 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Reporter Genes For Transgenic Fish Experiments Isabel Guillen,^1 Ricardo Lleonart,^1 Diana Garcia del Barco,^1 Rebeca Martinez,^1 Fidel Herrera,^1 Antonio Morales,^1 Maria Teresa Herrera,^1, 2 Reinold Morales^1 and Jose de la Fuente^1
^1 Division de Genetica Celular de Mamiferos. Centro de
Ingenieria Genetica y Biotecnologia. P.O. Box 6162, La Habana
6, Cuba.
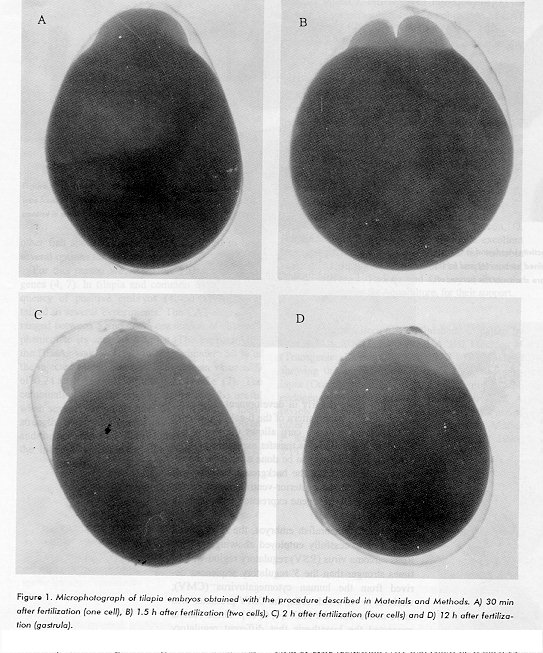
ABSTRACT The study of regulatory sequences in transient experiments employing reporter genes produce valuable data to design gene constructs for transgenesis. In this report, Escherichia coli chloramphenicol acetyl transferase (CAT), hepatitis B surface antigen (HBsAg), E. coli beta-galactosidase and the green fluorescent protein (GFP) from the jellyfish Aqueorea victoria were assayed as reporter genes for in vivo transient expression in tilapia, common carp and zebrafish. In tilapia and common carp, an endogenous beta-galactosidase-like activity was found in embryos, and after hatching, it was localized in the posterior-ventral part of the fry. This beta-galactosidase-like activity could interfere with the use of lacZ as a reporter gene in experiments with tilapia and common carp when the expression is assayed in the early stages of development. In zebrafish, however, lacZ was successfully employed. The HBsAg and CAT genes gave reproducible results in the species tested, thus constituting a choice when a second reporter gene is needed as an internal control. Finally, the GFP provided a simple and powerful mean of monitoring transient gene expression in live zebrafish embryos. Key words: carp, tilapia, zebrafish, galactosidase, embryo RESUMEN El estudio de secuencias reguladoras empleando genes reporteros en experimentos de expresion transitoria permite disenar y caracterizar construcciones geneticas para transgenesis. En este reporte, los genes para cloranfenicol acetil transferasa (CAT) y lacZ de Escherichia coli, el antigeno de superficie de la hepatitis B (HBsAg) y la proteina fluorescente verde (GFP) del pez Aequorea victoria se ensayaron como genes reporteros para ensayos de expresion transitoria en tilapia, carpa comun y pez zebra. En embriones de tilapia y carpa comun se encontro una actividad tipo galactosidasa beta que se localizo despues de la eclosion en la parte postero-ventral de la larva. Esta actividad tipo galactosidasa beta puede interferir con el empleo del gen lacZ como reportero en experimentos con tilapia y carpa comun cuando la expresion se ensaye en los estadios tempranos del desarrollo. En pez zebra, no obstante, el gen lacZ se empleo exitosamente. Los genes codificantes para HBsAg y CAT dieron buenos resultados en las especies ensayadas, constituyendo una opcion cuando se necesite un segundo gen reportero como control interno. Finalmente, la GFP resulto un medio simple y ventajoso para monitorear la expresion genica transitoria en embriones vivos de pez zebra. Palabras claves: carpa, tilapia, pez zebra, galactosidasa, embrion Introduction Transgenic fish have been generated by DNA microinjection into the germinal disc and, in some cases, the transgene has been expressed and inherited by the progeny of these animals (1-3). However, much remains to be elucidated before gene transfer can be fully manipulated to generate new strains of economically important fish species. In vivo transient experiments employing reporter genes produce valuable data to design gene constructs for transgenesis (4). The selection of the reporter gene for in vivo transient expression experiments requires that the reporter activity be easily detected given