 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
TERCERA REGION VARIABLE DE LA GLICOPROTEINA EXTERNA DEL VIRUS DE LA INMUNODEFICIENCIA HUMANA TIPO 1. ESTRUCTURA, FUNCIONES BIOLOGICAS E INMUNOGENICIDAD Carlos A Duarte Division de Vacunas, Centro de Ingenieria Genetica y Biotecnologia, apartado postal 6162, Cubanacan, Ciudad de La Habana, C.P. 10600, Cuba. Telefono (53-7) 218008, 218466, extension 1656. Correo electronico: carlos.duarte@cigb.edu.cu
Code Number:BA97001
Size of Files:
Text: 131.8K
Graphics: Line drawings (gif) -6.5K
ABSTRACT More than six million of AIDS cases have been reported since the beginning of the HIV/AIDS pandemic. In addition, there were 21.8 millions HIV-1 infected persons by June 1996. In spite of recent improvements in the therapeutics, the development of a preventive vaccine is critical for the control of AIDS. The aim of this review is to present a profound view of one of the most attractive regions of the viral envelope: the V3 loop. In V3, the principal neutralizing domain of HIV-1 has been located, as well as epitopes for helper and cytotoxic T cells. Because of its exceptional features, it has been included in the vast majority of the vaccine candidates in evaluation. The main findings about the structure and polymorphism within this region are summarized. Despite its variability, certain structural parameters are conserved in order to permit the V3 loop to mediate important functions in the viral infection. A general hypothesis about the role of V3 loop in the events of viral entrance and tropism is advanced. Different studies demonstrate the existence of cross-reactivity among V3 peptides. Some human monoclonal antibodies recognize conserved epitopes in V3 from different clades and neutralize primary isolates. However, the relevance of these aspects for vaccine development is still uncertain. Finally, an overview of the main vaccine strategies involving V3 immunogens is presented. Key words: HIV-1, neutralization, polymorphism, monoclonal antibodies, vaccines, AIDS RESUMEN La infeccion por VIH-1/SIDA es una importante pandemia que afecta a toda la humanidad. Actualmente 21,8 millones de personas estan infectadas con el VIH y alrededor de 7 000 contraen la infeccion cada dia. Desde el inicio de la epidemia se han reportado mas de seis millones de casos de SIDA. A pesar de recientes avances en la terapeutica con drogas antivirales, es evidente que el desarrollo de una vacuna preventiva es la unica opcion para lograr el control de la pandemia. El objetivo del presente trabajo es abordar con mayor profundidad el estudio de una de las regiones mas fascinantes de la envoltura viral: el lazo V3. En V3 se localiza el dominio principal de neutralizacion del VIH-1 y a su vez coexisten epitopos para celulas T auxiliadoras y citotoxicas. Por estas caracteristicas excepcionales, esta region ha sido incluida en casi todos los candidatos vacunales que se evaluan actualmente. Se resumen los principales hallazgos en cuanto a la estructura y el polimorfismo de esta region. A pesar de su variabilidad, determinados parametros estructurales se conservan para permitir a la region V3 mediar importantes funciones en el proceso infectivo. Se esboza una hipotesis general acerca del papel de V3 en el proceso de penetracion viral, y se discute su influencia sobre el tropismo viral. Varios trabajos indican la existencia de reactividad cruzada entre peptidos de V3. Algunos anticuerpos monoclonales humanos contra V3 definen epitopos considerablemente conservados y han logrado neutralizar cepas primarias. Por ultimo se ofrece una panoramica de las diferentes estrategias vacunales basadas en inmunogenos de la region V3. Palabras claves: VIH-1, neutralizacion, polimorfismo, anticuerpos monoclonales, vacunas, SIDA Introduccion Han pasado trece anos desde que se aislo por vez primera el virus de la inmunodeficiencia humana (VIH) (1) y doce desde que este fuera caracterizado como el agente etiologico del sindrome de inmunodeficiencia adquirida (SIDA) (2). Mas de seis millones de casos de SIDA han ocurrido en el mundo desde el inicio de la epidemia hasta 1996, y se estima que 21,8 millones de personas son actualmente portadores del VIH. Cada dia se reportan 7 000 nuevas infecciones; el 98 % de estas en el mundo subdesarrollado (3).
Durante los primeros anos de la infeccion, la terapia antiviral, basada en la droga AZT, ha sido poco eficaz. En los ultimos tres anos se han desarrollado una docena de nuevas drogas antivirales inhibidoras de las enzimas virales reverso-transcriptasa y proteasa; en la clinica, estas drogas han mostrado ser mas eficaces que el AZT. Por otra parte, resultados recientes de ensayos clinicos con combinaciones de dos o tres de estas drogas han sido muy alentadores y han abierto una etapa de mayor optimismo en la terapia del SIDA. No obstante, es evidente que la terapia antiviral no es la solucion para el SIDA como pandemia, ya que el 94 % de las personas infectadas con el VIH viven actualmente en paises del tercer mundo, donde no hay acceso a los medicamentos mas elementales. Estas razones respaldan con fuerza la necesidad urgente de desarrollar una vacuna eficaz contra el SIDA, direccion en la que se vienen realizando intensos esfuerzos en todo el mundo.
A pesar de indiscutibles avances en el conocimiento de los aspectos basicos de la infeccion por VIH, las investigaciones se desarrollan en una atmosfera de gran incertidumbre. La falta de conocimiento de los indicadores de proteccion, la ausencia de un modelo animal que reproduzca la enfermedad y la dificultad adicional que impone el polimorfismo del virus son los tres obstaculos principales para el desarrollo de una vacuna.
Esta situacion ha provocado que se exploren una amplia gama de estrategias vacunales, entre ellas: vacunas atenuadas (4), subunidades recombinantes (5), vectores vivos (6-8), peptidos sinteticos (9) y ADN desnudo (10, 11). Algunas de estas estrategias estan representadas en los 24 candidatos vacunales evaluados en un numero considerable de ensayos clinicos de Fase I y II en humanos. Existen una serie de revisiones recientes que abordan estos aspectos (12-14). El objetivo del presente trabajo es abordar con mayor profundidad el estudio de una de las regiones mas fascinantes de la envoltura viral: el lazo V3.
Posiblemente no exista en otro virus una region que haya sido tan estudiada y que genere opiniones tan controvertidas. En V3, epitopos de union de anticuerpos neutralizantes coexisten con epitopos para celulas T auxiliadoras (Th) y citotoxicas (CTL). Por otra parte, es considerada una pieza clave en el proceso infectivo, ejerce una influencia importante sobre el tropismo viral y se especula ademas que desempena un papel protagonico en la patogenesis de la enfermedad. Por sus caracteristicas excepcionales, esta region ha sido incluida de una u otra forma en casi todos los candidatos vacunales que se evaluan actualmente. Epitopos para anticuerpos neutralizantes en el VIH-1 Con la excepcion de un reporte aislado de anticuerpos neutralizantes contra la proteina externa de la capsida p17 (15), los epitopos blanco de la accion de anticuerpos neutralizantes se localizan en la envoltura del virus. Esta esta formada por dos glicoproteinas: la gp41 o proteina transmembranica y la gp120 o proteina externa. gp41 Se han descrito dos sitios blanco de anticuerpos neutralizantes en la proteina transmembranica. El primero fue reportado por Chanh TC et al. (16) en la region 735-747 y confirmado a traves de experimentos de inhibicion de la actividad neutralizante de sueros humanos (17) y mediante la expresion de este fragmento en poliovirus (18). Broliden PA et al. (17) reportaron por primera vez la region conservada 647-671 como blanco de anticuerpos neutralizantes. Los peptidos de esta region son capaces de inhibir el efecto neutralizante de sueros humanos. Este hallazgo fue confirmado por la obtencion de un anticuerpo monoclonal (AcM) humano neutralizante que reconoce la secuencia ELDKWA en la posicion 662-667 (19). La inmunogenicidad de estos sitios en humanos parece ser debil ya que la mayoria de los anticuerpos contra gp41 estan dirigidos contra una region conservada inmunodominante y no son neutralizantes (20). gp120 Uno de los primeros reportes de epitopos de anticuerpos neutralizantes en la gp120 fue el de Ho D et al. (21), quienes generaron antisueros neutralizantes en conejos a traves de la inmunizacion con un peptido del segundo dominio conservado (aa 254-274). Este resultado fue confirmado para primates (22), aunque otros autores no fueron capaces de reproducirlo (23).
Tambien se han descrito anticuerpos neutralizantes en macacos contra las regiones 152-176, 193-218 y 206-230 de la gp120 (22). La primera de estas regiones se ubica entre los dominios V1 y V2 y sobrelapa con el peptido 164-187 tambien descrito como sitio de neutralizacion en la region V2 (24). Otros autores tambien han reportado generacion de anticuerpos neutralizantes con peptidos de V2 (25). AcMs de raton contra la region C4 neutralizan diferentes cepas de VIH-1 (26). Esta region, que forma parte del sitio de union al receptor, es inmunosilente en humanos. Por otra parte, se ha descrito la presencia de anticuerpos neutralizantes contra el extremo carboxilo terminal (498-508) en conejos (27) y en sueros humanos (17). No obstante, el principal dominio neutralizante (PND) de la envoltura del VIH-1 se localiza en la region V3.
Hasta aqui mencionamos epitopos secuenciales y poco dependientes de la estructura terciaria de la proteina. Sin embargo, diferentes trabajos han demostrado que existen epitopos neutralizantes de naturaleza conformacional en la envoltura (28, 29). Los anticuerpos que reconocen estos epitopos muestran un espectro neutralizante mas amplio que la mayoria de los anticuerpos contra V3 y otras regiones. Se considera que los bajos titulos neutralizantes contra diferentes cepas de VIH-1 observados en sueros humanos son mediados por este tipo de anticuerpos (30), e incluso otros experimentos sugieren que la mayor parte de la respuesta contra gp120 esta dirigida contra epitopos conformacionales (31). La tercera region variable del VIH-1 contiene el determinante principal de neutralizacion del VIH-1 En 1987 se publicaron los resultados de un estudio de prediccion de epitopos la envoltura del VIH-1 (32). Para esto se empleo un programa que integra diferentes parametros como hidrofilicidad, probabilidad de giros, accesibilidad y movilidad. Dentro de las regiones senaladas como probable epitopo B estaba la tercera region variable (V3) de la gp120.
Estas predicciones se verificaron independientemente por varios grupos de investigadores. En el primero de estos trabajos se describio la generacion de anticuerpos neutralizantes en chivos inmunizados con el peptido de dieciocho aminoacidos SP10 acoplado al toxoide tetanico (33). Este peptido represento la region V3 del aislamiento IIIB y no indujo anticuerpos neutralizantes contra las cepas MN y RF. Por otra parte, un peptido similar con la secuencia de la cepa RF, neutralizo solo a esta y no a las cepas MN o IIIB. Rusche JR et al. (34) reportaron la obtencion de antisueros contra gp120 recombinante producida en celulas de insectos y contra un fragmento de esta (PB1) obtenido en bacterias. Estos antisueros inhibieron la formacion de sincicios provocada por el VIH-1 in vitro. Su actividad inhibitoria fue a su vez completamente bloqueada por un peptido de 25 aminoacidos de la region V3.
