 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Expression of Bacillus thuringiensis delta-Endotoxin Genes in Transgenic Plants Gustavo A de la Riva^1 and Michael J Adang^2
^1 Division de Plantas. Centro de Ingenieria Genetica y
Biotecnologia. P.O. Box 6162, C.P. 10600, Ciudad de La Habana,
Cuba.
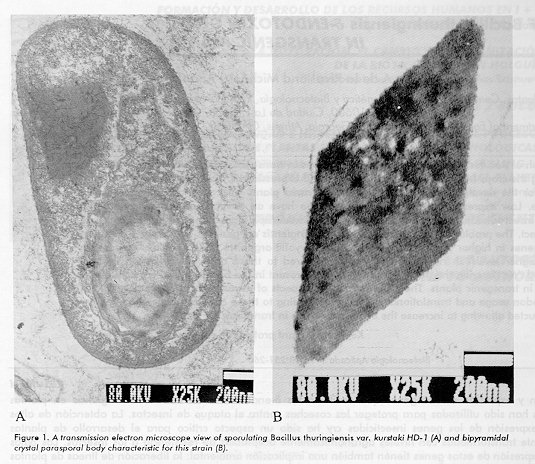
ABSTRACT The cloning and expression of Bacillus thuringiensis delta-endotoxin genes in transgenic plants have been used with the objective of protecting the crops from insect attack. The increased expression of the insecticidal cry genes in plants has been critical for the development of genetically transformed plants with agronomically acceptable levels of insect resistance. Low expression levels of such genes also have an environmental implication: the release of low expressing insect-tolerant transgenic plants may result in the rapid appearance of resistance to the Cry toxin in the target insect. The problem of the expression of B. thuringiensis cry genes is due to the expression of bacterial prokaryotic genes in higher plants or in any other eukaryotic organism. Fully modified genes can express up to 100-fold higher levels of the insecticidal toxin compared to those obtained when a wild-type bacterial gene is expressed. We describe the most important aspects present in the bacterial wild-type cry genes affecting their expression in transgenic plants. The analysis includes aspects of transcriptional regulation, mRNA stability, preferences in codon usage and translational efficiency. According to these considerations, modified cry genes have been reconstructed allowing to increase the expression levels in transgenic plants. Key words: plant protection, cry genes, modified genes RESUMEN La clonacion y expresion de los genes que codifican las delta- endotoxinas de Bacillus thuringiensis en plantas transgenicas han sido utilizadas para proteger las cosechas contra el ataque de insectos. La obtencion de altos niveles de expresion de los genes insecticidas cry ha sido un aspecto critico para el desarrollo de plantas geneticamente transformadas con niveles agronomicamente aceptables de resistencia a los insectos. Los bajos niveles de expresion de estos genes tienen tambien una implicacion ambiental: la liberacion de lineas de plantas transgenicas con bajos niveles de expresion pero tolerantes en determinado grado al ataque de insectos puede resultar en una rapida aparicion de insecto- resistencia a la toxina Cry por parte de la especie blanco. El problema de los bajos niveles de toxina radica en que los genes cry de B. thuringiensis, bacterianos y prokaryotas son expresados en plantas superiores u otros organismos eukaryotas. Genes totalmente modificados de acuerdo a las caracteristicas de los genes nucleares de plantas han permitido obtener niveles de expresion de hasta 100 veces superiores comparando con sus homologos salvajes. Nosotros describimos los aspectos mas importantes presentes en los genes bacterianos cry de B. thuringiensis y que afectan su expresion en las plantas transgenicas. Este analisis comprende los aspectos de regulacion transcripcional, estabilidad del mRNA, uso de codones y eficiencia traduccional. De acuerdo con estas consideraciones se han reconstruido genes cry modificados que permiten aumentar niveles de expresion en plantas transgenicas. Palabras claves: proteccion de planta, genes cry, genes modificados Introduction Bacillus thuringiensis is a Gram- positive spore-forming soil bacterium widely used in agriculture as a biological pesticide. The entomocidal activity resides mainly in a parasporal protein inclusion body or crystal that is produced during sporulation (Figure 1). That crystal is composed by one or more delta-endotoxins. Because these toxins combine high potency for target insects with safety for other animals, they are valuable tools for pest insect control. The mode of action is still a matter of research; but in general, B. thuringiensis inclusion bodies dissolve in the larval midgut under alkaline conditions and proteolytic cleavage, releasing one or more insecticidal monomers. These monomers bind specifically in the insect midgut brush border to the binding proteins also called receptors. The insertion of the active toxin into the membrane of the midgut epithelial cells disturbs the osmotic balance by generating pores in the cell membrane, leading to cell lysis (1, 2). Lysis of the midgut cells and its perforation lead to paralysis and death of the insect (3). B. thuringiensis is capable of disrupting the potassium flux created by proton pumps in the midgut globet cells. Insecticidal specificity is related to the specificity of the binding of a toxin to midgut receptors, but the molecular basis of toxicity is the ability to form an ion channel.
