesto Gonzalez and Jose Brito Molecular Biology Department. Quality Control Division. Center for Genetic Engineering and Biotechnology, P.O. Box. 6162, Havana City, Cuba.
Code Number:BA96078
Size of Files:
Text: 21.0K
Graphics:Line drawings (gif) - 5.1K
AbstractIn this paper two radioactive substrates obtained from lambda DNA digested with the restriction enzyme Hpa II were evaluated for the detection of 5'--> 3', 3'--> 5' single and double stranded-DNA dependent exonuclease and phosphatase activities found in DNA restriction and modifying enzyme preparations. A cloning simulation assay was performed using the same conditions established for the radioactive assay taking into account enzyme units and pmols of DNA ends used as substrate. As a result, it was found that for degradation of the radioactive DNA substrate per enzyme unit below 0.5 %, the false positives in the cloning simulation assay became less than 5 %. Finally, the use of the radiolabeled [gamma ^32P] ATP lambda Hpa II DNA substrate to detect 5'--> 3' single stranded-DNA dependent exonuclease and phosphatase contaminating activities is described at certain critical steps of the purification process of the restriction enzyme Kpn I. Key words: restriction/modifying enzymes, radiolabeled DNA, exonucleases, phosphatases Resumen En este trabajo se evaluaron dos sustratos radioactivos obtenidos a partir de ADN del fago lambda digerido con la restrictasa Hpa II, para la deteccion de exonucleasas con actividad 5'--> 3' dependiente de ADN de simple cadena, 3'-->5'dependiente de ADN doble cadena y fosfatasas presentes en preparaciones de enzimas de restriccion o modificacion del ADN. Fue realizada una simulacion de clonacion o ensayo de seleccion por blancas y azules donde se utilizaron las mismas condiciones empleadas para llevar a cabo el ensayo radioactivo en cuanto a cantidad de ADN (picomoles de extremos) empleado como sustrato y cantidades de enzimas ensayadas. Como resultado, se observo que para degradaciones del sustrato radiactivo menores al 0,5 % por unidad de enzima, el numero de falsos positivos encontrados en el experimento de clonacion se redujo a valores por debajo del 5 %. Finalmente, son mostrados algunos resultados obtenidos en la deteccion de exonucleasas 5'--> 3' dependientes de ADN simple cadena y fosfatasas en algunos pasos criticos en el proceso de purificacion de la enzima de restriccion Kpn I utilizando el sustrato [gamma ^32P] ATP lambda Hpa II. Palabras claves: enzimas de restriccion/modificacion, exonucleasas, fosfatasas Introduction DNA restriction and modifying enzymes are produced by different strains of microorganism and isolated to be used in molecular biology techniques. These isolation and purification procedures will ensure the elimination of non-specific activities such as exonucleases or phosphatases that could be present in a final preparation of these enzymes. Different quality control assays for the evaluation of the level of contaminants in final preparations of DNA restriction and modifying enzymes, have been developed by several companies involved in the production of these biological reagents.
The complete digestion of lambda DNA with the enzyme to be tested and long incubation periods at the selected temperature allow the detection of alterations in the expected DNA pattern bands due to different exonuclease activities, but phosphatase activities can not be detected using this method. On the other hand, the complete digestion of lambda DNA, subsequent ligation with T4 DNA ligase and recutting with the same enzyme, is a subjective functional assay (1) because the visualisation of the expected DNA patterns in each step (cut, ligation, recut) is carried out in agarose gels stained with ethidium bromide. Furthermore, the presence of low levels of contaminants in the enzyme preparations can not be detected using these methods and it is very difficult to give a quantitative result. The detection of phophatases and exonucleases using radiolabeled DNA substrates in radioactive detection assays, have shown to be more sensitive than those described above. In addition, a quantitative result in terms of substrate degradation percentage per enzyme unit as a measure of the level of contaminants in the enzyme preparations can be drawn. These assays are easy to perform and reproduce.
Although the results obtained with radioactive substrates have good detection limits for different contaminants, it is very difficult to determine the implications of this degradation percentage per enzyme unit in a true cloning experiment, or to know which is the highest limit of degradation percentage per enzyme unit allowing a successful cloning experience. Little information regarding these findings has been found in the literature.
All these uncertainties would be overcome by testing enzyme preparations in a cloning simulation as that of the blue/white cloning assay (1, 2). This assay is very rigorous for the measurement of contaminants and it can ensure a successful performance of the tested enzyme preparations in a cloning experiment.