Goudsmit J et al. (35) encontraron que sueros neutralizantes de chimpances infectados con VIH-1 eran capaces de reconocer el peptido de catorce aminoacidos de V3 IRIQRGPGRAVTIG. Sueros sin capacidad neutralizante no mostraron reactividad contra este peptido. Los residuos variables a ambos lados del centro GPGR resultaron criticos para la union de los anticuerpos.
Los trabajos citados, y otros posteriores, establecieron que en la region V3 esta ubicado el determinante principal de neutralizacion (PND) del VIH-1. Por ejemplo, la eliminacion de la region codificante para V3 del gen de la gp120 origino una proteina incapaz de inducir anticuerpos neutralizantes (36). Se demostro que solo ocho aminoacidos en la zona central pueden acomodar un epitopo neutralizante (37, 38). Los cinco aminoacidos PGRAF son antigenicamente activos frente a sueros humanos, aunque la avidez de la interaccion disminuye de forma significativa (39).
Existen elementos experimentales que indican que el PND se limita a una region de aproximadamente 15-20 aminoacidos en el centro del lazo V3. Un AcM dirigido contra la secuencia IGKIGNMRQ en la base carboxilo terminal del lazo no mostro actividad neutralizante (40).
Estos resultados permitieron identificar que el PND contiene epitopos poco dependientes de la estructura terciaria de la proteina. En terminos mas precisos, peptidos de esta region adoptan una conformacion que mimetiza con exito su propia estructura en la proteina nativa. Esta importante propiedad permite su manipulacion sin menoscabo de su integridad antigenica y manteniendo la capacidad de inducir una respuesta humoral neutralizante contra el VIH-1. Estructura y variabilidad del PND Estructura Las primeras predicciones de la estructura secundaria de esta region estimaron como muy probable la presencia de un giro beta en la zona central del lazo (41). La aplicacion de tecnicas de resonancia magnetica nuclear y dicroismo circular a peptidos de V3 corroboro la existencia de un giro de tipo II tanto en peptidos ciclicos de 42 aminoacidos como en peptidos lineales de 24 residuos (42). Estas observaciones indican que el giro beta no depende de la integridad del puente disulfuro y que existe un predominio de estructura secundaria al azar, aunque otros autores han encontrado una marcada tendencia a la formacion de helice alfa en la porcion C terminal (43). Evidencias mas precisas de la disposicion tridimensional se obtuvieron recientemente a traves del analisis cristalografico del complejo entre el fragmento Fab de los AcMs 59.1 y 50.1 y el peptido de V3 de la cepa MN (44, 45). La region central conservada mostro en efecto una estructura de giro tipo II (GPGR), pero ademas se encontro una conformacion general en forma de S, con la presencia de otros dos giros de tipo III y I (aminoacidos GRAF y RAFY, respectivamente) (Figura 1). La conformacion del peptido se estabiliza por tres puentes de hidrogeno internos entre los residuos G319-R322, G321-F324 y R322-Y325. Es muy interesante destacar que este peptido adopto una conformacion practicamente identica en su asociacion con estos dos anticuerpos monoclonales con diferentes caracteristicas de union (Tabla 1).
No obstante algunos trabajos sugieren la existencia de epitopos definidos por interacciones entre V3 y otros dominios de la gp120. Por ejemplo, cambios de aminoacidos fuera de V3 confirieron resistencia a la neutralizacion por un AcM contra V3 (62).
Se ha determinado que los aminoacidos de la secuencia 113-142 estan cercanos al V3 en la estructura tridimensional de la proteina y en su conjunto constituyen un epitopo discontinuo (63, 64). Mutaciones puntuales en estos epitopos afectan la neutralizacion del VIH-1 por anticuerpos contra V3. Adicionalmente se ha encontrado que la sustitucion de V3 en diferentes contextos de gp120 modula de forma importante la respuesta neutralizante contra el virus (65).
El conjunto de estas observaciones indica que no se puede subestimar el papel de la interaccion entre V3 y otros dominios de la gp120 en la definicion de epitopos de neutralizacion del VIH, aunque esto no esta en contradiccion con la existencia de epitopos en V3 que son independientes de estas restricciones estructurales.
--------------------------------------------------------------------------- Tabla 1. Anticuerpos monoclonales contra la region V3. --------------------------------------------------------------------------- AcM Origen Referencia Ka Epitopo Potencia --------------------------------------------------------------------------- 0,5beta murino 46 2 x 10^-8 RGPGRAFVTIGKIG S ID50 1 ug/mL 9248 murino 47 3 x 10^-8 NNTRKSIRIQRG S ID50 25 ug/mL BAT123 murino 48 - RIGRGPGRAFVTIGK N ID90 1 ug/mL P4D10 murino 49 - IQRGPGRAFV - N701-9B humano 50 - RIHIGPGRAFY N ID50 0,9 ug/mL 5025A humano 51 - GRAVTIG N ID90 10 ug/mL 19b humano 50, 52, 53 - I----G--FY-T - NM-01 murino 54 - GPGR N ID50 1 ug/mL 257-20 humano 55 4 x 10^-6 KRIHI N ID50 3 ug/mL 268-110 humano 55 8 x 10^-7 HIGPGR N ID50 23 ug/mL 447-D humano 56 - GPGR NID90 <10ug/mL Loop 2 humano 57 - SISGPGRAFYTG - M77 murino 58 - IRIGRGPGRAFVTI N ID90 0,3 ug/mL 50-1 humano 59 3 x 10^-5 GPGRAF N S ID90 25 ug/mL 83-1 humano 59 3 x 10^-7 HIGPGRAF N S ID90 2,5 ug/mL 58-2 humano 59 1 x 10^-8 IXIGPGR N S ID90 0,25 ug/mL 10F10 murino 60 x 10^-8 IHIGP-R N ID50 1 ug/mL 2C4 murino 60 x 10^-8 IHIG--R N ID50 1 ug/mL 694/98D humano 61 - GRAF - IS) inhibicion de la formacion de sincicios. N) inhibicion de la produccion de p24 viral. ID50(90)) concentracion de anticuerpo necesaria para reducir en un 50 (90) % la infeccion por el virus. --------------------------------------------------------------------------- Polimorfismo genetico El polimorfismo genetico del VIH, principalmente de las proteinas de la envoltura, es muy elevado. Esto viene dado por la baja fidelidad de copia de la reverso transcriptasa viral (RT) que introduce aproximadamente un error por cada 700 a 10^4 bases incorporadas (66). No obstante esta tasa de mutaciones no es diferente de la de otros retrovirus. Experimentos recientes sobre la dinamica de la infeccion por VIH indican que la coexistencia de un numero tan elevado de variantes virales (cuasiespecies) en el organismo infectado es consecuencia, en primer termino, de la alta velocidad de replicacion del virus. Segun calculos de dos grupos independientes se generan diariamente 10^9 particulas virales (67, 68).
El estudio de la variabilidad genetica de la region V3 puede dividirse en tres etapas bien definidas. La primera se enmarca entre la aparicion de la primera secuencia del genoma de VIH-1 (69) y la publicacion del trabajo de La Rosa GJ et al. (41). En esta etapa se secuencio el V3 de una veintena de cepas en su gran mayoria provenientes de Europa y Estados Unidos (Tabla 2).
--------------------------------------------------------------------------- Tabla 2. Secuencia de la region central de V3 de16 cepas de laboratorio de VIH-1. --------------------------------------------------------------------------- Denominacion Subtipo Secuencia --------------------------------------------------------------------------- US.CDC42 B KRVTLGPGRVWYTT ZR.NDK D QRTSIGLRQSLYTI US.BRVA B KRITMGPGRVYYTT FR.LAI B IRIQRGPGRAFVTI HT.RF B KSITKGPGRVIYAT US.BAL B KSIHIGPGRAFYTT US.SC B RSIHIGPGRAFYAT US.SF162 B KSITIGPGRAFYAT US.SF2 B KSIYIGPGRAFHTT US.NY5CG B KGIAIGPGRTLYAR US.JRSCF B KSIHIGPGRAFYTT US.MN B KRIHIGPGRAFYTT ZR.MAL D RGIHFGPGQALYTT ZR.ELI D QRTPIGLGQSLYTT ZR.JY1 D QSTPIGLGQALYTT ZR.Z2Z6 D QRTSIGLGQALYTT --------------------------------------------------------------------------- El citado trabajo de La Rosa GJ et al. (41) fue el primer reporte extenso sobre la variabilidad del V3. En este se describio la composicion de la region V3 en 245 aislamientos, principalmente de Estados Unidos. Las conclusiones de este trabajo fueron estimulantes para los defensores de la importancia del V3 como candidato vacunal, pues se observo que el polimorfismo no era tan grande como se estimo a partir de los primeros datos de secuencia. Las sustituciones de aminoacidos encontradas fueron generalmente conservativas y se confirmo que la porcion central del lazo, fundamentalmente el tetrapeptido GPGR, es la mas conservada. La tercera etapa se caracteriza por una explosion informativa sobre secuencias de esta region que han sido recogidas en la recopilacion de datos de secuencias de nucleotidos y aminoacidos de retrovirus publicados por el laboratorio de Los lamos (70). Esta base de datos que se actualiza anualmente contiene la informacion de cepas circulantes en diversas areas geograficas. Un aporte importante a este conocimiento lo ha brindado el programa de estudios de polimorfismo viral que ha conducido en los ultimos anos la OMS en Brasil, Ruanda, Uganda y Tailandia como preparacion para el desarrollo de futuros ensayos de eficacia de vacunas.