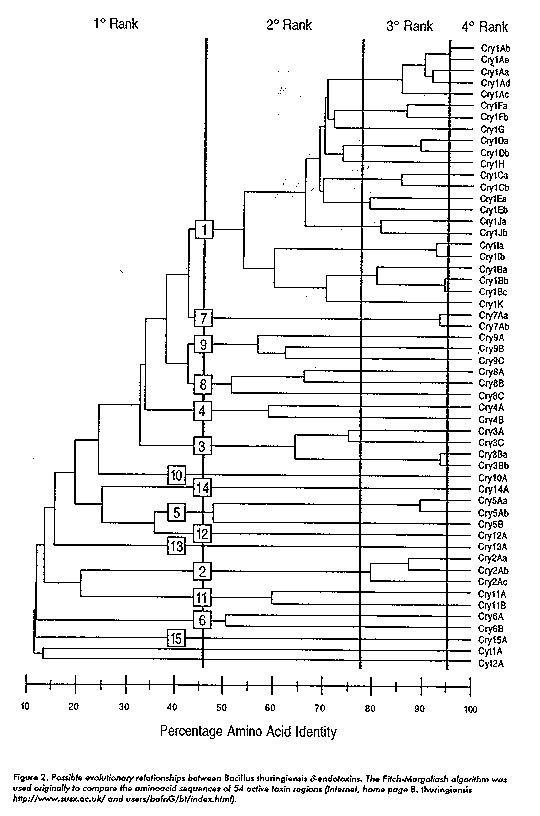
Most strains are active against certain larvae of lepidopteran species but other isolates show toxicity against dipteran, coleopteran and nematodes (Figure 2) (4). For more than three decades, formulations of B. thuringiensis have been used as biological pesticides to control agricultural pest and insect vectors of a variety of human and animal diseases. delta-endotoxins are encoded by cry genes which have been extensively studied and expressed in a wide number of prokaryotic organisms (Escherichia coli, Bacillus subtilis, Pseudomonas fluorescens) and plants. The transfer and expression of such genes in plants has provided a potentially powerful alternative for the protection of crops against insect damage. The expression of bacterial genes such as the genes cry implies the use of plant regulatory sequences as well as the modification of the gene's coding region according to the characteristics present in other plant genes.
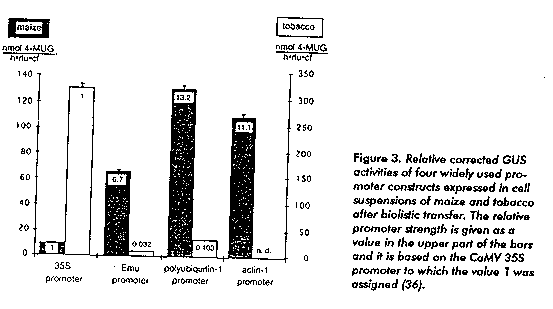
We present some of the most important aspects involved in the expression of bacterial genes in transgenic plants. Particular emphasis was made in the structure and composition of the coding region of B. thuringiensis cry genes and their decisive role in the low yields of the recombinant proteins when they are expressed in transformed plants. Plant Genetic Engineering It has become possible to introduce and express foreign genes in plant cells. The first record on transgenic tobacco plants expressing foreign genes goes back to the past decade (5, 6). Since that outstanding moment in plant sciences, a great progress has been obtained in tissue culture, plant transformation and plant molecular biology. At first, most genetically engineered plants were transformed by Agrobacterium-mediated transformations (6). Agrobacterium tumefaciens, a plant pathogen causing tumorous crowngalls on infected dicotyledonous plants, does not infect monocotyledonous plants; for this reason, many economically important plants such as cereals, remained inaccessible for genetic engineering manipulation for a long time. For those cases alternative direct transformation methods have been developed (7), such as polyethyleneglycol- mediated transfer (8), microinjection (9), protoplast and intact cell electroporation (10-14) and particle bombardment or gene gun technology (15). The gene gun technology and the electroporation of intact cells allow the direct delivery of DNA into plant cells obviating the difficulties in the regeneration of transgenic plants from transformed protoplasts. However, significant parts of the regenerated plants are not wholly transformed but form mosaic transgenic plants. To avoid this problem, the development of high efficiency Agrobacterium-mediated transformations of monocotyledonous plants is necessary. Some results in Agrobacterium-mediated transformations of monocotyledonous plants have been reported (16-19) and the most important groups increase the efforts in this direction. The generation of transgenic plants with resistance or tolerance to insect and pest attack is one of the most extensively explored fields in plant genetic engineering. The advantages of the biodegradability, selectivity and total safety for man and the environment have led to the use of BT- based products. Despite the distinct advantages of BT-based insecticides, certain reasons limit their use. These include ineffective field performance, mainly because of the toxin susceptibility to the environmental conditions and ultraviolet radiation from sunlight and, in some cases, the difficulty of the toxin for reaching the target pest inside the stem or leaves (stem borer, leave mining). Recombinant DNA technology has the potential to complement the efforts of plant breeders. The cloning and expression of cry genes in E. coli (19-24) and B. subtilis (22, 25) allow the study of the structure, molecular organization of individual cry genes and the entomocidal activity of the individual recombinant delta-endotoxins. Transgenic plants that are resistant or tolerant to insect attacks have been produced in tomato, tobacco, potato, cotton (26-32), maize (33) and more recently in sugarcane (34). Expression of Bacterial Genes in Plants During the past decade a great number of different plant genes have been cloned and well characterized. According to the accumulated data, we can conclude that the plant genes are very much like animal or yeast genes. The plant genes use the same genetic code (are split by introns) and basically use the same approaches of gene regulation. However; plant genes have differences compared to animal genes. In plant genes the requirements for splice site recognition are subtly different. Differences can be observed in plant promoters; although in general, the basic aspects remain the same. However, they are extremely different when they are compared to bacterial genes. This aspect is very important to understand why the expression levels of bacterial genes expressed in plants is very low. Reports on engineering plants with different crystal protein genes, mostly under the control of the strong and constitutive CaMV 35S promoter, have been published. Compared to other genes transferred to plants, the cry genes are weakly expressed in transgenic plants. Plants transformed with the full length cry sequences encoding the protoxin have extremely low levels of delta-endotoxin. Significant increases on recombinant protein production are obtained when a truncated version, encoding only the toxic fragment of the protoxin, is transferred and expressed in the plant. Using a truncated version of the cry IA(b) gene, the recombinant