good sensitivity and specificity. Furthermore, because in many experiments it is necessary to monitor for two different reporters to employ one as a test signal and the second to normalize for intrinsic variations of the system, several options for reporter genes must be available for fish species suitable for laboratory work. To select appropriate reporter genes for in vivo transient expression experiments in tilapia, common carp and zebrafish fish, species which provide good laboratory models, several genes were assayed. The E. coli lacZ gene is one of the most useful reporter genes because it gives a good sensitivity and quickness. The histochemical staining using X-gal allows the detection of this enzyme in cells and tissues (5). The CAT gene has been widely employed as a reporter in in vivo and in vitro experiments (6, 7). The HBsAg was tested because of the high specificity and sensitivity of the analytical ELISA systems currently used for antigen determination (8) and the GFP for the possibility of employing this system in live fish (9). In tilapia (Oreochromis aureus) and common carp (Cyprinus carpio L.), an endogenous beta-galactosidase (beta-gal)-like activity was found in fish fry and embryos. This activity appeared soon after fertilization and continued during the yolk sac resorption. In more advanced developmental stages, the endogenous beta-gal-like activity was confined to the stomach and intestine. This beta-gal-like activity interferes with the use of lacZ as a reporter gene in transgenic fish experiments when the expression is assayed in the early stages of development. However, in zebrafish (Brachydario rerio) the lacZ reporter gene was successfully employed. CAT, HBsAg and GFP reporters showed reproducible results in the fish species tested. Materials and Methods Fish embryos and fry Adult tilapia were obtained from the Empresa Nacional de Acuicultura (Cotorro, La Habana) and placed, separately according to sex, in 1 L tanks at 18-20 C. Tilapia were fed ad libitum. Mature tilapia were selected by visual examination and placed in a 500 L glass aquarium with a biological filter under controlled temperature and photoperiod (28-30 C and 14 h light x 10 h dark). After spawning and fertilization, the female was carefully removed from the aquarium and the eggs were placed in a glass cylinder (dia. 20 cm) with a 500 mL incubation solution (0.1 % NaCl, 0.004 % CaCl2 2H2O, 0.003 % KCl, 0.016 % MgSO4 7H2O, 0.0001 % methylene blue). Embryos were incubated with gentle shaking (60 rpm) and the solution was changed every 8 h until hatching when shaking was stopped for further incubation. For common carp, embryos and fry were obtained as described by Hernandez, et al. (10). Microinjection of tilapia and carp embryos was conducted as described by de la Fuente, et al. (11) and Hernandez, et al. (10), respectively. Zebrafish embryos were obtained and microinjected as reported by Stuart, et al. (12). Detection of beta-galactosidase Tilapia and common carp fry and embryos were fixed for 1 h in 0.1 M NaHPO4 containing 4 % paraformaldehyde and 0.2 % glutaraldehyde at 0 C. Then, the samples were washed three times at room temperature with PBS, and incubated for 1 h in PBS containing 0.2 % BSA (fraction V, Sigma) and 0.1 % Triton X-100. Then, they were incubated for 14-16 h at 37 C in a solution of 1 mg/mL X-gal in PBS containing 5 mM K3[Fe(CN)6], 5 mM K4[Fe(CN)6] and 1.3 mM MgCl2. Before staining, adult organs were washed with sterile water for beta-galactosidase activity. Then, they were processed as described for fry and embryos but including 0.02 % NP 40 in the fixing solution and incubating with X-gal