Actualmente los aislamientos de VIH-1 se clasifican en dos grandes grupos de acuerdo a la secuencia de ADN del gen de la envoltura. El grupo M o grupo principal se divide a su vez en ocho "clades" o subtipos desde el A hasta el H, mientras que el O ("outliers") reune un pequeno grupo de secuencias que divergen considerablemente del grupo principal. La Tabla 3 muestra las secuencias consensos de la region central de V3 para cada uno de esos subtipos. Sin embargo, dentro de estos subtipos existe heterogeneidad en la secuencia de aminoacidos de la region V3, y por otra parte, cepas que clasifican en diferentes subtipos presentan considerable homologia dentro de V3. Por su importancia en el desarrollo de vacunas Korber y Myers (71) han propuesto un sistema de clasificacion paralelo basado en la secuencia de aminoacidos de la region central de V3. Este sistema es denominado fenotipico y agrupa las secuencias de V3 en 20 grupos. Hasta el momento no se ha estudiado detalladamente el grado de reactividad cruzada entre peptidos de diferentes grupos. Es muy probable que ni aun la clasificacion fenotipica responda a un criterio inmunologico funcional de reactividad cruzada entre cepas. En Cuba se ha venido desarrollando en los ultimos tres anos un estudio genetico y virologico de las cepas circulantes. Los resultados obtenidos con mas de 40 secuencias de la region C2-V3 indican la coexistencia de cepas de subtipos B y C (72, 73). Los estudios longitudinales realizados demuestran que el polimorfismo en la secuencia de VIH-1 aumenta dentro de cada individuo (74, 75) y la region V3 no es una excepcion. Se ha descrito que la heterogeneidad de V3 puede aumentar hasta el 19,5 % entre clones de un mismo paciente (76). Al estudiar la secuencia de V3 en seis ninos infectados por transfusion y la del donante de plasma, se encontraron importantes diferencias entre los infectados. La distancia entre los aislamientos virales de estos y la cepa original aumento con el tiempo y esta variacion fue dependiente del hospedero, rapida e independiente del nivel de antigeno expresado (77). La aparicion de nuevas variantes en la region V3 y la relacion observada entre la respuesta de anticuerpos contra V3 y la diversidad de secuencia en individuos infectados (78) refleja la existencia de una fuerte presion selectiva del sistema inmune. Adicionalmente, segun se ha discutido antes, mutaciones en otras regiones de la envoltura pueden provocar cambios que afectan la conformacion de V3 y sus interacciones con otros dominios de la gp120 y le permiten evadir la respuesta inmune del hospedero (79, 80). Sin embargo, a pesar de que la variacion dentro de V3 de cada individuo aumenta con el tiempo, la secuencia consenso apenas sufre variaciones en un periodo de diez anos (81). Por otra parte, se observa una tendencia a conservar el caracter (tipo particular y posicion) de los aminoacidos de esta region (41). Esto es, probablemente, el reflejo de una presion selectiva dirigida a conservar una estructura especifica en el V3, e indica que este debe desempenar una funcion biologica importante para el virus. En resumen, la composicion de aminoacidos de V3 parece ser la resultante de dos fuerzas. Por una parte, la respuesta inmune del hospedero que tiende a seleccionar mutantes de escape, y por otra, el propio papel de V3 en el proceso infectivo que tiende a conservar las variantes mejor adaptadas para la replicacion exitosa. Polimorfismo antigenico. Serologia dirigida con peptidos de V3 Los anticuerpos anti V3 en sueros humanos aparecen en los primeros seis meses despues de la seroconversion y la subclase predominante en los sueros humanos es IgG1 (38). La relacion entre la serologia con peptidos de V3 y la secuencia de aminoacidos de las cepas circulantes ha sido objeto de controversia en el mundo. Los anticuerpos contra el V3 de la cepa MN son predominantes no solo en sueros de Estados Unidos y las Americas (95 %) sino tambien en otros paises de frica Occidental (80 %) como Zaire y Zimbawe (82, 83). Por el contrario sueros provenientes de frica Oriental muestran solo un 30 % de reactividad contra MN. Esta observacion no concuerda con los datos de secuencia del ADN viral, ya que el V3 de los aislamientos de frica es muy diferente al del MN (71).
Carrow EN et al. (82) encontraron mayor frecuencia de respuesta en sueros africanos contra un peptido de la cepa MN que contra un peptido de la cepa africana Z3. No obstante, la secuencia de V3 de Z3 difiere bastante del consenso de los grupos circulantes en la region, por lo que es muy cuestionable su empleo como prototipo de las cepas africanas. Se argumenta que la elevada frecuencia de aparicion de anticuerpos contra el V3 consenso de America y Europa en frica puede ser indicativa de un origen africano para este tipo de aislamiento (83). No obstante, esta hipotesis no parece ser convincente para explicar la ausencia de esta secuencia en los virus aislados en frica.
Una explicacion alternativa es la existencia de reactividad cruzada entre cepas de frica (subtipos A, C y D) y America (subtipo B). En este sentido se ha descrito que sueros de individuos infectados con virus del subtipo B reconocen peptidos consenso del subtipo D y viceversa (84). En nuestro laboratorio se logro inhibir la reactividad de sueros contra el peptido consenso B (KSIHIGPGRAFYATG) con peptidos de cepas caracteristicas de frica (KSIRIGPGQAFYATG) (85). Adicionalmente, en Etiopia se reporto un 39 % de sueros con reaccion cruzada entre los subtipos A y C (86), mientras que otros investigadores tambien han encontrado alta frecuencia de reactividad cruzada entre peptidos de la misma area geografica (87).
A pesar de que la existencia de reactividad cruzada incrementa la ambiguedad de los resultados, se ha demostrado la utilidad potencial de la serologia para la tipificacion de las cepas circulantes dentro de una region determinada. Por ejemplo, en Tailandia fue posible discriminar por serologia entre individuos infectados por virus de los subtipos B y E, con un 93 % de coincidencia entre serotipaje y genotipaje (88, 89). En un estudio similar, con sueros de diferentes areas geograficas, se encontro un 90 % de coincidencia entre la secuencia de ADN y el serotipo (90). Esto fue posible, a pesar de la existencia de sueros con reactividad cruzada, por la aplicacion de un ELISA en condiciones limitantes de antigeno en el recubrimiento. Este ensayo favorece la deteccion de anticuerpos de mayor afinidad y por tanto menos polireactivos. No obstante, en otros estudios con sueros provenientes de cuatro paises diferentes no se encontro una correlacion tan directa entre genotipo y fenotipo (91).
--------------------------------------------------------------------------- Tabla 3. Secuencias consenso de la region central de V3 de los subtipos de VIH-1. --------------------------------------------------------------------------- Subtipo Secuencia -------------------------------------------------------------------------- A RKSVRIGPGQAFYAT B RKSIHIGPGRAFYTT C RKSIRIGPGQTFYAT D RGRTHIGPGQALYTT E RTSITIGPGQVFYRT F RKSIHLGPGQAFYAT G RKSITIGPGQAFYAT H RKSIRIGPGQAFHAI 0 VQEIRIGPMAWYSMG ---------------------------------------------------------------------------
El conjunto de estos resultados sugiere que solo en determinadas areas epidemiologicas donde coexistan cepas virales divergentes en la region V3 puede ser de utilidad la serologia, ya que donde haya promiscuidad de cepas relacionadas antigenicamente la existencia de reactividad cruzada puede hacer imposible la tipificacion por esta via. La seleccion de los peptidos adecuados para cada region, asi como la metodologia empleada, parecen determinantes para lograr un resultado exitoso.
Tambien se reporta la existencia de sueros "no tipables" que no reconocen a ninguno de los peptidos empleados. Estos individuos pueden haber sido infectados con variantes virales muy alejadas antigenicamente de los peptidos usados en el tipaje. Por ejemplo, el suero de un individuo infectado en Cuba con una cepa del grupo B con la secuencia atipica en el V3 GRGR no reconocio ninguno de los peptidos de V3 que se evaluaron, pero si al peptido homologo (73).
A pesar de la vigorosa respuesta humoral desarrollada desde fases tempranas, no se logra contener la evolucion de la enfermedad. Se argumenta que el sistema inmune del hospedero no es capaz de adaptarse a la variacion sostenida de las principales regiones antigenicas del virus. Se ha comprobado que en los primeros tiempos despues de la seroconversion, la especificidad de los anticuerpos anti V3 coincide con la secuencia de ADN encontrada para cada individuo. En individuos estudiados cinco anos despues de la seroconversion el patron de respuesta humoral contra el V3 no difiere generalmente del anterior, sin embargo, el tipo de secuencia predominante en el virus varia (92).
Nara y Goudsmit (93) acuden a la hipotesis conocida como "Original Antigenic Sin" (OAS) para explicar esta observacion. Segun esta teoria algunos antigenos de naturaleza variable como el V3 predeterminan el comportamiento del sistema inmune, de forma que al ponerse en contacto con una nueva variante se estimulan preferentemente los clones de celulas B de memoria cuyos receptores de superficie reconocen las regiones comunes a ambas variantes. Segun estos autores, esto "congela" de cierta forma el repertorio de inmunoglobulinas generado, obstaculizando su diversificacion. Objeto de debate ha sido tambien la relacion entre anticuerpos anti V3 y la evolucion a SIDA. Segun Fenouillet E et al. (94) la presencia de anticuerpos de alta afinidad contra peptidos de V3 correlaciono con el estadio clinico del paciente. Resultados semejantes fueron encontrados en pacientes cubanos, y adicionalmente, tambien se evidencio correlacion entre anticuerpos antipeptidos V3 y la progresion a la enfermedad (85). Estos resultados fueron confirmados empleando polipeptidos multiepitopicos que incluyen varias regiones V3 (Duarte C, resultados no publicados). Sin embargo Holmback K et al. (87) no encontraron correlacion entre los anticuerpos contra peptidos de V3 autologos y el estado clinico. La explicacion para esta discrepancia puede estar en que en este estudio, a diferencia de los anteriores, solo se evaluo la respuesta anti V3 de forma cualitativa.
El papel de los anticuerpos contra V3 en la transmision materno-fetal del VIH-1 ha sido muy discutido en la literatura. Se ha reportado que la presencia de anticuerpos contra un peptido de la base de V3 en madres infectadas correlaciona con la ausencia de transmision a sus hijos (95). Posteriormente Devash Y et al. (96) encontraron correlacion entre la presencia de anticuerpos de alta afinidad para V3 en madres y la ausencia de transmision del virus a los recien nacidos. Sin embargo, otros autores no han sido capaces de corroborar esta observacion (97-99). El hallazgo, por estos ultimos autores, de anticuerpos contra variantes de V3 en el suero del recien nacido y no en el de su madre, sugiere que la infeccion es causada por mutantes que escapan a la respuesta inmune materna.
Los resultados de serologia con peptidos sinteticos deben ser analizados con precaucion, pues la metodologia varia de forma importante de laboratorio en laboratorio y se ha comprobado que estas variaciones pueden provocar diferencias en los resultados obtenidos. Por ejemplo, la inclusion de Tween 20 en el diluyente de las muestras amplia la reactividad de los anticuerpos con peptidos de V3 de diferentes cepas y no siempre la reactividad de AcMs contra peptidos de V3 en ELISA predice satisfactoriamente su capacidad de union a gp120 (100).
Segun la experiencia de nuestro laboratorio la presentacion de los peptidos acoplados a seroalbumina bovina mejora significativamente la capacidad de deteccion de sueros humanos (85). Este procedimiento evita la perdida de epitopos producida por la interaccion de residuos importantes con el plastico. Funciones Biologicas de V3 V3, infeccion viral y tropismo El V3 es importante para la entrada viral y la formacion de sincicios
El VIH penetra en la celula blanco a traves de un proceso directo de fusion entre la membrana viral y la membrana citoplasmatica. Este mecanismo de fusion, independiente del pH del medio, es analogo a la formacion de sincicios que tiene lugar entre las celulas infectadas y no infectadas. Diversos estudios indican que V3 juega un papel importante en ambos eventos.
Cuando se mutan las cisteinas que forman el puente disulfuro que define el lazo V3, o se elimina completamente esta region, la proteina gp160 no sufre procesamiento proteolitico ni es capaz de enlazar CD4 y el virus resultante no es infectivo (101-103). Esto demuestra que la existencia de este lazo es esencial para la conformacion tridimensional correcta y funcional de las proteinas de la envoltura.