toxin detected in the leaves of transgenic tobacco plants represented up to 0.07 % of the total leaf protein; while only 0.0001 % is reported when full length genes were expressed (26). Even when only truncated versions of cry genes are considered for the generation of transgenic plants, the expression level remains low. Despite this fact, plants showed a remarkable entomocidal activity against target insects (27, 34). Although low expressing plants were protected against certain pests, higher expression levels and more specific crystal proteins are required to control agronomically important insects. Plant Promoters The detailed studies clearly demonstrated that the promoter is not simple at all but a rather complex array of various regulatory cis-acting elements and the sum of all these specialized "promoter units" that results in a constitutive or specialized promoter. This model is also applicable to other eukaryotic regulatory systems and it is defined as the combinatorial model of promoter function (35). Small cis- acting elements have usually been localized in the upstream sequences that can confer correct regulation upon the foreign gene. Proteins that bind these specific cis- acting elements have been isolated and their coding genes cloned and characterized. Now, one of the most important goals in plant molecular biology is to understand how the different transcription factors cooperate with each other and interact with the cis-acting elements in promoters, bringing the fine way of regulation of individual genes. The high yields of production of foreign proteins in transgenic plants by efficient gene expression remains as one of the most important aspects in plant molecular biology. The 35S Cauliflower Mosaic Virus (CaMV) is widely used in plant genetic engineering and it is considered as one of the strongest promoters available up to now. However, the 35S CaMV promoter shows preferential activity in dicotyledonous plant cells (Figure 3) (36). The base composition and function of 35S CaMV promoter sequence has been established (35, 37). Better results are obtained in monocotyledonous plants by using other strong and constitutive promoters: the rice actin 1 promoter (38), the maize Emu promoter (39) and the maize polyubiquitin 1 promoter (40, 41). The combination of these elements, previously reported as important for transcriptional activity in their original gene systems, offers the possibility to obtain promoters that could drive gene expression more efficiently or in a specific manner. In the design of chimeric promoters for the high efficiency expression of foreign genes in dicotyledonous or monocotyledonous transgenic plants, elements from different gene systems are often combined. This includes small enhancer sequences such as the 18 bp enhance element of the A. tumefaciens octopine synthase promoter (18 bp OCS). Usually, such elements are chemically synthesized and inserted in the promoter sequence in single or multiple forms at different distances from the gene transcription start.
Expression of Bacterial Genes in Transgenic Plants. Modification of Cry Genes Modifications of the cry coding sequence resulted in increased expression levels in tobacco, tomato cotton, potato (30-32) and maize (33). To make the bacterial cry genes more "plant-like", it is necessary to analyze some important aspects of the gene expression process in order to determine the modifications in the typical cry gene sequence that could enhance the gene expression in plant cells. The modifications we suggest are: A- Introduction of the preferential codon usage presented by plant genes. B- Elimination of CG and TA dinucleotides at codon positions 2 and 3. C- Conservation of the A+T base composition according to the proportion presented by plant genes. D- Modification of sequences that might cause mRNA destabilization, including premature polyadenilation signals, mRNA degradation signals, RNA-polymerase II termination signals and signals for splicing E- Reduction of secondary structure hairpins of the mRNA. F- Optimization of the ATG consensus flanking nucleotides for protein translation initiation and termination. G- Introduction of viral untranslatable mRNA leader to improve the initial steps of translation. The codon usage patterns in bacteria and plants exhibit remarkable differences (Table 1) (42). The modified gene could contain less AT-rich regions but it is recommended that its codon usage pattern must be intermediate between the more AT- rich bacterial cry codons and the more GC-rich monocotyledonous codon preference (29). The dinucleotide frequencies of GC used at codon position 2 and 3 must be conserved when no contradictions with plant preferential codon usage are observed. The CG dinucleotides are strongly avoided in plant genes, probably due to regulation involving methylation (42). Codons for Ala, Pro, Ser and Thr containing CG at positions 2 and 3 are rarely used in plants. In dicotyledonous plants the CG-ending codons are always the least favored but in monocotyledonous plants this is not always the case. In most eukaryotes, including plants, the TA- ending codons are also less favored. Concerning the A+T composition, the cry genes show about 60 % to 70 % A+T, representing a proportion 10 % to 15 % higher than that found in typical plant gene coding regions. In plants the A+T is particularly abundant in the intergenic and regulatory regions. Table 1. Codon usage in pooled sequences of Bacillus thuringiensis cry genes and nuclear genes of higher plants. The analysis included 30 410, 46 547, 39 155 and 39 155 codons from B. thuringiensis, dicotyledonous, and monocotyledonous plant genes respectively.