for 18 h at 30 C. In zebrafish embryos, beta-galactosidase activity was detected as described by Garcia del Barco, et al. (4). Detection of CAT and HBsAg CAT activity was detected in zebrafish embryos as described by Garcia del Barco, et al. (4). For tilapia and carp embryos, the procedure is emphasized in Lleonart, et al. (7). Briefly, two plasmids were used in the experiments: pE300, which has the CAT gene under the control of the human cytomegalovirus (CMV) enhancer-promoter region and 3' polyadenylation sequences from the SV40 genome (11), and pUCHBV (13), which contains the entire hepatitis B virus (HBV) genome except for the core sequence inserted in pUC19. Plasmids were diluted to 50 ug/mL in injection buffer (10 mM TrisHCl pH 7.4, 0.25 mM EDTA, 0.1 % Phenol Red) and about 1 nL was injected into one cell embryos 30 to 60 min after fertilization. The injected embryos were incubated for two to four days and, before hatching, they were transferred to eppendorf tubes, resuspended in 100 L of 250 mM TrisHCl pH 8, and about 50 uL of glass beads (0.5-1 mm) were added. Tubes were vortexed for 1 min, freeze-thawed three times (-70 C, +37 C) remaining 5 min at each temperature, and after the addition of 1 L of 0.5 M EDTA, the tubes were heated 10 min at 60 C. CAT assay was done by standard procedures and the HBsAg was determined by ELISA in embryo homogenates. Detection of GFP One cell zebrafish embryos were microinjected with the plasmid pRSGFP-C1, which contains the jellyfish GFP gene under the control of CMV promoter and SV40 polyadenylation signal (14). Thirty hours after the injection, embryos were placed under UV light in an Olympus IMT2 microscope (Japan) and the GFP visualized as described by Kevin, et al. (15). Results and Discussion The use of reporter genes to study regulatory sequences in vivo, allows the design of appropriate gene constructs for gene transfer experiments in fish (4). The possibility of employing the E. coli lacZ and CAT genes, the HBsAg and the jellyfish GFP as reporters in zebrafish, tilapia and common carp, was studied. With the procedure described in this paper, tilapia embryos were reproducibly obtained with a high rate of normal development (Figure 1).
In tilapia, an endogenous beta-gal-like activity was found in the fertilized egg associated with the embryos (Figure 2 A). Soon after hatching, this activity was localized in the posterior-ventral part, in the region between the stomach and the yolk sac (Figure 2 B). In the adult animal, the activity was present only in the stomach and intestine (Figure 3).
Figure 3. A beta-galactosidase-like activity was localized in adult tilapia stomach and intestine. The organs were extracted from adult tilapia and stained (A, C and E) or not (B, D and F) for beta-galactosidase-like activity. A and B) gonads, C and D) stomach, E and F) intestine. The analysis for each organ was repeated at least three times giving similar results. In carp, the endogenous beta-gal activity was found in fertilized eggs. This activity continued later in development and was found in four days old fry showing different expression patterns (Figure 4). This variegated gene expression may represent a form of gene regulation (16), and could be related to the age of the cell, its health or its position within the cell cycle (17). After yolk sac resorption (seven days after fertilization), the fry were stained again for beta-gal-like activity. As in tilapia, the activity appeared localized in the posterior-ventral region of the fry. Further characterization in adult organs confirmed that this beta-gal-like activity remained localized in the stomach and intestine.