Por otra parte, la delecion de fragmentos de V3 (103, 104), la insercion de cinco aminoacidos (105), y mutaciones puntuales en esta region (106-111), no alteran el procesamiento de la gp160 ni su capacidad de union a CD4, pero si la infectividad del virus y su propiedad de inducir la formacion de sincicios.
Los aislamientos de VIH-1 se han clasificado de acuerdo a su fenotipo y tropismo en dos grupos: (a) monocitotropicos (MT), no inductores de sincicios (NSI) y de baja capacidad replicativa (SL) y (b) linfocitotropicos (LT), inductores de sincicios (SI) y de alta capacidad de replicacion (RH). La aparicion de cepas SI ha sido asociada a la progresion a SIDA (112-115). El reemplazo de V3 de una cepa SI a una NSI le confirio la capacidad de inducir sincicios y viceversa (100).
Se ha especulado que el caracter basico de V3 es lo que le permite interactuar con moleculas negativas en la superficie celular para establecer una primera interaccion que facilite el encuentro con el receptor CD4 (117). Para apoyar esta hipotesis existen datos experimentales que demuestran la interaccion de peptidos de V3 con sulfato de dextran (118), sulfato de heparan (119) y heparina (120).
Adicionalmente se ha demostrado la importancia del numero y la posicion de los residuos positivos en V3 en el fenotipo viral (121-126). En particular la presencia de residuos basicos en las posiciones 306 y 320 esta fuertemente asociada al fenotipo de SI y RH (116). V3 y tropismo
En los ultimos anos se ha evidenciado que el tropismo del VIH-1 es mas complejo que lo que inicialmente se penso. Ademas de linfocitos T4, los monocitos y otras celulas del sistema reticulo endotelial que presentan el receptor CD4, se ha demostrado que el virus infecta de celulas de origen nervioso y epitelial.
El tropismo por linfocitos o monocitos es muchas veces excluyente en las cepas de VIH-1 y fue asociado inicialmente con una region de la envoltura que incluye V3 (127, 128). Mas tarde, se preciso que el intercambio de un fragmento de 20 aminoacidos de la region V3 entre la cepa linfocitotropica IIIB y la monocitotropica BaL (Tabla 4) es suficiente para revertir ambos fenotipos (129).
--------------------------------------------------------------------------- Tabla 4. Fragmento de V3 que definio el tropismo de las cepas BaL y IIIB de VIH-1. --------------------------------------------------------------------------- Cepa Secuencia AA+ AA- -------------------------------------------------------------------------- BaL SIHI..GPGRALYTTGEIIGDI 2 1 IIIB K-R-QR-----F--I-K.--NM 4 0 AA+): numero de residuos con carga positiva. AA-): numero de residuos cargados negativamente. -): igual aminoacido. .): no existe residuo en esa posicion. --------------------------------------------------------------------------- Otros cambios dentro de V3 tambien afectan el tropismo por monocitos (130- 133) o celulas nerviosas (134, 135). Finalmente virus mutantes en los cuales la prolina en posicion 313 del V3 fue sustituida por alanina son capaces de infectar las lineas MOLT4, CEM y SUPT1, de origen T, pero no la linea de origen B AA5 (109).
Estos resultados demuestran que la secuencia de V3 define el tropismo del VIH por diferentes sustratos celulares. Papel de V3 en el proceso infectivo Se conoce que el V3 no forma parte del sitio de union con alta afinidad al receptor CD4. ¨Cual es entonces su papel en el proceso de penetracion del virus? A continuacion se enumeran algunos resultados que pueden ayudar a aclarar este aspecto.
A partir de estos resultados se ha propuesto que el corte de V3 por una ectoproteasa de la membrana celular es un paso necesario para la entrada viral. Esta escision induciria un cambio de conformacion de la gp120 de forma tal que quede expuesto el fragmento hidrofobico N terminal de la gp41 que interviene directamente en el mecanismo de fusion de membranas. 2. Diferentes autores coinciden en que peptidos sinteticos de la region V3 son capaces de unirse a CD4 y a un peptido del dominio D2 de esta molecula (142, 143). El dominio D2 de CD4 no participa en la principal interaccion con gp120 y ha sido asociado con el proceso de fusion de membranas (144-146). Cambios puntuales de aminoacidos en V3 aumentaron la resistencia del virus a la neutralizacion por CD4 soluble (147). El significado biologico de esta interaccion no esta claro, ya que mientras peptidos de V3 potenciaron la infectividad del VIH a traves de un mecanismo dependiente de CD4 (148) otros estudios han reflejado un efecto completamente opuesto (149). Adicionalmente, peptidos de seis o catorce aminoacidos ramificados alrededor de un nucleo de lisinas, (Multi-Antigenic-Peptides, MAPs) demostraron capacidad de union a CD4 en linfocitos y macrofagos y efecto inhibitorio de la formacion de sincicios (150-153). 3. Existen evidencias que sugieren que V3 participa en un evento previo a la union de alta afinidad gp120- CD4 y posiblemente de corta duracion, ya que el virus necesita menos tiempo para escapar de los anticuerpos contra V3 que de anticuerpos contra CD4 (154). Estos resultados resultan dificiles de conciliar con el hecho de que V3 participe en la fusion de membranas pues este evento debe ocurrir necesariamente despues de la union al receptor. Es posible que estos datos experimentales esten viciados por las caracteristicas particulares de los AcMs empleados, por lo que deberian ser comprobados empleando otros anticuerpos. Es posible tambien que el virus escape mas rapido del AcM anti V3 porque esta region quede "inaccesible" a los anticuerpos despues de la interaccion entre gp120 y CD4.
Tomando en cuenta estas observaciones, junto a los elementos previamente expuestos que relacionan V3 con el tropismo, es posible proponer la siguiente hipotesis generalizadora:
2. Despues de ser "atraido" a la celula el virus se asocia fuertemente a CD4 a traves de la interaccion de alta afinidad. La region V3 queda protegida. 3. V3 interactua con otras moleculas de membrana, ya sean ectoproteasas, CD26, el dominio D2 del propio CD4 u otra molecula no identificada. Como resultado de esta interaccion ocurren cambios conformacionales importantes que bien pudieran involucrar un corte proteolitico de V3 que facilitan la exposicion del dominio fusogenico N terminal de gp41 y su insercion en la membrana y asi provocar la fusion.
En este tipo de mecanismo el lazo V3 desempenaria una doble funcion y la abundancia relativa de sus ligandos en la celula blanco dictaria la susceptibilidad de esta a la infeccion por una u otra cepa viral.
Finalmente, el esfingolipido galactosil ceramida (Gal-Cer) ha sido descrito como receptor alternativo para el VIH-1 en celulas de tejido epitelial (155) y nervioso (156, 157). Adicionalmente se demostro que peptidos de V3 tambien pueden unirse a esta molecula y bloquear la infeccion por VIH, inhibiendo directamente su interaccion con gp120 (158). Inmunidad Humoral Contra V3 Anticuerpos monoclonales contra V3
AcMs murinos y humanos contra V3. Epitopos y caracteristicas generales Diferentes AcMs murinos y humanos han sido generados contra la region V3 de diferentes cepas de VIH-1 (Tabla 1). La actividad biologica de estos anticuerpos ha confirmado la importancia del V3 como epitopo principal de neutralizacion.
A traves del empleo de peptidos con sustituciones de algunos aminoacidos se ha definido cuales son los residuos claves en la interaccion con el AcM. Estos aminoacidos pueden ser importantes, no solo en la interaccion con las regiones determinantes de la complementariedad (CDR) de los anticuerpos, sino tambien en la estabilizacion de la estructura del peptido a traves de las interacciones debiles entre sus cadenas laterales.
El caracter restringido del efecto neutralizante de la mayoria de los AcMs generados (46-50, 55, 58, 60), hizo que se catalogara de forma general a los AcMs contra V3 como tipos especificos o cepas especificas. No obstante, este concepto ha tenido que ser revisado con la aparicion de AcMs dirigidos contra las regiones mas conservadas (51, 54, 56, 59), los cuales muestran un patron de neutralizacion de cepas mucho mas amplio.
Algunos anticuerpos monoclonales generados contra la region V3 del subtipo B reaccionaron en ELISA con la gp120 del subtipo F, y en un caso, con varios aislamientos del subtipo E, pero fallaron en reconocer cepas primarias de los demas subtipos (52). El AcM 19b mostro amplia reactividad contra gp120 de los subtipos A, B, C, E y F pero solo logro neutralizar los del subtipo B y debilmente los del C (159). Esto demuestra que la actividad de union a peptidos no siempre es un buen indicador de actividad neutralizante, aunque se ha correlacionado la constante de disociacion (Kd) de la interaccion entre cada AcM y su peptido con su capacidad neutralizante (160).
Otras funciones mediadas por AcMs contra V3
Se ha comprobado que algunos de estos AcMs no solo presentan accion neutralizante de la infeccion por virus libre y de la inhibicion de la formacion de sincicios, sino que ademas son capaces de mediar in vitro la citotoxicidad celular dependiente de anticuerpos (ADCC) (49) y la virolisis por fijacion de complemento (161-162). Estos mecanismos pueden jugar un papel importante en la destruccion de celulas infectadas y particulas virales, respectivamente. Sinergismo de la accion entre AcMs
Diferentes autores han demostrado que la combinacion de AcMs contra V3 con otros AcMs desarrolla una actividad neutralizante superior a la simple adicion de los efectos individuales.
AcMs humanos dirigidos contra el V3 (447-D) y la region de union a CD4 (588-D) mostraron efecto sinergico (163). Sueros de individuos inmunizados con vaccinia-gp160 y gp160 soluble potenciaron la neutralizacion mediada por un AcM contra el sitio de union al CD4 (164). La actividad neutralizante de estos sueros humanos fue inhibida totalmente por incubacion con peptido de V3, lo cual sugiere que los anticuerpos neutralizantes estan dirigidos esencialmente contra esta region.
De forma opuesta se ha descrito que dos anticuerpos dirigidos contra el lazo V3 pueden establecer relaciones de competencia entre ellos. De esta manera un anticuerpo de menor capacidad neutralizante (9284) atenua el efecto de uno mas potente (0,5beta) (47). Una posible explicacion a este fenomeno puede ser la modulacion, provocada por la union del AcM 9284, de la conformacion del lazo, de forma tal que se oculte el epitopo reconocido por 0,5beta. Anticuerpos de segunda generacion
El empleo de anticuerpos monoclonales contra V3 en la inmunoterapia pasiva hasido un objetivo de diferentes investigadores. Son conocidas las limitacionesque presentan los AcMs murinos para la clinica, como por ejemplo la induccionde respuesta inmune y la incapacidad para mediar algunas funciones biologicas. Por otra parte, si bien se ha logrado obtener AcMs humanos, estosclones son generalmente inestables, de pobre crecimiento y baja secrecion deanticuerpos, lo que dificulta su produccion industrial. La sustitucion mediante manipulacion genetica de las regiones constantes murinas por humanasha permitido la obtencion de AcMs quimericos, los cuales conservan laafinidad y la actividad neutralizante del AcM original y a la vez incorporanotras funciones, como por ejemplo la capacidad para mediar la citotoxicid addependiente de anticuerpos (ADCC) (48, 49, 165). La capacidad (mostrada por fragmentos Fab2 (57) y Fab (166) de AcMs contra V3) de neutralizar la infeccion confirma que este proceso es independiente delas funciones biologicas del fragmento Fc o de la facultad de unir simultaneamente dos moleculas de gp120 en la superficie de un virion, o dosparticulas virales independientes (cross-linking). Incluso, un peptidos intetico de la tercera region determinante de la complementariedad (CDR3)del AcM F58 neutralizo la infeccion por la cepa LAI aunque con mucho menor eficiencia que el anticuerpo completo (167).