In this report the behavior of two DNA substrates radiolabeled in both 5' and 3' ends using [gamma ^32P] ATP or [8-^3H] dGTP and [5-^3H] dCTP, to detect phosphatases and 5'--> 3', 3'--> 5' exonucleases activities dependent on single or double stranded-DNA substrates is evaluated in the final preparations and during the manufacturing process of restriction and modifying enzymes. In the search for a relationship between radioactive and functional methods the blue/white cloning assay is performed using the same conditions established for the radioactive assay (pmols of ends of DNA substrate and enzyme units) to test different final enzyme preparations produced in our laboratory and the same preparations obtained from a leader manufacturer as a quality reference. Finally, the applications of the radiolabeled [gamma ^32P] ATP lambda Hpa II DNA substrate is described for the detection of phosphatases and 5'--> 3' single stranded-DNA dependent exonucleases at certain critical steps of the production process of the restriction endonuclease Kpn I. Materials and Methods Enzymes and chemicals: X-Gal (5-bromo-4-chloro-3- indolyl-alpha- D-galactoside), IPTG (isopropil-thio-beta- D-galactoside) were obtained from Sigma. Restriction enzymes and lambda DNA used in this trial were produced at CIGB (Cuba) and also obtained from Promega and New England Biolabs. Inc. Exonuclease III, alkaline phosphatase and Mung bean nuclease were purchased from Promega and New England Biolabs Inc. [gamma ^32P] ATP, [8-^3H] dGTP and [5-^3H] dCTP were obtained from Amersham.
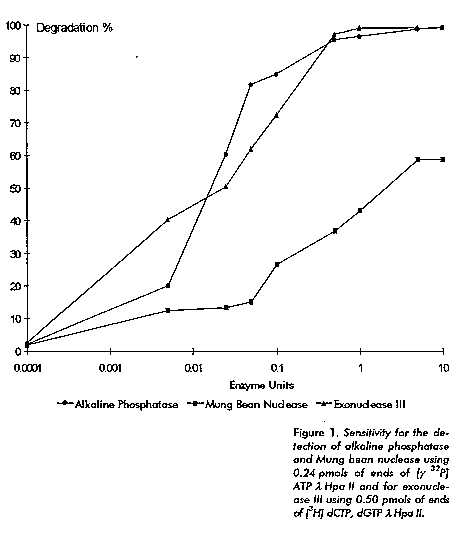
The alkaline phosphatase (CIP) treatment of the DNA substrates (lambda Hpa II) and the labeling at the 5' end with [gamma ^32P] ATP and 3' end with [8-^3H] dGTP or [5-^3H] dCTP were carried out using current methods (3, 4). The purification of radiolabeled DNA substrates to separate incorporated from unincorporated radiactivity was done using gel filtration in Nicks columns (3). PEI cellulose thin layer chromatography (5) was employed to determine the incorporation percentage of [gamma ^32P] ATP and [8-^3H] dGTP or [5-^3H] dCTP. Radiolabeled DNA substrates specific activities and incorporation percentage were calculated according to Amersham (1994/95) (6). Determination of the sensitivity of the radioactive method using radiolabeled substrates was carried out by mixing 0.24 pmols of [gamma ^32P] ATP lambda Hpa II or 0.5 pmols of [8-^3H] dGTP and [5-^3H] dCTP lambda Hpa II, 2 uL of the optimum reaction buffer 10x, and 0.0001; 0.001; 0.01; 0.1; 1 and 10 units of a commercial double or single stranded-DNA dependent exonuclease (exonuclease III or Mung bean nucleases) or phosphatase (alkaline phosphatase) respectively for each reaction tube. Distilled and sterilized water was added to complete 20 uL of the final reaction volume. Reactions were incubated 30 min at 37 C. After this time, 3 uL of the reaction were loaded and developed in a thin layer chromatography (5) using a PEI-cellulose thin layer plate. The percentage of radioactivity released from the ends of the radiolabeled DNA substrate is plotted as a function of the amount of restriction or modifying enzyme added. Assays to detect exonucleases and phosphatases contaminating activities in the restriction and modifying enzymes preparations were done by mixing in one eppendorf tube 0.24 pmols of [gamma ^32P] ATP lambda Hpa II or 0.5 pmols of [8-^3H] dGTP or [5-^3H] dCTP lambda Hpa II, 2 ul of the enzyme reaction buffer 10x, 1 uL of the enzyme dilution and water to complete the reaction volume to 20 uL. This reaction was incubated 30 min at 37 C. Results were observed as described above. Blue/white cloning assay was done using pUC19 DNA and competent cells (7) produced at CIGB (Cuba). Results and Discussion The evaluation of the sensitivity of the radioactive method for the detection of exonucleases and phosphatases using commercial preparations is displayed in Figure 1.