Amino Codons Bacillus Plants Dicotyl Monocotyl
Acid thuringiensis edonous edonous
n= 40 n= 280 Plants Plants
No. % No. % n= 155 n= 125
No. % No. %
--------------------------------------------------------------
Gly GGG 356 19.4 1157 17.1 391 11.0 764 24.0
GGA 784 42.8 1980 29.3 1448 40.6 533 16.7
GGT 480 26.2 1654 24.5 1183 33.1 478 15.0
GGC 213 11.6 1963 29.1 1545 15.3 1418 47.3
Glu GAG 1475 74.1 2777 58.5 1471 48.2 1304 77.1
GAA 517 25.9 1971 41.5 1583 51.8 388 22.9
Asp GAT 1338 81.6 1740 45.9 1383 60.4 356 23.8
GAC 301 18.4 2048 54.1 908 39.6 1139 76.2
Val GTG 328 16.6 1911 33.3 913 28.2 1006 40.1
GTA 824 41.6 540 9.4 378 11.7 165 6.6
GTT 605 30.9 1851 32.2 1406 43.4 439 17.5
GTC 216 10.9 1440 27.1 540 16.7 897 35.8
Ala GCG 255 15.3 1170 16.5 913 28.2 940 26.5
GCA 623 37.4 1457 21.0 378 11.7 513 14.5
GCT 563 33.8 2383 33.5 1406 43.4 736 20.7
GCC 225 13.5 2083 29.0 540 16.7 1359 38.3
Arg AGG 146 9.3 943 25.7 503 24.5 439 27.9
AGA 666 42.4 380 21.3 638 30.5 137 8.7
CGG 61 3.9 343 9.3 70 3.4 270 17.2
CGA 262 16.7 266 7.3 192 9.2 71 4.6
CGT 353 22.5 592 16.0 461 22.0 133 8.5
CGC 81 5.2 746 20.4 228 10.4 521 33.2
Ser AGT 366 25.2 711 12.3 564 16.9 153 6.3
AGC 185 8.2 1208 20.9 592 17.8 618 25.3
TCG 149 6.6 557 9.6 168 5.0 388 15.9
TCA 484 21.6 1011 17.5 671 20.1 337 13.8
TCT 623 27.8 1195 20.7 829 24.9 360 14.7
TCC 237 10.6 1097 19.0 508 15.3 587 24.0
Lys AAG 243 18.3 2957 67.8 1665 59.2 1292 83.1
AAA 1085 81.7 1405 32.2 1145 40.8 262 16.9
Asn AAT 1657 74.9 1294 37.9 1025 47.8 270 21.3
AAC 557 25.1 2117 62.1 1118 52.2 998 78.7
Met ATG 456 100.0 1988 100.0 1125 100.0 865 100.0
Ile ATA 600 30.1 643 15.4 447 17.8 199 12.0
ATT 1040 52.3 1577 37.9 1178 46.7 403 24.4
ATC 350 17.6 1945 46.8 894 35.5 1053 63.6
Thr ACG 381 17.9 506 12.1 154 5.2 352 20.9
ACA 894 40.8 960 22.9 1171 39.8 227 13.5
ACT 603 28.5 1303 31.1 983 33.4 321 19.0
ACC 271 12.8 1423 33.9 638 21.6 787 46.0
Trp TGG 429 100.0 1080 100.0 601 100.0 485 100.0
Cys TGT 213 70.5 574 34.7 382 45.8 188 23.1
TGC 89 29.5 1080 65.3 452 54.2 626 76.4
Tyr TAT 1171 79.5 926 33.9 652 43.2 274 22.4
TAC 302 20.5 1808 66.1 857 56.8 951 77.6
Leu TTG 289 10.9 1534 21.4 1025 26.7 509 15.2
TTA 1244 47.1 523 7.3 410 10.7 110 3.3
CTG 153 5.8 1208 16.8 294 7.7 916 27.4
CTA 332 12.6 634 8.9 303 7.9 337 10.1
CTT 511 19.4 1542 21.5 1108 28.9 439 13.1
CTC 110 4.2 1731 24.1 694 18.1 1034 30.0
Phe TTT 1047 78.9 1362 39.3 955 46.9 411 28.4
TTC 280 21.1 2108 60.7 1080 53.1 1034 71.6
Gln CAG 232 16.9 2468 40.4 699 39.8 1774 40.8
CAA 1143 83.1 3634 59.6 1057 60.2 2576 50.2
His CAT 499 85.7 745 44.4 475 55.2 266 33.2
CAC 83 14.3 934 55.6 386 44.8 536 66.8
Pro CCG 204 16.7 934 16.8 204 7.8 732 24.6
CCA 557 45.5 2374 42.6 1136 43.6 1237 41.6
CCT 405 33.1 1311 23.6 885 34.1 427 14.3
CCC 57 4.7 951 17.0 377 14.5 580 19.5
STOP TGA 6 15.0 100 35.7 42 27.1 68 54.4
TAG 6 15.0 78 27.9 28 18.1 32 25.6
TAA 28 70.0 102 36.4 85 54.8 25 20.0
n= The number of DNA sequences in the sample.
No.= The number of ocurrences of a given codon in the sample.
%= Percent ocurrence for each codon within a given aminoacid
in the sample.In eukaryotic cells, as a result of transcription, primary transcripts from nuclear genes called heterologous nuclear mRNA (hnRNA) are generated and then extensively processed. The hnRNA processing includes 5' capping, intron splicing and polyadenilation to form the mature polyA-mRNA. This is the only translatable form of mRNA. Control of gene expression at the level of mRNA stability is very important in plants (43, 44). Plants are sessile organisms, unable to move from adverse situations and forced to respond to environmental stimuli by altering endogenous gene expression. An important aspect of this regulation is the control of transcript processing. Most of RNAs in higher eukaryotes are relatively stable with a half-life period in the order of several hours (43). For many transcripts, differences in mRNA stability contribute to the establishment of steady-state mRNA levels and also to the speed at which those levels are achieved after a modification in transcription rate (45). Proteins that are required only transiently by the cell are encoded by unstable transcripts with half-lives in the order of an hour or less. Unstable transcripts are expected to act mostly in the rapid modification in gene expression that are present at mRNA level. This is necessary because, even after the immediate stop of transcription, such transcripts could remain functional for translation during several hours (46). In plants, the genes with a functional role in growth, development and rapid environmental responses are known to be encoded by unstable transcripts (46-48). One example is the phytocrome (Phy A) message in oats (49). Treatment of etiolated oat seedlings with red light causes a large decrease in the transcription rate of the Phy A gene and a rapid decrease in the level of steady-state Phy A mRNA. The cry genes contain DNA sequences that might potentially contribute to RNA instability in plants. Some of them are plant polyadenilation signals such as AATAAA and its variants, others could be a potential termination signal for the RNA-polymerase II. The consensus sequences on this type of signal are not completely defined; however, according to results obtained in electroporated Arabidopsis thaliana protoplasts, the sequences CAN[7-9]AGTNNA next to the 3' end of the U2 snRNA coding region is likely to play an important function in transcription termination (50). There are also mRNA degradation signals such as poly ATTTA (51) and sequences signaling wrong mRNA splicing. Recent results show several different mechanisms of mRNA degradation in eukaryotic cells. One mRNA decay pathway starts by the shortening of the poly(A) tail followed by decapping and 5' to 3' exonucleolytic degradation of the transcript. Some specific transcripts can be degradated through the deadenilation- independent decapping followed by 5' to 