These results indicated that the E. coli lacZ gene, when employed as a reporter gene in tilapia and carp, could interfere with the endogenous beta-gal-like activity if assayed early in development. However, the adult expression pattern of the beta-gal-like activity in tilapia and common carp allows the use of the E. coli lacZ gene in mass transfer experiments if the assay for its activity is to be done after yolk sac resorption. In this case, the background activity will be restricted to the posterior-ventral region of the fry, allowing the transgene expression to be assayed in other tissues. However, in zebrafish embryos, the E. coli lacZ gene was successfully employed showing that the Rous sarcoma virus (RSV) regulatory region was 17 times stronger than the 5' regulatory sequences derived from the human cytomegalovirus (CMV); while in in vitro fish cells, the CMV promoter was more active than the RSV (4, 6, 18). These results suggested the hypothesis that different regulatory requirements exist in cells and embryos and that chimeric constructs designed for transgenic experiments should be assayed in transient expression experiments in fish embryos (4). This hypothesis was recently corroborated in transgenic tilapia lines generated with transgenes containing different regulatory regions (11), showing that RSV enhancer promoter sequences were more active than those derived from the CMV (Hernandez, et al., manuscript in preparation). Endogenous background activities have been reported for beta-galactosidase (19) and neomycin (20) in other fish species. This confirms the need to have several options for reporter genes. For this reason we tested the HBsAg and CAT genes (4, 7). In tilapia and common carp, high frequency of positive embryos (40-50 %) were obtained in several experiments. The CAT expression ranged between 5 and 50 % conversion of chloramphenicol to its acetylated forms. The expression of the HBsAg was observed in approximately 50 % of the injected tilapia embryos with a mean value +/-SD of 0.74 +/- 0.16 ng/mL of the homogenate (7). The combination of these reporter genes may be useful when several constructs have to be tested for theirstrength, employing one of the genes as a reporter and the second as an internal standard to normalize the intrinsic variations of the system (amount of plasmid injected, efficiency of injection) (4, 7). In fact, in zebrafish embryos, the combination of CAT and lacZ reporter genes has been employed in our laboratory to assay different chimeric genes for transgenic fish experiments (4). Recent results with the luciferase gene in medaka (Oryzias latipes) and with the GFP gene in zebrafish embryos indicate that it can be used as a potential source for monitoring gene expression in live transgenic fish (9, 21). We have also shown the GFP expression in zebrafish embryos 30 h after injection and experiments are under way to assay this reporter gene in tilapia and common carp. Acknowledgments We would like to thank Drs. Viviana Falcon, O. Hernandez and Fidel O. Castro for their excellent technical assistance; Mario P. Estrada for valuable suggestions during the course of the experiments; and C. Uliver, J. Padron and O. Leyva, from the National Enterprise for Aquaculture, for their support. Note added in proof When this manuscript was finished, a paper by Samsul Akam MD, Popplewell A and Maclean N (Transgenic Research 1996; 5:87-95) appeared showing the expression of a lacZ transgene in tilapia (Oreochomis niloticus) and the staining of the endogenous beta-gal activity described in our work. A review manuscript also appeared covering the use of other reporter transgenes in fish (Iyengar A, Muller F and Maclean N. Transgenic Research 1996; 5:147-166). References 1. MacLean N, Penman D. The application of gene manipulation to aquaculture. Aquaculture 1990;85:1-20. 