These radioactive substrates were very sensitive to the action of alkaline phosphatase, Mung bean nuclease and exonuclease III. In fact, a complete substrates degradation was obtained by the addition of less than one unit of both exonuclease III and alkaline phosphatase and more than 50 % for S units of Mung bean nuclease, (Figure 1). Four final preparations of the restriction enzymes Sal I, EcoR I, Cla I and Bgl II obtained in our laboratory (*) and the same preparations from a leader manufacturer (**) were assayed for the presence of phosphatases and exonucleases contaminating activities, and the degradation percentage per enzyme unit (5, 6) was calculated for each preparation (Table 1). It can be observed that the values of the degradation percentage per enzyme unit of the enzyme preparations obtained in our laboratory were similar to those obtained from a leader manufacturer, displaying for both cases the radiolabeled DNA substrate degradation percentages per enzyme unit below 0.5 % (after background subtraction), except for the Cla I preparation produced in our laboratory that displayed the highest degradation percentage per enzyme unit, 1.7 %. The same enzyme preparations and the same conditions were used in the radioactive assay (pmols of DNA and enzyme units) to carry out a blue/white cloning assay or cloning simulation assay, to learn the relationship between the results obtained in the radioactive assays and in a functional experience. This blue/white cloning assay is a functional test for contaminant detection based on restriction digestion of sites in the multiple cloning regions of the pUC19 vector which contains a sequence encoding the lac alpha peptide. Non recombinant pUC19 vectors produce a functional alpha peptide which complements the product of the lac ZdeltaM15 gene producing active beta galactosidase. Bacterial colonies from strains harboring the lac ZdeltaM15 gene on an episome F' (JM109, XL1-Blue, NM522) become blue when they are transformed with an undisrupted pUC19 vector and plated on an indicator medium containing X-gal and IPTG. If the reading frame of the lac alpha peptide is disrupted either by insertion of a foreign DNA fragment or deletion of vector sequences due to exonucleases or phosphatase activities, complementation does not occur and the bacterial colonies remain white or occasionally pale blue. Thus, from the percentage of white colonies, a measure of the level of contaminants can be obtained. The results of this cloning simulation is shown in the right column of Table 1. The percentage of white colonies (false positives) found in the plate corresponding to the ligated vector DNA increased in the same way as the values of the degradation percentage per enzyme unit obtained in the radioactive assay.
-----------------------------------------------------------
Table 1. Degradation percentage per enzyme unit of the
radiolabeled DNA substrate and the percentage of white
colonies found in the plate corresponding to the ligated
vector obtained in the blue/white cloning assay using enzyme
preparations produced in our laboratory (*) and purchased from
a leader manufacturer (**). For radioactive assay, values
were obtained after the estimation of the mean of three different
trials. For blue/white cloning, assay values were also
obtained after the estimation of the mean of three different
trials in which more than 300 colonies were analyzed.
Coefficient of variation (CV) was less than 10 % for both
assays.
--------------------------------------------------------------
Enzymes Radiolabeled DNA White colonies in the plate
degradation per enzyme corresponding to the
unit (%) DNA vector (%)
--------------------------------------------------------------
Cla I (**) 0.55 +/- 0.15 2.80 +/- 0.10
Cla I (*) 1.70 +/- 0.15 11.50 +/- 0.15
EcoRI (**) 0.30 +/- 0.19 1.50 +/- 0.15
EcoRI (*) 0.45 +/- 0.20 3.30 +/- 0.12
Sal I (**) 0.33 +/- 0.15 2.90 +/- 0.15
Sal I (*) 0.44 +/- 0.17 3.10 +/- 0.10
Bgl II (**) 0.48 +/- 0.20 2.10 +/- 0.10
Bgl II (*) 0.32 +/- 0.19 2.70 +/- 0.12
--------------------------------------------------------------
The ratio between the results obtained for Enzymes produced in our laboratory (*) and Enzymes produced by a leader manufacturer (**) displayed in Table 2, brought about very similar values for both, radioactive and functional methods, indicating a coincidence in the level of contaminants determined by these two different methods.
--------------------------------------------------------------
Table 2. Values of the ratio, enzymes produced in our
laboratory (*) / enzymes produced by a leader manufacturer
(**), for both, radioactive and functional methods. These
values were obtained using the data displayed in Table 1.
