 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
CELLOBIOSE DEHYDROGENASE - AN UNSOLVED PROBLEM IN FUNGAL WOOD DEGRADATION Goran Pettersson,^1 Manuel Raices^2 and Gunnar Henriksson^1
^1 Department of Biochemistry, University of Uppsala, P. O.
Box 576, 751 23 Uppsala, Sweden.
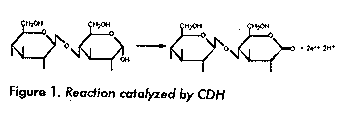
CDH binds strongly and specifically to cellulose (7, 8). No binding was discovered to chitin, starch, or to the wood components xylan and mannan. The binding was not affected by NaCl, but was inhibited by ethylene glycol and stimulated by ammonium sulphate (5). Thus hydrophobic interaction and/or charge transfer probably play an important role in the binding. The bound protein is still active and therefore the cellulose binding appears to be separated from the catalytic site (7, 8). It has been suggested that the cellulose binding is due to a cellulose binding domain similar to the ones of fungal cellulases. However, no such domain is found in the amino acid sequence of CDH (9) and the cellulose binding mechanism of this enzyme also differs from the one of the fungal cellulase CBH 1 (5). The shape of CDH and its fragments have been determined by SAXS by Lehrner and Zipper. This molecule is long (180 A^o) and thin with a "head" consisting of the FAD fragment and a "tail" corresponding to the heme fragment (10). A cDNA of CDH has been cloned and sequenced. The 5 end was obtained by PCR amplification. The cDNA contains 2310 translated bases excluding the poly (A) tail. The deduced mature protein contains 770 amino acid residues, procceded by a 18 residues long signal peptide. Data from protein characterization, i.e. sequencing, amino acid composition and molecular weight determinations are in good agreement with these finding. The regions of amino acid sequence corresponding to the heme and FAD domains were identified as well as the nucleotide binding motif. No homologous sequence were found for the heme domain, however the FAD domain appears to be related to a family of oxidoreductases including Aspergillus niger Glucose oxidase. The homologous sequences seems to be restricted to a flavin binding domain (5, 9). Although rather much is known about CDH, its function is still an open question. There are three suggestions that are popular for the moment. First CDH may work as a generator of Fe^2+ in a Fenton's reaction (11) (Figure 2). The created hydroxyl radical should then cause damage to various wood components and thereby stimulate enzymatic degradation. We have demonstrated that CDH in presence of hydrogen peroxide and complexed Fe (III)-ions can degrade cellulose, hemicellulose and lignin (12). Other suggestions are that CDH cooperates with the lignolytic enzymes lignin peroxidase and manganese peroxidase. In the case of lignin peroxidase the function should be to inhibit repolymerization of phenoxy radicals on naked cellulose (6, 13). Cooperation with manganese peroxidase should be based on the fact that CDH can solubilise insoluble Mn (IV) and provide cellobionic acid as chelating agent for Manganese ions (14). CDH can indeed work in both this ways, but the fact that CDH is produced by fungi that lacks these peroxidases talks however against this suggestion (5). A third attractive hypothesis is that CDH is the first member in an electron transport chain with the function to capture the easily available reducing energy of cellobiose, which is the major product of cellulose degradation (15). It is however not likely that this could happen without the involvement of a membrane system. In eukaryotic organisms such systems are located in mitochondria. Thus, such an electron transport chain must involve a transmembrane transport. Alternatively, an ATP generating system could exist in the cytoplasmic membrane. References 1 Perera IK, et al. Rapid Comm Mass Spec 1990;4:285- 289. 2 Ayers A, et al. Eur J Biochem 1978; 90:171-181. 3 Morpeth F. Biochemical J 1985; 228: 557-564. 4 Ander P. FEMS Microbiol Rev 1994; 13:297-312.
5 Henriksson G. Ph D Thesis, University of Uppsala Sweden 1995. 6 Henriksson G, et al. Biochem Biophys Acta 1993;1114:184-190. 7 Henriksson G, et al. Eur J Biochem 1991;196:101- 106. 8 Renganathan V, et al. Appl Microbiol Technol 1990;32:609-613. 9 Raices M, et al. FEBS Letters 1995. 10 Lehrner D, et al. Biochem Biophys Acta (Submitted) 1995. 11 Kremer S, Wood P. Eur J Biochem 1992;208:809-815. 12 Henriksson G, et al. Appl Microbiol Technol 1995;42:790-796. 13 Ander P, et al. J Biotechnol 1990; 13:189-198. 14 Roy BP, et al. J Biol Chem 1994; 269 (31):19745- 19750. 15 Eriksson KE, et al. Enz Microbiol Technol 1993;15:1002-1008. Copyright 1996 Elfos Scientiae The following images related to this document are available:Line drawing images[ba96088a.gif] |
| |||||||||

{kind=link}