2. de la Fuente J, Hernandez O, Guillen I, Castro FO, Aguilar A, Herrera L, et al. Transgenesis in fish. Applications in Biotechnology. Biotecnologia Aplicada 1991;8:123-139. 3. de la Fuente J, Hernandez O, Martinez R, Guillen I, Estrada MP, Lleonart R. Generation, characterization and risk assessment of transgenic tilapia with accelerated growth. Biotecnologia Aplicada 1996;13(3):221-230. 4. Garcia del Barco D, Martinez R, Hernandez O, Lleonart R, de la Fuente J. Differences in the transient expression directed by heterologous promoter and enhancer sequences in fish cells and embryos. Journal of Marine Biotechnology 1994;1:203- 205. 5. Hall CV, Jacob PE, Ringold GM, Lee F. Expression and regulation of E. coli lacZ gene fusions in mammalian cells. Mol Appl Genet 1983;2:101-109. 6. Hernandez O, Theron MC, Puissant C, Bearzotti M, Attal J, leBail PY, et al. Effect of intervening sequences in the transient expression of tilapia growth hormone in mammalian and fish cells. Biotecnologia Aplicada 1993;10:158- 161. 7. Lleonart R, Martinez R, Garcia del Barco D, Hernandez O, Castro FO, de la Fuente J. Reporter genes for in vivo transient gene expression studies in tilapia (Oreochromis aureus) and common carp (Cyprinus carpio) one celled embryos. Theriogenology 1994;41:240. 8. Fernandez de Cossio ME, Diaz T, Sewer M, Jorge O, Garcia G, Reyes O, et al. Murine Monoclonal Antibodies specific for the HBsAg Determinant. Biotecnologia Aplicada 1995;12(2):89. 9. Peters KG, Rao PS, Bell BS, Kindman LA. Green fluorescent fusion proteins: Powerful tools for monitoring protein expression in live zebrafish embryos. Devel Biol 1995;171(1):252-257. 10. Hernandez O, Castro FO, Aguilar A, Uliver C, Perez A, Herrera L, de la Fuente J. Gene transfer in common carp (Cyprinus carpio L.) by microinjection into the germinal disc. Theriogenology 1991;35:625-632. 11. de la Fuente J, Martinez R, Estrada MP, Hernandez O, Cabrera E, Garcia del Barco D, et al. Towards growth manipulation in tilapia (Oreochromis sp.):generation of transgenic tilapia with chimeric constructs containing the tilapia growth hormone cDNA. Journal of Marine Biotechnology 1995;3:216-219. 12. Stuart GW, McMurray JV, Westerfield M. Replication, integration and stable germ-line transmission of foreign sequences injected into early zebrafish embryos. Development 1988;103:403-412. 13. Castro FO, Perez A, Aguilar A, de la Riva G, Martinez R, de la Fuente Jose, Herrera L. Expression of hepatitis B surface antigen in transgenic mice. Biotecnologia Aplicada 1989;3:251-257. 14. CLONTECH, Laboratories Inc. Aplication Notes (PT2040-1) 1996:43. 15. Kevin GP, Prema SR, Bridgit SB, Kindman A. Green fluorescent fusion proteins: Powerful tools for monitoring protein expression in live zebrafish embryos. Developmental Biology 1995; 171:252-257. 16. Stuart GW, Vielkind JR, McMurray JV, Westerfield M. Stable lines of transgenic zebrafish exhibit reproducible patterns of transgene expression. Development 1990;109:577-584. 17. Chelly J, Concordet JP, Kaplan JC, Kahn A. Illegitimate transcription: transcription of any gene in any cell type. Proc Natl Acad Sci USA 1989; 86:2617-2621. 18. Hernandez O, Attal J, Theron MC, Puissant C, Houdebine LM. Efficiency of introns from various origins in fish cells. Mol Mar Biol Biotechnol 1993;2:181-188 19. McEvoy T, Stack M, Keane B, Barry T, Sreenan J, Gannon F. The expression of a foreign gene in salmon embryos. Aquaculture 1988;68:27-38. 20. Yoon SJ, Hallerman EM, Gross ML, Liu Z, Schneider JF, Faras AJ, et al. Transfer of the gene for neomycin resistance into goldfish, Carassius auratus. Aquaculture 1990;85:21-33. 21. Tamiya E, Sugiyama T, Masaki K, Hirose A, Okoshi T, Krube I. Spatial imaging of luciferase gene expression in transgenic fish. Nucleic Acids Res 1990; 8:1070. Copyright 1996 Elfos Scientiae
The following images related to this document are available:Photo images[ba96106c.jpg] [ba96106b.jpg] [ba96106a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}