3' degradation. This process can also begin by endonucleolytic cleavage in the transcript. All these mechanisms are more or less related and suggest a model of RNA turnover in which polydenilated RNAs are degradated by the 'default' pathway initiated by poly(A) shortening (51). The eukaryotic mRNA can be degradated by endonucleolytic cleavage prior to deadenilation (52, 53). The degradation signal ATTTA (AUUUA) can destabilize transcripts in plants as well as in animals. This fact suggests that the pathway of such degradation may be highly conserved among many eukaryotes (53-55). For transcription termination, it is determined that TAA is preferentially used in dicotyledonous plants where monocotyledonous plants prefer the UGA termination codon (Table 1). The most important aspect in the stop codon context is the preference for A in the position +1 the based placed, following the stop codon and the avoidance of C in the same position. It has been demonstrated in E. coli that release factors require a tetranucleotide sequence for recognition and the same was observed in rabbit reticulocyte cell-free. Recent data suggest that the plant stop codon context may be UAAA, UGAA and UAGA (56). The BT-toxin transcripts are generally AU-rich and contain multiple copies of both AUUUA and AUUAA motifs that may contribute to mRNA instability (57). The cry IA(b) gene, for example, contains in the coding region 18 potential plant polyadenilation signals and 13 mRNA degradation signals (33). The expression of cry genes in transgenic plants requires mRNA levels buffering against modifications in transcription rates and must be stable. For this reasons the sequences must be modified and the signaling consensus eliminated (1, 29) and the consensus sequence of plant translational initiation and plant stop codon context introduced. The secondary structure hairpins of the mRNA could be reduced in order to enhance the translocation speed of ribosomes through the mRNA (58-61). It has been demonstrated that the negative effect of minor arginine codons on gene expression is suppressed by the introduction of the preferential codons within the first 25 genes in E. coli (62, 63). The cry IA(b) gene was modified at its 5' end to prevent such structures in the mRNA during initiation of translation which is considered to be one of the critical steps in the gene expression process. The modification consists of the substitution of the first 55 codons by their most abundant homologues in plant genes (Figure 4). The modifications introduced in the coding region cover only 8 % of the full- length transcript. The secondary structure hairpins were predicted for mRNA of wild-type and modified versions of the truncated cry IA(b) taking into account the use of the CaMV 35S promoter, the 70 bp TMV omega-fragment (Figure 4). Increases of up to five folds were obtained when the modified cry IA(b) gene was expressed, driven by the CaMV 35S promoter in electroporated tobacco protoplasts (de la Riva, unpublished). Surprisingly, the expression of a modified gene in E. coli resulted in over-production of the recombinant toxin although the modifications introduced were designed to enhance the expression in plant cells. These facts suggest a prevailing role of the 5' end secondary structure of mRNA in the modulation of gene expression particularly during the initial steps of translation.
Translation Efficiency Almost all plant mRNAs are polyadenilated. It has been established that poly(A) increases message stability and it also functions as a regulator of translation efficiency (64). The regulation of mRNA stability is independent from a similar function of the cap at the 5' end. Despite this fact, the poly(A) tail functions as a translation regulator only when the transcript is capped. During initiation, the activity of the cap depends on the poly(A) tail. This synergism suggests that this functional conjunction with the associated protein complex is in communication during translation. This contact could direct protein-protein contacts between eIF-4F and poly(A) binding protein (PAB), or it could be mediated by other factors (65, 66). Translation in plants is more dependent on the factors mentioned above than it is in animals. Little is known about the mechanism involved in the protection from nucleases of mRNA mediated by the poly(A) tail. Coincidentially, the removal of the poly(A) tail may constitute an early step in the mRNA degradation process (51). In order to form the consensus of the plant translation start sequence, the modification of the ATG flanking sequence consists of the introduction of minimal modifications in the original cry sequence but it enhances significantly the yields of the recombinant protein (65). Together with the modification of the coding region, it is common to find the use of viral untranslatable leaders, like the 70 bp TMV omega- fragment and the 600 bp leader preceding the CaMV open reading frame VII, in order to increase the affinity of mRNA to the small 40S ribosomal subunit (66, 67). During the first step of viral infection, this type of sequences helps viral transcripts to be more "competitive" in binding ribosomes than the host mRNA, increasing the translation efficiency. The same approach can be designed to favor the termination step of mRNA translation. These elements interact with translation initiation factors present in dicotyledonous plants such as plant-eIF-4E, the cap-binding protein and plant-eIF-4A (a putative RNA helicase). This RNA helicase is thought to unwind secondary RNA structure in the 5' leader of mRNA to enable the scanning ribosome to reach the initiator AUG. The plant-eIF-4A is active in other organisms like Xenopus oocytes where they can activate dormant mRNAs. The translation initiation factors from wheat are active in rabbit reticulocyte cell-free. They are encoded by a multigene family of highly divergent