La mas reciente tendencia en el desarrollo de anticuerpos monoclonales es sin duda la tecnologia de obtencion de fragmentos Fab a traves de la construccion de librerias de genes variables en fagos. Se ha reportado la obtencion y caracterizacion de Fabs de mediana afinidad contra epitopos conformacionales de la gp120 y contra la region V3 utilizando este sistema (57, 168, 169). Recientemente se logro incrementar la afinidad y el rango de neutralizacion de un Fab contra el dominio de union al CD4 mediante mutaciones en los CDR (170). Neutralizacion de cepas primarias
El ensayo de neutralizacion es un indicador de la capacidad de determinados anticuerpos de inhibir la infeccion de celulas susceptibles in vitro. Hasta ahora nos hemos referido a ensayos que utilizan como blanco lineas celulares CD4+ y cepas adaptadas a replicarse en estas condiciones (cepas de laboratorio). No obstante, en los ultimos anos se ha encontrado que los anticuerpos que muestran actividad neutralizante en estas condiciones no son capaces de neutralizar aislamientos primarios (AP), replicados exclusivamente en celulas mononucleares de sangre periferica (PBMC). Estos aislamientos son fundamentalmente de fenotipo MT, NSI, SL. Vancott TC et al. (171) depletaron de anticuerpos contra V3 el suero de tres individuos infectados y encontraron que estos juegan un papel mas importante en la neutralizacion de cepas de laboratorio que en la de aislamientos primarios. En este estudio se empleo un ensayo de reduccion del titulo viral y su principal limitacion es el bajo numero de sueros y aislamientos primarios utilizados.
Se ha detectado con baja frecuencia actividad neutralizante de AP en sueros de personas infectadas y en varios AcMs humanos (172). En relacion con los AcMs contra V3 el primer reporte de neutralizacion de AP fue con el AcM 447D (173). En otro estudio realizado en varios laboratorios se ha reportado que el AcM 257-D neutralizo un aislamiento primario en tres de diez laboratorios en los cuales se evaluo, mientras el AcM TH1 solo fue neutralizante en uno de diez laboratorios (172). Esto indica que existe un margen alarmante de variabilidad entre laboratorios para esta tecnica. De acuerdo con los resultados reportados por Bou-Habib DC et al. (174), la region V3 esta poco expuesta en cepas primarias y esto las hace mas resistentes a la neutralizacion.
Es muy interesante el hecho de que solo anticuerpos humanos, ya sean monoclonales o policlonales, han mostrado la cualidad de neutralizar cepas primarias. Esto puede significar que la presentacion natural de la gp120 en el virus es determinante, y hasta ahora no se ha conseguido mimetizar satisfactoriamente en los distintos inmunogenos evaluados en animales y en seres humanos. El significado biologico de estos resultados es objeto de amplia discusion actualmente. Recientemente se ha demostrado que las celulas activadas con mitogenos ("blastos"), usadas en este tipo de ensayo, son mucho mas susceptibles a la infeccion con VIH que los linfocitos en reposo. La actividad neutralizante de sueros humanos frente a cepas primarias fue considerablemente mayor en un ensayo alternativo, en el cual las celulas no fueron activadas con fitohemaglutinina (PHA) (175). La capacidad de este ensayo para predecir la actividad in vivo de los anticuerpos ha sido puesta en entredicho cuando chimpances inmunizados con gp120 resistieron un reto con esta cepa multiplicada en PBMC, aun cuando sus sueros no habian logrado neutralizar previamente la misma cepa in vitro (176).
Facilitacion de la infeccion mediada por anticuerpos
Otro de los elementos que generan preocupacion en torno a la respuesta humoral contra la envoltura del VIH es el reporte por varios autores de la facilitacion de la infeccion mediada por anticuerpos (antibody dependent enhancement, ADE) (177, 178). Este proceso involucra los receptores de complemento y FC y tiene un efecto modesto en comparacion al descrito para otros virus como el dengue. Sus posibles implicaciones in vivo aun estan por demostrar. Estudios realizados con sueros de individuos vacunados con gp160 no han logrado demostrar actividad ADE (179).
Algunas regiones dentro de la envoltura viral se han definido como blanco de este tipo de anticuerpo (180, 181). Se ha reportado que anticuerpos contra V3 pueden mediar ADE en cepas divergentes (182, 183). Estos experimentos fueron realizados con peptidos que cubren toda la region V3 y es posible ofrecer dos hipotesis alternativas para explicar este resultado:
2. El efecto ADE es modulado por la afinidad de la interaccion entre cada anticuerpo y su epitopo. Segun esta, dos anticuerpos contra el mismo epitopo pueden mediar funciones opuestas en dependencia de su afinidad. En este caso, los anticuerpos generados contra el V3 de una cepa muestran menos afinidad contra cepas divergentes y su efecto neutralizante se trueca en ADE. Sera necesario acumular mas conocimientos sobre el mecanismo de ADE a nivel molecular para poder comprender cual es su verdadero papel en la infeccion por VIH-1. Hasta el momento los expertos estiman que las evidencias encontradas no son suficientes como para invalidar los estudios clinicos efectuados actualmente con diversos candidatos vacunales.
Inmunoterapia pasiva con AcMs contra V3 Modelos animales. Diferentes modelos de inmunoterapia pasiva han demostrado la eficacia de anticuerpos contra V3 para proteger contra el reto viral in vitro e in vivo. El anticuerpo monoclonal BAT123 confirio proteccion en el modelo del raton con Inmunodeficiencia Severa Combinada (SCID), repoblado con 20 x 10^6 celulas mononucleares de sangre periferica (CMSP) humanas ante un reto con la cepa IIIB. El reto se efectuo una hora despues de administrada una dosis de 40 mg del AcM por kilogramo de peso. Segun los estudios farmacocineticos, esta cantidad garantizo una concentracion en sangre de 100 g/mL del AcM, el doble de la concentracion necesaria para lograr neutralizacion completa in vitro. Se logro aislar VIH en cultivo de cinco de los seis controles y no se aislo en los doce ratones tratados. El tiempo de vida media del anticuerpo fue entre nueve y doce dias (186).
Por otra parte, el AcM Cbeta1, version quimerica del 0.5beta, logro proteger chimpances del reto con VIH-1. La administracion del AcM se efectuo tanto a una como a 24 h antes del reto. La dosis suministrada fue de 36 mg/kg de peso (187). Anteriormente se habia comprobado que la incubacion previa in vitro por 90 min del VIH-1 con anticuerpos policlonales contra gp120 evitaba la infeccion de chimpances (188). En este mismo ensayo el AcM 0.5beta no protegio completamente, pero retardo significativamente la multiplicacion del virus en el animal inoculado. Como este anticuerpo es murino existe la posibilidad de que los inmunocomplejos formados no fueran eliminados adecuadamente de la circulacion y el virus conservara su potencialidad infectiva.
Los experimentos de proteccion en animales por transferencia pasiva de anticuerpos neutralizantes han constituido un elemento importante para la investigacion en vacunas profilacticas. Constituyen hasta la fecha la unica evidencia inequivoca de que un tipo definido de respuesta inmune logra proteger contra la infeccion del VIH-1, e indica ademas, que manteniendo un nivel determinado de anticuerpos en sangre es posible impedir la colonizacion irreversible del sistema inmune por el VIH-1.
Estudios clinicos en humanos. Estos resultados han alentado por una parte la busqueda de anticuerpos de mas amplia reactividad cruzada, o de una mezcla de AcM contra los principales grupos de VIH-1 para la inmunoterapia pasiva del SIDA. Existen dos situaciones donde este proceder puede ser claramente eficaz: la transmision materno-fetal y la exposicion accidental al virus. Mas dificil es predecir si tendra algun efecto beneficioso sobre el curso de la enfermedad en individuos infectados.
Los resultados de dos ensayos clinicos de Fase I en los que se emplearon el AcM murino P4D10 (189) y un AcM quimerico humano/raton construido a partir del AcM BAT123 (190) (Tabla 1) mostraron que ambos fueron bien tolerados y seguros. Aunque estos ensayos de Fase I no estan disenados para evaluar la eficacia del tratamiento, en uno de estos se encontro una disminucion alentadora de la carga viral en algunos pacientes (190). Estos AcMs contra la cepa IIIB no son idoneos para el tratamiento por lo limitado de su reactividad contra las cepas de VIH mas frecuentes. Este estudio brindo la posibilidad adicional de comprobar que la vida media en sangre de este AcM quimerico (ocho a dieciseis dias) es muy superior a la descrita para un AcM murino (15 h) (191), aunque sigue siendo algo inferior a los 21 dias reportados para la IgG normal (192). Homologias y reactividad cruzada con proteinas humanas
Existen algunos reportes en la literatura de homologias entre la region V3 y proteinas humanas. En algunos casos esta solo existe a nivel de secuencia y en otros se ha evidenciado la presencia de anticuerpos con reactividad cruzada entre las moleculas homologas. A continuacion comentamos algunos de estos resultados.
Existe homologia entre la region central de V3 de la cepa NY5 con colageno (193) (Figura 2).
--------------------------------------------------------------------------- Figura 2. Homologias en las secuencias de proteina del colageno y el V3 de la cepa NY5. --------------------------------------------------------------------------- EKGEAGDPGRPGD Cadena alpha1 del colageno humano tipo VI *KG**G*PGR*** Consenso de colagenos homologos KKGIAIGPGRTLY V3 NY5 --------------------------------------------------------------------------- Los sueros de pacientes VIH+ reconocen tanto el peptido de V3-NY5 como el consenso de colageno. En este trabajo solo se presentan los datos de un suero, aunque se plantea que se estudiaron dieciseis VIH+ y quince VIH-. No obstante al purificar los anticuerpos por inmunoafinidad con el peptido NY5 la reactividad cruzada contra el peptido de colageno en ELISA es muy reducida y poco convincente. Previamente se habia sugerido que la presencia de anticuerpos contra colageno en personas VIH negativas podia aumentar su susceptibilidad a la infeccion (194).
De forma similar existen epitopos compartidos entre el V3 de la cepa IIIB y proteinas de celulas nerviosas (195), asi como con una proteina de 70 kD asociada al "splicing" de ARN (196), mientras que la reactividad con un antigeno nuclear parece ser la causa de la localizacion en nucleo de un AcM contra el aislamiento IIIB (197).