--------------------------------------------------------------
Ratio Radioactive Blue/White
assays cloning assay
--------------------------------------------------------------
Cla I (*) / Cla I (**) 3.09 4.01
EcoRI (*) / EcoRI (**) 1.50* 2.20
Sal I (*) / SAL I (**) 1.30 1.06
Bgl II (*) / BGL II (**) 1.08 1.28
--------------------------------------------------------------
From these trials it is concluded that for all enzyme preparations studied in this report, degradation percentages per enzyme unit of the radiolabeled DNA substrate below 0.5 guarantee a low percentage of white colonies in the plate corresponding to the ligated vector DNA using a biological trial as that of the blue/white cloning assay. Therefore, this value of degradation percentage is assumed to be a quality criterion for every enzyme preparation. Finally, the [gamma ^32P] ATP lambda Hpa II is used in the "in process" quality control for the detection of 5 3' single stranded-DNA dependent exonucleases as well as phosphatase activities.
Samples from certain critical purification steps were assayed for the presence of single stranded-DNA dependent exonucleases and phosphatase activities. Particularly, in the production process of four enzyme preparations: Kpn I, T4 DNA ligase, Sal I and Bgl II. All the eluted fractions from the salt gradients from each chromatography step were tested and only those displaying the lowest degradation percentage of the gamma ^32P lambda Hpa II per enzyme unit were pooled; in order to reach the end of the process with a final preparation having a degradation percentage per enzyme unit below 0.5. The efficiency of each separation step could also be evaluated by determining its capacity to separate exonucleases and phosphatases from restrictase activities (results not shown). This "in process" quality control proved to be very sensitive and effective. As a result, all final enzyme preparations were free of such contaminants. Table 3 shows some of the results corresponding to the evaluation of the degradation percentage of the [gamma ^32P] ATP lambda Hpa II in different critical purification steps from the restriction endonuclease Kpn I production process. This process includes cell disruption and two chromatographic steps using P-11 and Q-sepharose ionic exchanger resins.
--------------------------------------------------------------
Table 3. Degradation percentage per enzyme unit of the
[gamma ^32P] ATP lambda Hpa II in different steps of the
purification
procedure. These values were obtained after the
determination of the enzymatic activity of Kpn I in each
purification step.
--------------------------------------------------------------
Purification steps Degradation percentage of the [gamma
^32P] ATP lambda Hpa II per enzyme unit
--------------------------------------------------------------
Crude extract applied to the P-11 100.0 %
Pass from P-11 chromatography 66.3 %
Pooled fractions applied to the
Q-sepharose chromatography. 31.6 %
Pass from Q-sepharose chromatography 33.0 %
Final pooled fractions 0.35 %
--------------------------------------------------------------
This table also shows that the level of contaminants detected using this radioactive substrate, decreased throughout the purification process. It is very significant that the greatest level of contaminants are in the step of the P-11 chromatography while the Kpn I restrictase activity was found to be bound completely to the P-11 chromatographic resin, a fact that accounts for the high separation efficiency of this chromatography step. In view of the results obtained in this study, it is concluded that radioactive assays for contaminant detection are very sensitive and convenient to perform; they can be used for both, quality control of final enzyme preparations or for the detection of contaminants throughout the production process. Functional methods also exhibit good sensitivity for contaminant detection but are time consuming and become more expensive when they are introduced in a production process control. Results obtained with radioactive and blue/white cloning assays have shown a correspondence for all of the enzyme preparations assayed. References 1 Hung L, Murray EE, Murray W, Bandziulis R, Lowwery R, Williams R, Nobble R. A blue/white cloning assay for quality control of DNA restriction and modifying enzymes. Promega notes 1991;41:1-4. 2. Murray E, Singer K, Cash K, Williams R. Cloning -qualified blunt end restriction enzymes: Causes and cures. Promega notes 1993;33:11-13. 3. Sambrook J, Fritsh EF, Maniatis T. Molecular cloning. A Laboratory manual. Cold Spring Harbor, Laboratory. Press, New York, USA 1989. 4. Protocols and application guide from Promega. Nucleic acid detection, purification and labeling. USA 1993:123-153. 5 Life Technologies. Gibco BRL Catalogue. Enzymes for Molecular Biology 1993/94 Chapter 6-1. 6 Amersham Life Science Catalogue Data for adiochemical users: England 1993/94;36-48. 7 Hanahan D. Studies on transformation of E. coli with plasmids. J Mol Biol 1983;166:557-580. Copyright 1996 Elfos Scientiae The following images related to this document are available:Line drawing images[ba96078a.gif] |
| |||||||||

{kind=link}