genes. The translation initiation factors in dicotyledonous plants enhance the translation of the mRNA carrying the TMV omega-fragment and interacting with the leader sequence, while in monocotyledonous plants, the recognition of a leader sequence by the translation initiation factor is less effective and in consequence, the enhancement of translation is less favored. On this basis it is possible to understand why the TMV omega-fragment shows a better effect in dicotyledonous plants (68). None of the mutations in these sequences influence steady-state mRNA level. This fact allows to conclude that the effects must be at a translation level. A translational enhancer derived from tobacco mosaic virus is functionally equivalent to a Shine-Delgarno sequence (68, 69). Such elements have been described for the 5'-leaders of several plant viruses. The elucidation of the function of the viral tRNA-like structure in the cytoplasmatic gene regulation could bring about new ideas for further strategies to improve the expression of foreign genes in transgenic plants (70). Ribosome translocation was thought to proceed at a constant speed but it has been observed in vitro that the ribosome can pause and stack (58). As it was discussed above, we can conclude that the stable hairpin secondary structure mRNA might result in a less efficient translation initiation. The same effect occurs during the elongation of a polypeptide chain and it might also cause pausing and stacking of the ribosomes. Little is known about how elongation is really regulated. It has been established that in vitro ribosome translocation is sensitive to pH, which is an important fact for the control of translation during conditions of hypoxia in which the cytosol acidifies (64). Concluding Remarks The first results on insect control by transgenic plants expressing the cry gene were published in 1987 (24, 26). Other experiments have been performed since that moment. The insecticidal activity on tobacco plants were tested in tobacco hornworm (Manduca sexta) and tobacco budworm (Heliothis virescens), the major pests of this crop. Experiments were also carried out with transgenic potato plants expressing a coleopteran-active cry IIIA gene from B. thuringiensis var. tenebrionis. Transgenic plants resistant or tolerant to insect attacks have been produced in tomato, tobacco, potato, cotton, maize and more recently in sugarcane. The potentials and the limitations of engineering insect resistance in plants by the expression of cry genes have been intensively explored. Plants frequently attacked by insects highly susceptible to B. thuringiensis delta- endotoxin can be protected using genetic engineering. The effectiveness of the control depends on the yields of recombinant toxin in the plant. High gene expression level can be obtained only if a modified "eukaryotic-like" cry gene is transferred to plants under the control of an efficient promoter. The possibilities for the development of resistance to B. thuringiensis delta-endotoxin in genetically modified plants is a polemic topic subject to active and permanent discussion. The fact that different B. thuringiensis delta-endotoxins can bind different proteins could help prevent the development of resistance. In laboratory conditions, the development of resistance to one delta-endotoxin can be studied. Therefore; the susceptibility of the resistant insect to others can be known. These tests lead us to suggest the combination of different toxin genes in plants or the alternation of transgenic plants expressing different toxins. Another strategy consists of the limitation of gene expression only to the economically important organs of the plant but this requires an extensive work for the construction and use of efficient and specific tissue promoters. The determination of the molecular aspects of the mode of action of delta-endotoxins, including the structure and function of a larval midgut receptor, will be essential in order to design strategies to control the development of resistance in insects. During the laboratory evaluation, the most promising transgenic plant lines must be selected for further experiments in field conditions. At the same time, a great number of independent clones is needed in order to obtain some high expressing plant lines. To test all these plants, whenever possible, it is recommended to carry out a challenge experiment, feeding neonate larvae of the target insect on transgenic clones. The selected clones must be characterized by molecular biology (CPR, Southern blot, Northern blot) and immunochemical methods (ELISA, IRMA, Western blot) and the transgenic plant lines showing effective protection against insect attack must be evaluated at field conditions. The screening projects could lead to the discovery of B. thuringiensis strains with novel entomocidal specificities which may be the source of novel delta-endotoxin genes potentially suitable for expression in transgenic plants. Extensive screening programs are being carried out by various groups in order to search for B. thuringiensis with such new insecticidal spectra. The production of transgenic plant lines resistant or tolerant to insect attack is not the answer for all insect problems in agriculture but it is an alternative method to be integrated in the current pest management strategies. Other methods for the manipulation and use of B. thuringiensis cry genes, including the development of genetically engineered strains with improved delta-endotoxin production and novel entomocidal specificities are very promising. Acknowledgements We wish to thank Viviana Falcon (CIGB, Havana, Cuba) for B. thuringiensis microphotography and other related works. References 1.Adang MJ. Bacillus thuringiensis insecticidal crystal proteins: gene structure, action and utilization. In: Maramorosch K (ed.) Biotechnology for biological control of pest and vectors. CRC Press, Boca Raton, Fl., USA 1991;3-24 2.Luo K, Adang MJ. Removal of adsorbed toxin fragments that modify Bacillus thuringiensis CryIC delta-endotoxin iodination and binding by sodium dodecyl sulfate treatment and renaturation. Appl and Environ Microbiol 1995;60:2905-2910. 