Por ultimo la region que abarca el V3 y los diez aminoacidos precedentes tiene homologia con la region variable pesada del grupo III de inmunoglobulinas humanas (198). Entre el 3 y 6 % de los linfocitos B de individuos VIH- reaccionan con gp120 y en los individuos con SIDA se observa un deficit clonal de celulas B con genes de la familia H3 (199). Estos elementos han servido para elaborar la hipotesis de que el V3 puede interferir con la red de regulacion idiotipica del individuo y actuar como un superantigeno para celulas B que expresan VH3 (200, 201). De esa forma se estaria empobreciendo el repertorio de inmunoglobulinas del hospedero, lo cual facilitaria el escape del virus. Sin embargo, llama poderosamente la atencion que la reactividad contra gp120 sea mediada por anticuerpos de clase IgM y no IgG (198), ya que se conoce que las IgM tienden a ser moleculas polirreactivas de baja afinidad y elevada avidez. Por otra parte, un posible superantigeno ha sido localizado, pero no en V3 sino dentro de la region C2 de la gp120 (202). Inmunidad Celular Contra V3 Epitopos para celulas T citotoxicas
En el mismo ano en que fue descrito como epitopo B, Takahashi H et al. (203) reportaron la region V3 de la cepa IIIB como epitopo inmunodominante para celulas T citotoxicas (CTL) en ratones de haplotipo H2^d. El peptido reportado comprende la secuencia RIQRGPGRAFVTIGK de la cepa IIIB. Este hallazgo fue corroborado por diversos autores (204, 205).
Este peptido fue tambien reconocido por CTL de individuos seropositivos (206) y de voluntarios inmunizados con las proteinas de la envoltura (207, 208). En estos ultimos se ha descrito reconocimiento restringido por los alelos HLA-A2, HLA-A3 y HLA-A11.
Takahashi H et al. (209, 210) reportaron por primera vez la existencia de celulas CD4+ con actividad citotoxica en voluntarios inmunizados. Tambien se detecto actividad citotoxica tanto en linfocitos CD4+ como CD8+ en tres de ocho individuos inmunizados con conjugado V3MN-PPD (9) y se vinculo a la presencia del alelo HLA-B7. Una asociacion de este tipo habia sido reportada previamente en casos de seroconversion aguda (211).
La variacion de la region V3 puede tambien generar variantes de escape para celulas CTL (212, 213), aunque tambien se ha descrito la posibilidad de activar clones de CTL polirreactivos contra varias cepas virales (214). Epitopos para celulas T auxiliadoras Diferentes estudios han demostrado que peptidos de V3 pueden ser presentados tambien por los antigenos del Sistema Principal de Histocompatibilidad de Clase II para activar clones de celulas T auxiliadoras (Th) tanto en ratones (204, 206, 215, 216), como en personas infectadas (217), y voluntarios sanos vacunados con gp120 (218). No obstante, no se detecto actividad linfoproliferativa frente al peptido de 24 aminoacidos de V3-IIIB (N N T R K S I R I Q R G P G R A F V T I G K I G) en cinco voluntarios vacunados con gp120 IIIB producida en baculovirus y adyuvada con alumina (219). El V3 como candidato vacunal Estrategias vacunales El principal elemento en contra de una vacuna basada exclusivamente en el PND es el elevado polimorfismo de esta region que genera principalmente anticuerpos tipo especificos (34, 37, 92). Antisueros generados contra un aislamiento viral muestran actividad neutralizante contra la cepa homologa, pero generalmente no son capaces de neutralizar cepas heterologas que divergen en la secuencia de V3 (83).
A pesar de esto, y debido a sus excepcionales propiedades como inmunogeno, diferentes laboratorios han centrado su estrategia vacunal en la region V3. Se han ensayado diferentes formas de presentacion con el proposito de incrementar la inmunogenicidad y lograr titulos altos de anticuerpos neutralizantes.
A continuacion resumimos algunas de las estrategias usadas: Peptidos ciclicos Se han ensayado diferentes variantes de ciclizacion de peptidos de V3. Chimpances inmunizados con mezclas de 21 peptidos ciclicos de la region V3 completa de diferentes cepas desarrollaron anticuerpos contra todos los peptidos, con titulos superiores a 1/25 000 en seis de siete animales (220). Se observo que la respuesta contra gp120 nativa y los titulos neutralizantes fueron superiores en los animales que habian sido previamente inoculados con gp120. En este caso la estructura ciclica se logro en condiciones de oxidacion por formacion de puente disulfuro entre las dos cisteinas. Tambien se han ensayado otras vias de ciclizar el peptido buscando una estructura mas cercana a la de la molecula nativa sin lograr mejoras apreciables (221). Peptidos hibridos T-B Diferentes experimentos han mostrado la factibilidad de generar anticuerpos neutralizantes contra mas de un aislamiento viral a traves de la combinacion de dos o mas peptidos, o el empleo de peptidos hibridos (222, 223). Un considerable esfuerzo se ha dirigido a desarrollar un inmunogeno peptidico definido, que no necesite de proteinas transportadoras. Varios reportes han demostrado que peptidos hibridos, con un contenido de alrededor de quince aminoacidos de la region central del V3 y un epitopo para celulas T auxiliadoras, generan una respuesta de anticuerpos neutralizantes en varias especies (224, 225). La disposicion relativa de los epitopos T y B en estas moleculas parece ser importante en su inmunogenicidad, aunque los resultados en este sentido han sido contradictorios. Algunos autores sugieren que, mas que la polaridad del peptido, la conservacion de los sitios internos de procesamiento proteolitico es el factor que determina su inmunogenicidad. Peptidos multiantigenicos La misma idea ha sido extrapolada al sistema conocido como MAPS, del ingles "Multi Antigenic Peptide System", el cual consiste en un nucleo de lisinas del cual se derivan cuatro o mas ramas compuestas por los peptidos de interes. MAPS divalentes con peptidos de V3 y peptidos T fueron mas eficientes en generar respuesta neutralizante en ratones y cobayos que MAPS monovalentes (226). La inmunogenicidad en conejos fue elevada para ambas moleculas. No obstante, estudios comparativos con peptidos lineales o ciclicos, no han encontrado ventajas significativas para este sistema (227). Peptidos conjugados a proteinas "carriers" Varias proteinas han sido empleadas como carrier de peptidos de V3. Para esto se han empleado metodos quimicos de conjugacion como glutaraldehido (9) o carbodiimida (228).
Cryz SJ et al. (228) inmunizaron con trece aminoacidos de V3 de la cepa MN acoplados a la toxina A de Pseudomona aureoginosa. Este inmunogeno genero titulos de anticuerpos (1:1 000 en ELISA) en cobayos inmunizados con cuatro dosis sin adyuvante. Los sueros de estos animales neutralizaron al MN (1:750) pero solo con bajos titulos las cepas IIIB y RF. Por otra parte, se describio una fuerte reactividad cruzada en ELISA contra un peptido de la cepa SC y el peptido repetitivo (GPGRAF)3, mientras que esta fue muy debil con RF, NY5 y CDC-42.
Por ultimo, otro enfoque interesante consiste en la presentacion de peptidos del V3 acoplados a alfa 2 microglobulina para lograr un transporte dirigido hasta el receptor de esta molecula en monocitos (229). Se demostro que el uso de este sistema es superior a la adyuvacion de los peptidos directamente con adyuvante de Freund. Lipopeptidos
El empleo de lipopeptidos tiene, en teoria, la ventaja de facilitar la presentacion de epitopos T por las moleculas del sistema mayor de histocompatibilidad y potenciar de esta forma la activacion de celulas T citotoxicas. La racionalidad de su empleo estriba en la facilidad del componente lipidico para mediar la introduccion del peptido en el citoplasma celular y propiciar su interaccion con las moleculas de clase I. Se ha descrito la induccion de anticuerpos neutralizantes y respuesta citotoxica en cobayos utilizando MAPS de V3 acoplados a tripalmitoil-S gliceril cisteina sin el empleo de adyuvantes (230). Las ventajas de este sistema en comparacion con otras vias de inducir respuesta humoral y celular estan aun por demostrar. Polipeptidos multiepitopicos
Otra forma de presentacion de la region V3 al sistema inmune son los polipeptidos multiepitopicos (PMEs). Estos son proteinas quimericas que contienen varias regiones V3 separadas entre si por secuencias espaciadores. A traves de la sintesis quimica del gen y su clonacion en un plasmidio de expresion fusionado a un fragmento de la IL2 humana se obtuvo un primer PME que contiene la region central de V3 de las cepas MN, SC y WMJII (60). Esta proteina fue capaz de inducir anticuerpos monoclonales neutralizantes para la cepa MN.
En estudios mas recientes se han generado variantes de PMEs con seis regiones V3 y se ha comprobado la capacidad de inducir altos titulos de anticuerpos contra todos los peptidos incluidos y con actividad neutralizante en conejos contra las cepas MN y LAI (231). A pesar de que el diseno de estos PMEs no esta basado en consideraciones estructurales, los epitopos incluidos quedan bien expuestos y son reconocidos por AcMs especificos.
Esta estrategia tiene las siguientes ventajas y desventajas en relacion con el uso de peptidos sinteticos: Ventajas
- Posibilita la clonacion del gen en un vector vivo. - Permite la inmunizacion con ADN desnudo.
Insercion de V3 en proteinas recombinantes Wagner et al. (232) insertaron la region V3 en diferentes posiciones del gen gag y lo clonaron en el virus Vaccinia. La proteina recombinante expresada tanto en celulas CV1 como en celulas de insectos, mantuvo la capacidad de autoensamblarse y formar particulas parecidas a virus ("virus-like-particles", VLP). Estas VLP expusieron la region V3 hacia la superficie, donde fue reconocida por el 90 % de los sueros humanos VIH-1 positivos. Luo LZ et al. (233) insertaron la region V3 en un fragmento de 41 kD de la proteina precursora de la capsida. Sueros de conejos inmunizados con este producto reconocieron la gp120 y mostraron actividad neutralizante. Una estrategia analoga se siguio con la insercion de V3 en el retrotranposon Ty de levaduras, capaz de formar VLP (215). Anticuerpos anti-idiotipicos
Se han generado AcMs que reconocen el idiotipo de AcM anti V3 (anti-idiotipicos, AI) y se ha logrado demostrar que mimetizan la region V3 y que son capaces de generar anticuerpos neutralizantes en conejos (234) y ratones (235). Esta estrategia en general tiene la desventaja de la baja inmunogenicidad de estas moleculas. En el caso especifico de los modelos de V3 empleados, estos son AcMs tipo especificos, y por consiguiente, solo mimetizan un numero limitado de variantes de V3, y generan una respuesta de espectro estrecho. Uno de los AcMs contra las regiones conservadas (GPGR) podria generar un AI mas universal, pero parece mas atractivo emplear AcMs contra otros epitopos conformacionales mas conservados incluso entre diferentes subtipos (236). Otras variantes Una estrategia original ha sido la sintesis de peptidos repetitivos en los cuales secuencias de V3 se insertan en regiones de la mucina humana MUC1 (237). Esta proteina se utilizo para aportar un marco adecuado para la exposicion de la region V3. Estudios previos indicaban que MUC1 presenta un motivo estructural que garantiza la correcta exposicion de las regiones inmunodominantes. Empleando tecnicas de resonancia magnetica nuclear (NMR), se demostro que los fragmentos de V3 insertados estaban correctamente expuestos, y consecuentemente fueron reconocidos por sueros humanos y anticuerpos monoclonales especificos. Respuesta de voluntarios sanos vacunados con peptidos de V3 Algunas variantes con peptidos de V3 se encuentran ya en ensayos clinicos de Fase I en humanos. United Biomedical Inc. (UBI) realizo un primer ensayo clinico de Fase I donde se evaluaron tres dosis de un MAP con la cepa MN adyuvado con Al(OH)3. La inmunogenicidad y los titulos neutralizantes fueron mayores en la dosis mas alta de 500 g. En todos los voluntarios se detectaron titulos contra el peptido por ELISA despues de tres inmunizaciones y actividad neutralizante en ocho de los diez vacunados (238).
Esta compania tiene otro protocolo clinico en curso con una mezcla de quince MAPS con peptidos de cinco subtipos diferentes. Esta por demostrar si es posible lograr titulos neutralizantes contra este inmunogeno en el que cada uno de los V3 esta mucho menos representado.
En otro ensayo clinico se utilizo un peptido de V3 conjugado a Derivados Proteicos Purificados (PPD) de BCG por el Instituto Suizo de Sueros y Vacunas. Con cinco dosis de solo 640 ug de peptido de quince aminoacidos de MN conjugado a PPD sin adyuvante, se logro generar titulos neutralizantes entre 20 y 200 en ocho voluntarios sanos (9). Tambien se encontro reactividad cruzada en ELISA con SC pero no con IIB ni RF. Se especula que la existencia de una respuesta de memoria previa contra PPD es responsable de la amplificacion de la respuesta contra el peptido.
Una combinacion de peptidos de V3 de las cepas LAI, MN y RF conjugados a hemocianina de "keyhole limpet" (KLH) fue empleado por investigadores de la Universidad de Yokohama (12).
Por ultimo un peptido de V3 (MN) se empleo en un ensayo clinico por Pasteur Merieux despues de dos dosis con gp160 recombinante (239). Este esquema de inmunizacion indujo altos titulos de anticuerpos neutralizantes pero restringidos a la cepa de VIH homologa.
En general, estos inmunogenos, al igual que otros evaluados en Fase I, han demostrado ser bien tolerados y seguros en humanos. En cuanto a la respuesta inmune observada, hasta el momento se ha comprobado la capacidad de despertar anticuerpos neutralizantes de la cepa homologa en la mayoria de los vacunados, si bien estos han sido discretos. Estan aun por evaluar las estrategias mas ambiciosas que se proponen generar una respuesta amplia contra diversos tipos de VIH-1. Aun cuando la informacion disponible no permite concluir que un preparado vacunal basado solamente en la region V3 sea suficiente para brindar proteccion, se espera que al menos su combinacion con otros inmunogenos sea muy positiva. Dada la situacion critica de los modelos animales en SIDA, solo la conduccion de pruebas de eficacia en humanos daran una respuesta satisfactoria a esta interrogante.
Referencias
1.Barre-Sinoussi F, Chermann JC, Rey F, Nuguyere MT, Chamaret S, Gruest J et al. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science 1983; 220: 868-871. 2.Popovic M, Sarngandharan MG, Read E, Gallo RC. Detection isolation and continuous production of cytopathic retrovirus (HTLV-III) from patients with AIDS and pre AIDS. Science 1984; 224: 497-500. 3.UNAIDS. The Global Status and Trends of the Global HIV/AIDS Pandemic, Final Report, July 1996. 4.Derosiers RC. HIV with multiply gene deletions as a life atenuated vaccine for AIDS. AIDS Res Hum Retroviruses 1992; 8: 411-421. 5.Keefer MC, Graham BS, Belshie R, Schwartz D, Corey L, Bolognesi DP et al. Studies of high doses of a human immunodeficiency virus type 1 recombinant glycoprotein 160 candidate vaccine in HIV type 1 seronegaive individuals. AIDS Res Hum Retroviruses 1994; 10(12): 1713-1723. 6.Kieny MP, Rautmann G, Schmitt D, Dott K, Wain-Hobson S, Alizon M et al. AIDS virus env protein expressed from a recombinant vaccinia virus. Biotechnology 1986; 4: 790-795. 7.Stover CK, de la Cruz V, Fuerst TR et al. New use of BCG for recombinants vaccines. Nature 1991; 351: 456-460. 8.Pialoux G, Excler JL, Riviere Y, Gonzalez-Canali G, Feuillie V, Coulaud P et al. A prime boost approach to HIV preventive vaccine using a recombinant canary pox virus expressing gp160 (MN) followed by a recombinant glycoprotein 160 (MN/LAI). AIDS Res Hum Retroviruses 1995; 11: 373-381. 9.Rubinstein A, Goldstein H, Pettoello-Mantovani M, Mizrachi Y, Bloom B, Furer E et al. Safety and immunogenicity of a V3 loop synthetic peptide conjugated to purified protein derivative in HIV-seronegative volunteers. AIDS 1995; 9: 243-251. 10.Wang B, Ugen KE, Srikantan V, Agadjanyan M, Dang K, Refali Y et al. Gene inoculation generates immune responses against human immunodeficiency virus type 1. Proc Natl Acad Sci USA 1993; 90: 4156-4160. 11.Coney L, Wang B, Ugen KE, Boyer J, McCallus D, Srikantan V et al. Facilitated DNA inoculation induces anti HIV-1 immunity in vivo. Vaccine 1994; 12(16): 1545-1550. 12.Fast PE, Walker MC. Human trials of experimental AIDS vaccines. AIDS 1993; 7, (suppl.) 1: 147. 13.Esparza J, Heyward WL, Osmanov S. HIV vaccine development: from basic research to human trials. AIDS 1996; 10 (suppl. A): S123-S132. 14.Hoff R, McNamara J, Fowler MG, McCauley M. HIV vaccine development and clinical trials. Acta Paediatr 1994; (suppl.) 400: 73-3. 15.Papsidero LD, Sheu M, Ruscetti FW. human immunodeficiency virus type 1-neutralizing monoclonal antibodies which react with p17 core protein: characterization and epitope mapping. J Virol 1989; 63: 267-272. 16.Chanh TC, Dreesman GR, Kanda P, Linette GP, Sparrow JT, Ho DD et al. Induction of anti-HIV neutralizing antibodies by synthetic peptides. EMBO J 1986; 5: 3065-3071. 17.Broliden PA, von Gegerfelt A, Clapham P, Rosen J, Fenyo EM, Wharen B et al. Identification of human neutralization-inducing regions of HIV-1 envelope glycoprotein. Proc Nat Acad Sci USA 1992; 89: 461-465. 18.Vella C, Ferguson M, Dunn G, Meloen R, Langedijk H, Evans D et al. Characterization and primary structure of a human immunodeficiency virus type 1/HIV-1 neutralization domain as presented by a poliovirus type 1/HIV-1 chimaera. J Gen Virol 1993; 74: 2603-2607. 19.Muster T, Steindl F, Purtscher M, Trkola A, Klima A, Himmler G et al. A conserved neutralizing epitope on gp41 of human immunodeficiency virus type 1. J Virol 1993; 67: 6642-6647. 20.Gnann JW, Nelson JA, Oldstone MBA. Fine mapping of an immunodominant domain in the transmembrane glycoprotein of human immunodeficiency virus. J Infec Dis 1987; 156: 261-267. 21.Ho D, Kaplan JC, Rackausas I, Gurney ME. Second conserved domain is important for HIV infectivity and antibody neutralization. Science 1988; 239: 1021-1023. 22.Vahlne A, Horal P, Eriksson K, Jeansson S, Rymo L, Hedstrom KG et al. Immunizations of monkeys with synthetic peptides disclose conserved areas on gp120 of human immunodeficiency virus type 1 associated with cross-neutralizing antibodies and T cell recognition. Proc Natl Acad Sci USA 1991; 88: 10744-10748. 23.Ronco J, Charbit A, Dedieu JF, Mancini M, Michel ML, Henin I et al. Is there a neutralization epitope in the second conserved domain of HIV-1 envelope protein? AIDS Res Hum Retroviruses 1991; 7(1): 1-3. 24.Neurath R, Strick N. Confronting the hypervariability of an immunodominant epitope eliciting virus neutralizing antibodies from the envelope glycoprotein of the human immunodeficiency virus type 1 (HIV-1). Mol Immunol 1990; 27: 539-549. 25.Fung MSC, Sun CRY, Gordon WL, Liou RS, Chang TW, Sun WNC et al. Identification and characrterization of a neutralization site within the second variable region of human immunodeficiency virus type 1 gp120. J Virol 1992; 66: 848-856. 26.Sun N, Ho D, Sun C, Liou R, Gordon W, Fung M et al. Generation and characterization of monoclonal antibodies to the putative CD4 binding domain of human immunodeficiency virus type 1 gp120. J Virol 1989; 63: 3579-3585. 27.Neurath R, Strick N, Lee ES. B-cell epitope mapping of human immunodeficiency virus envelope glycoproteins with long (19-36 residues) synthetic peptides. J Gen Virol 1990; 71: 85-95. 28.Ho D, Fung MSC, Yoshiyama H, Cao Y, Robinson J. Discontinuous epitopes on gp120 Important in HIV-1 neutralization. AIDS Res Hum Retroviruses 1992; 8(8): 1337-1339. 29.Roben P, Moore J, Thali M, Sodroski J, Barbas C, Burton D. Recognition of a panel of human recombinant fab fragments to the CD4 binding site of gp120 that show differing abilities to neutralize HIV 1. J Virol 1994; 68(8): 4821-4828. 30.Steimer K, Scandella CJ, Skites PV, Haigwood N. Neutralization of divergent HIV-1 isolates by conformation-dependent human antibodies to gp120. Science 1991; 253: 1-4. 31.Moore JP, Ho D. Antibodies to discontinuous or conformationally sensitive epitopes on the gp120 glycoprotein of human immunodeficiency virus type 1 are highly prevalent in sera of infected humans. J Virol 1993; 67(2): 863-875. 32.Modrow S, Hahn B, Shaw GM, Gallo RC, Staal F, Wolf H. Computer-assisted analysis of envelope protein of seven human immunodeficiency virus isolates. Prediction of antigenic epitopes in conserved and variable regions. J Virol 1987; 61: 570-578. 33.Palker TJ, Clark M, Langloise AJ, Mathews TJ, Weinhold KJ, Randall RR et al. Type specific neutralization of the human immunodeficiency virus with antibodies to env-encoded synthetic peptides. Proc Natl Acad Sci USA 1988; 85: 1932-1936. 34.Rusche JR, Javaherian K, McDanal C, Petro J, Lynn DL, Grimaila R et al. Antibodies that inhibit fusion of human immunodeficiency virus-infected cells bind a 24-amino acid sequence of viral envelope gp120. Proc Natl Acad Sci USA 1988; 85: 3192-3202. 