3.Jun Chen X, Curtiss A, Alcantara E, Dean D. Mutations in domain of Bacillus thuringiensis delta-endotoxin cryIA(b) reduce the irreversible binding of toxin to Manduca sexta brush border membrane vesicles. J of Biol Chem 1995;270:6412-6419. 4.Feiltelson JS, Payne J, Kim L. Bacillus thuringiensis: Insects and beyond. Biotechnology 1992;10:271-275. 5.Herrera-Estrella L, G Den Broeck Van, Maenhaut R, Van Montagu M, Schell J, Timko M, Cashmore A. Ligh-inducible and chloroplast-associated expression of a chimeric gene introduced into Nicotiana tabacum using a Ti plasmid vector. Nature 1984;310:115-120. 6.Horsch RB, Fry JE, Hoffman NL, Wallroth M, Eichholtz D, Rogers SG, Fraley RT. A simple and general method for transferring genes into plants. Science 1985; 227: 1229- 1231. 7.Shillito R, Saul M, Paszkowski J, Muller M, Potrikus I. High efficiency direct transfer to plants. Biotechnology 1985;3:1099-1103. 8.Uchimiya H, Fushimi T, Hashimoto H, Harada H, Syono K, Sugawara Y. Expression of a foreign gene in callus derived from DNA-treated protoplasts of rice (Oryza sativa L.). Mol Gen Genet 1986;204:204-207. 9.de la Pena A, Lorz H, Schell J. Transgenic rye plants obtained by injecting DNA into young floral tillers. Nature 1987; 325: 274-276. 10.Arencibia A, Molina P, de la Riva GA, Selman-Housein G. Production of transgenic sugarcane (Saccharum officinarum L. plants)by intact cell electroporation. Plant Cell Report 1995;14:305-309. 11.Fromm M, Taylor L, Walbot V. Expression of genes transferred into monocotyledonous and dicotyledonous plant cells by electroporation. Proc Natl Acad Sci USA 1985;82:5824- 5828. 12.Fromm M, Taylor L, Walbot V. Stable transformation of maize after gene transfer by electroporation. Nature 1986; 319: 791- 793. 13.Lorz H, Baker B, Scell J. Gene transfer to cereal cells mediated by protoplast transformation. Mol Gen Genet 1985; 199: 473-497. 14.Potrikus I. Gene transfer to cereals: an assessment. Trends Biotech 1989;8:33-38. 15.Sanford J. The biolistic process. Trends Biotechnol 1988;6:299-302. 16.Graves A, Goldman S. The transformation of Zea mays seedling with Agrobacterium tumefaciens: detection of T-DNA specified enzymes activities. Plant Mol Biol 1986; 7:43- 50. 17.Hooykaas G, Hooykaas P, Schilperoot R. Expression of Ti plasmid genes into monocotyledonous plants infected with A. tumefaciens. Nature 1984;311:763-764. 18.Rainieri D, Bottino P, Gordon M, Nester E. Agrobacterium- mediated transformation of rice (Oriva sativa L.). Biotechnology 1990; 8:33-38. 19.de la Riva GA, Oramas P, Goyenechea B, Perez S. Cloning and expression of a Bacillus thuringiensis var. berliner delta-endotoxin gene by PCR. Interferon y Biotecnologia 1989;6:234-241. 20.de la Riva GA, Xoconostle-Cazares B, Gutierrez C, Moran R, lvarez A, Herrera-Estrella L, Perez S. Heterologous expression of Bacillus thuringiensis var. tenebrionis delta-endotoxin gene in E. coli. Biotecnologia Aplicada 1992;9:31-37. 21.Held GA, Bulla LA, Ferrari E, Hoch J, Aronson AI, Minnich A. Cloning and localization of the Lepidopteran protoxin gene of B. thuringiensis subsp. kurstaki. Proc Natl Acad Sci USA 1982;79:6065-6069. 22.Klier A, Fargette F, Ribier J, Rapoport G. Cloning and expression of the crystal protein genes of B. thuringiensis strain berliner 1715. EMBO J 1982;1:791-799. 23.Schnepf E, Whiteley HR. Cloning and expression of B. thuringiensis crystal protein gene in E. coli. Proc Natl Acad Sci USA 1981;84:7036-7040. 24.Vaithilingan S, Thompson DV, Mororey MJ, Bookland RG, Adang MJ. Molecular cloning and characterization of the insecticidal crystal protein gene of B. thuringiensis var. tenebrionis. Proc Natl Acad Sci USA 1987;84: 7036- 7040. 25.Shivakumar AG, Gundling GL, Benson TA, Casuto D, Miller MF, Spear BB. Vegetative expression of the delta-endotoxin genes of B. thuringiensis subsp. kurstaki in B. subtilis. J of Bacteriol 1986;166:194-204. 26.Vaeck M, Reynaerts A, Hofte H, Jansens S, De Beukeleer M, Dean C, Zabeau M, Van Montagu M, Leemans J. Transgenic plants protected from insect attack. Nature 1987; 28:33-37. 27.Delannay X, La Vallee BJ, Proksch RK, Fuchs RL, Sims SR, Greenplate JT, Marrone PG, et al. Field performance of transgenic tomato plants expressing the B. thuringiensis var. kurstaki insect control protein. Biotechnology 1989;7:1265-1269. 28.Perlack FJ, Deaton RW, Armstrong TA, Fuchs RL, Sims SR, Greenplate JT, Fischhoff DA. Insect resistant cotton plants. Biotechnology 1990;8:939-943. 29.Perlak FJ, Fuchs RL, Dean DA, McPherson SL, Fischhoff DA. Modification of the coding sequence enhances plant expression of insect control protein genes. Proc Natl Acad Sci USA 1991;88:3324-3328. 30.Adang MJ, Rocheleau TA, Merlo DJ, Murray EE. Synthetic insecticidal crystal protein gene. EPO patent application 0 359 472 A2. 1990. 31.Adang MJ, Brody MS, Cardineau G, Eagan N, Roush RT, Shewmaker CK, Jones A, Oakes JV, McBride K. The reconstruction and expression of a Bacillus thuringiensis cryIIIA gene in protoplasts and potato plants. Plan Mol Biol 1993;21:1131- 1145. 32.Perlak FJ, Stone TB, Muskopf YM, Petersen LJ, Parker GB, McPherson SL, Wyman J, Love S, Reed G, Biever G, Fischhoff DA. Genetically improved potatoes: protection from damage by colorado potato beetles. Plant Mol Biol 1993;22:313-321. 33.Koziel GM, Beland GL, Bowman C, Carozzi NB, Crenshaw R, Crossland L, et al. Field performance of elite transgenic maize plants expressing an insecticidal protein derived from Bacillus thuringiensis. Biotechnology 1993;11:194-200. 34.Arencibia A, Vazquez R, Coego A, Prieto D, de la Riva GA, Selman-Housein G. Transgenic sugarcane (Saccharum officinarum L.) plants are tolerant to stem borer (Diatraea saccharalis F.) attack despite the low expression levels of cryIA(b) gene from B. thuringiensis var. kurstaki HD-1. Advances in Modern Biotechnology 1995;3: 71. 35.Benfey PN, Chua NH. The cauliflower mosaic virus 35S promoter: combinational regulation of transcription in plants. Science 1990;250:959-966. 36.Rong-Xiang F, Nagy F, Sivasubramanian S. Multiple cis regulatory elements for maximal expression of the CaMV promoter in transgenic plants. The Plant Cell 1989; 1:141- 150. 37.Guilley H, Dubley RK, Jonard G, Balazs E, Richards KE. Transcription of cauliflower mosaic virus DNA: detection of promoter sequences and characterization of transcripts. Cell 1982;30:763-773. 38.McElroy D, Zhang W, Cao J, Wu R. Isolation of an efficient actin promoter for use in rice transformation. The Plant Cell 1990;2:163-171. 39.Last DI, Brettell RIS, Chamberlaine DA, Chaudhury AM, Larkin PJ, Marsh EL, et al. pEmu: An improved promoter for gene expression in cereal cells. Theor Appl Gen 1991;81:581-588. 40.Schledzewski K, Mendel R. Quantitative transient gene expression: comparison of the promoters for maize polyubiquitin 1, rice actin 1, maize derived Emu and CaMV 35S in cells of barley, maize and tobacco. Transg Res 1994;3:249- 255. 41.Christensen AH, Sharrock SA, Quail PH. Maize polyubiquitin genes: structure, thermal perturbation of expression and transcript splicing, and promoter activity following transfer to protoplasts by electroporation. Plant Mol Biol 1992; 18:675-689. 42.Murray EE, Lotzer J, Eberle M. Codon usage in plant genes. Nucl Acid Res 1989; 17:477-498. 43.Green PJ. Control of mRNA stability in higher plants. Plant Physiol 1993; 102: 1065-1070. 44.Gallie DR. Post-transcriptional regulation of gene expression in plants. Ann Rev Plant hysiol and Plant Molec Biology 1993; 44:77-105. 45.Herrick D, Parker R, Jacobson A. Identification and comparison of stable and unstable mRNA in Saccharomyses cerevisiae. Mol Cell Biol 1990;10:2269-2284. 46.Taylor CB, Green PJ. Identification and characterization of genes with unstable transcripts (GUTs) in tobacco. Plant Mol Biol 1995;28:27-38. 47.Mahadevan LC, Edwards DR. Signalling and superinduction. Nature 1991; 349: 747-748. 48.Sullivan MJ, Green PJ. Post-transcriptional regulation of nuclear-encoded genes in higher plants: the role of mRNA stability and translation. Plant Mol Biol 1993; 23: 1091- 1104. 49.Seeley KA, Byrne DH, Colbert JT. Red light-independent instability of oat phytochrome mRNA in vivo. Plant Cell 1992; 4:29-38. 50.Shaw G, Kamen R. A conserved AU sequence from the 3' untranslated region of GM-CSF mRNA mediates selective mRNA degradation. Proc Natl Acad Sci USA 1986; 84:7036-7040. 51.Beelman CA, Parker R. Degradation of mRNA in eukaryotes. Cell 1995;81:179-183. 52.Brock ML, Shapiro DJ. Estrogen stabilizes vitellogenin mRNA against cytoplasmic degradation. Cell 1983;34:207-214. 53.Stockle MY, Hanafusa H. Processing of 9E3 mRNA and regulation of its stability in normal and Rous Sarcoma Virus- transformed cells. Mol Cell Biol 1989; 9:4738-4745. 54.Kozak M. Regulation of translation in eukaryotic systems. 1992. Annu ev Cell Biol 1989;229-241. 55.Hunt AG. Messeger RNA 3' end formation in plants. Ann Rev Plant Physiol Plant Mol Biology 1994;45:47-60. 56.Angenon G, Van Montagu M, Depicker A. Analysis of the stop codon context in plant nuclear genes. FEBS Lett 1990; 271: 144-146. 57.Ohme-Takagi M, Taylor CB, Newman TC, Green PJ. The effect of sequences with high AU content on mRNA stability in tobacco. Proc Natl Acad Sci USA 1993;90:11811-11815. 58.Wolin SL, Walter P. Ribosome pausing and stacking during translation of eukaryotic mRNA. The EMBO Journal 1988; 7: 3559-3569. 59.Gallie DR. The cap and poly(A) tail function sinergically to regulate mRNA translational efficiency. Genes and Devel 1991;5:2108-2116. 60.Gallie DR, Feder JN, Shimke RT, Walbot V. Post- transcriptional regulation in higher eukaryotes: The role of the reporter gene in controlling expression. Mol Gen Genet 1991;228:258-264. 61.Kozak M. Influences of mRNA secondary structureon initiation by eukaryotic ribosomes. Proc Natl Acad Sci USA 1986; 83: 3850-3854. 62.Gia-Fen C, Inouye M. Suppression of the negative effect of minor arginine codons on gene expression: preferential use of codons within the first 25 codons of the E. coli genes. Nucl Aci Res 1990; 18: 1465-1473. 63.Inouye M, Gia-fen C. Regulation of gene expression by minor codons in E. coli; minor codon modulation hypothesis. NATO ASI Series 1990;49:217-225. 64.Webster C, Kim C, Roberts JM. Elongation and termination reaction of protein synthesis in maize root-tips polyribosomes studied in a homologues cell. Plant Physiol 1991;96:418- 425. 65.Kozak M. Point mutations define a sequence flanking AUG initiator codon that modulates translation by eukaryotic ribosomes. Cell 1986;44:283-292. 66.Taylor JL, Jones JDG, Sandler S, Mueller GM, Bedbrook J, Dunsmuir P. Optimizing the expression of chimeric genes in plant cells. Mol Gen Genet 1987;210:572-577. 67.Jobling SA, Gehrke L. Enhanced translation of chimeric messenger RNAs containing a plant vial untranslated leader sequence. Nature 1987;325:622-625. 68.Gallie DR, Kado CI. A translational enhancer derived from tobacco mosaic virus is functionally equivalent to a Shine- Delgarno sequence. Proc Natl Aca Sci USA 1989; 86: 129-132. 69.Gallie DR. The 5'-leader sequence of tobacco mosaic virus RNA enhances the expression of foreign gene transcripts in vitro and in vivo. Nucl Aci Res 1987; 15: 8693- 8711. 70.Gallie DR, Feder JN, Schimke RT, Walbot V. Functional analysis of the tobacco mosaic virus tRNA like structure in cytoplasmatic gene regulation. Nucl Aci Res 1991; 19:5031- 5036. Copyright 1996 Elfos Scientiae
The following images related to this document are available:Photo images[ba96101a.jpg]Line drawing images[ba96101b.gif] [ba96101c.gif] [ba96101d.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}