35.Goudsmit J, Debouck C, Meloen RH, Smith L, Bakker M, Asher DM et al. Human immunodeficiency virus type 1 neutralization epitope with conserved architecture elicits early type-specific antibodies in experimentally infected chimpanzees. Proc Natl Acad Sci USA 1988(a); 85: 4478-4482. 36.Putney S, Javaherian K, Profy A, Emini E, Langlois A, Ross K et al. In: Hum Retroviruses 1990; 323-335, Edited by Alan R. Liss. 37.Javaherian K, Langlois AJ, McDanal C, Ross KL, Eckler LI, Jellis CL et al. Principal neutralizing domain of the HIV-1 envelope protein. Proc Natl Acad Sci USA 1989; 86: 6768-6772. 38.Putney S, Javaherian K, Rusche J, MatthewsT, Bolognesi D. Features of the HIV envelope and development of a subunit vaccines in AIDS Vaccine Research and Clinical trials, Eds. Putney S and Bolognesi D. Marcel Dekkler, New York, 1990; 3-61. 39.Goudsmit J, Boucher AB, Meloen RH, Epstein LG, Smit L, Hoek et al. Immunodominant B-cell epitopes of HIV-1 envelope recognized by infected and immunized host. AIDS 1988(b); 2: 157-164. 40.Laman JD, Schellekens MM, Lewis GK, Moore JP, Matthews TJ, Langedijk JP et al. A hidden region in the third variable domain of HIV-1 IIIB gp120 identified by a monoclonal antibody. AIDS Res Hum Retroviruses 1993; 9(7): 605-612. 41.La Rosa GJ, Davide JP, Weinhold K, Waterbury J, Profy AT, Lewis JA et al. Conserved sequence and structural elements in the HIV-1 principal neutralizing determinant. Science 1990; 249: 932-935. 42.Chandrasekhar K, Profy AT, Dyson HJ. Solution conformational preferences of immunogenic peptides derived from the principal neutralizing determinant of the HIV-1 envelope glycoprotein gp120. Biochemistry 1991; 30: 9187-9194. 43.Vranken WF, Budesinsky M, Fant F, Boulez K, Borremans FAM. The complete consensus V3 loop peptide of the envelope protein gp120 of HIV-1 shows pronounced helical character in solution. FEBS Letters 1995; 374: 117-121. 44.Wilson I, Stanfield R, Ghiara J, Stura E, Satterthwait A, Profy A. Structural basis for recognition of HIV-1 by two broadly neutralizing monoclonal antibodies. IVth Int Conf AIDS, Berlin, 1993, WS-A21-1, Abstract Book 1: 37. 45.Ghiara J, Stura E, Stanfield R, Profy A, Wilson I. Crystal structure of the Principal Neutralization Site of HIV-1. Science 1994; 264: 82-85. 46.Matsushita S, Robert-Guroff M, Rusche J, Koito A, Hattori T, Javaherian K et al. Characterization of a human immunodeficiency virus Neutralizing Monoclonal Antibody and Mapping of the Neutralizing Epitope. J Virol 1988; 62(6): 2107-2114. 47.Skinner MA, Ting R, Langlois AL, Weinhold KJ, Lyerly HK, Javaherian K et al. Characteristics of a neutralizing monoclonal antibody to the HIV envelope glycoprotein. AIDS Research and Human Retroviruses 1988; 4(3): 87-197. 48.Liou RS, Rosen EM, Fung MSC, Sun WNC, Sun C, Gordon W et al. A chimeric mouse-human antibody that retains specificity for HIV gp120 and mediates the lysis of HIV-infected cells. J Immunol 1989; 143: 3967-3975. 49.Broliden PA, Ljunggren K, Hinkula J, Norrby E, Akerblom L Wahren B. A monoclonal antibody to human immunodeficiency virus type 1 which mediates cellular cytotoxicity and neutralization. J Virol 1990; 64: 2. 50.Scott CF, Silver S, Profy AT, Putney SD, Langlois A, Weinhold K et al. Human monoclonal antibody that recognized the V3 region of human immunodeficiency virus gp120 and neutralizes the human T-lymphotropic virus typeIII MN strain. Proc Natl Acad Sci USA 1990; 8: 8596-8601. 51.Durda PJ, Bacheler L, Clapham P, Jenoski AM, Leece B, Matthews TJ et al. HIV-1 neutralizing monoclonal antibodies induced by a synthetic peptide. AIDS Res Hum Retroviruses 1990; 6(9): 1115-1213. 52.Moore JP, McKutchan FE, Poon SW, Mascola J, Liu J, Cao Y et al. Exploration of antigenic variation in gp120 from clades A through F of human immunodeficiency virus type 1 by using monoclonal antibodies. J Virol 1994; 68(12): 8350-8364. 53.Robinson JE, Yoshijama H, Holton D, Elliot S, Ho D. Distinct antigenic sites on HIV gp120 identified by a panel of human monoclonal antibodies. J Cell Biochem 1992; (suppl.)16E: 71. 54.Ohno T, Terada M, Yoneda Y, Shea KW, Chambers RF, Stroka DM et al. A broadly neutralizing monoclonal antibody that recognizes the V3 region of HIV-1 glycoprotein gp120. Proc Natl Acad Sci USA 1991; 88: 10726-10729. 55.Gorny MK, Xu JY, Gianakos V, Karwowska S, Williams C, Sheppard HW et al. Production of site selected neutralizing antibodies against the third variable domain of the human immunodeficiency virus type 1 envelope plycoprotein. Proc Natl Acad Sci USA 1991; 88: 3238-3242. 56.Buchbinder A, Karwowska S, Gorny MK, Burda ST, Zolla-Pazner S. Synergy between human monoclonal antibodies to HIV extends their effective biologic activity against homologous and divergent strains. AIDS Res Hum Retroviruses 1992; 8(4): 425-427. 57.Barbas C, Bjorling E, Chiodi F, Dunlop N, Cabada D, Jones TM et al. Recombinant human Fab fragments neutralize human type 1 immunodeficiency virus in vitro. Proc Natl Acad Sci USA 1992; 89: 9339-9343. 58.Pal R, Di Marzo Veronese F, Nair BC, Rahman R, Hoke R et al. Characterization of a neutralizing monoclonal antibody to the external glycoprotein of HIV-1. Intervirology 1992; 86: 86-93. 59.White-Scharf ME, Potts B, Smith LM, Sokolowski KA, Rusche JR, Silver S. Broadly neutralizing monoclonal antibodies to the V3 region of HIV-1 can be elicited by peptide immunization. Virology 1993; 192: 197-205. 60.Duarte CA, Montero M, Seralena A, Valdes R, Jimenez V, Benitez J et al. Multiepitope polypeptide of the HIV-1 envelope induces monoclonal antibodies against V3 loop. AIDS Res Hum Retroviruses 1994; 10(3): 235-243. 61.Forthal DN, Landucci G, Gorny M, Zolla-Pazner S, Robinson WE. Functional activities of 20 human immunodeficiency virus type 1 (HIV-1) specific human monoclonal antibodies. AIDS Res Hum Retroviruses 1995; 11(9): 1095-1099. 62.Nara P, Smet L, Dunlop N, Hatch W, Merges M, Waters D et al. Emergence of resistant to neutralization by V3-specific antibodies in experimental HIV-1 III-B infection of chimpanzees. J Virol 1990; 64: 3779. 63.Willey R, Ross E, Buckler-White A, Theodore T, Martin M. Functional interaction of constant and variable domains of HIV-1 gp120. J Virol 1989; 63: 3595-3600. 64.Neurath AR, Strick N, Jiang S. Synthetic peptides and anti-peptide antibodies as probes to study interdomain interactions involved in virus assembly: The envelope of the HIV-1. Virology 1992; 188: 1-13. 65.Robert-Guroff M, Louie A, Myagkikh, Michaels F, Kieny MP, White-Scharf ME, Potts B, Groog D, Ritz MS. Alteration of the V3 loop context within the envelope of Human Immunodeficiency Virus type 1 enhances neutralization. J Virol 1994; 68(6): 3459-3466. 66.Luciw P, Shacklett BL. Molecular biology of the human immunodeficiency viruses. In HIV Molecular Organization, Pathogenicity and Treatment, 1993 Eds Morrow WJW and Haigwood NL. Elsevier Science Publishers. 67.Ho D, Neuman AU, Perelson AS, Chen W, Leonard JM, Markowitz. Rapid turn-over of plasma virions and CD4 lymphocytes in HIV-1 infection. Nature 1995; 373: 123-126. 68.Wei X, Ghosh SK, Johnson VA, Emini E, Deutsch P, Lifson JD et al. Viral dynamics in human immunodeficiency virus type 1 infection. Nature 1995; 373: 117-122. 69.Ratner L, Haseltine W, Patarca R, Livak KJ, Starcich B, Josephs S et al. Complete nucleotide sequence of the AIDS virus, HTLV-III. Nature 1985; 313: 277. 70.Myers G, Korber B, Wain-Hobson S, Smith R, Pavlakis GN (eds). Human Retroviruses and AIDS. A Compilation and Analysis of nucleic acid and amino acid sequences, 1993. Theoretical Biology and Biophysics Group T-10, Mail Stop K710, Los lamos National Laboratory, Los lamos, New Mexico 87545, USA. 71.Myers G, Korber B, Wain-Hobson S, Smith R, Pavlakis GN (eds). Human Retroviruses and AIDS. A Compilation and Analysis of nucleic acid and amino acid sequences. 1994, Theoretical Biology and Biophysics Group T-10, Mail Stop K710, Los lamos National Laboratory, Los lamos, New Mexico 87545, USA. 72.Rolo FM, Miranda L, Wainberg MA, Gu ZX, Lobaina L, Noa E, Mato J et al. Envelope V3 region sequences of Cuban HIV-1 isolates. J Acquired Immune Deficiency Syndromes and Human Retrovirology 1995; 9(2): 123-125. 73.Gomez C, Fernandez JR, Iglesias E, Lopez AE, Lobaina L, Noa E et al. Complete DNA sequence of the gene coding for the external glycoprotein (gp120) from a Cuban HIV-1 isolate. AIDS Res Hum Retroviruses 1996; 12(6): 553-555. 74.Balfe P, Simmonds O, Ludlam CA, Bishop JO, Brown AJL. Concurrent evolution of human immunodeficiency virus type 1 sequence heterogeneicity to stage of disease. J Virol 1990; 64: 6221-6233. 75.Nowak MA, Anderson RM, McLean AR, Wolfs TFW, Goudsmit J, May RM. Antigenic diversity thresholds and the development of AIDS. Science 1991; 254: 963-969. 76.McNearny Y, Hornickova Z, Markham R, Birdwell A, Arens M, Saah A et al. Relationship of human immunodeficiency virus type 1 sequence heterogeneicity to stage of disease. Proc Natl Acad Sci USA 1992; 89: 10247-1251. 77.De Wolf TFW, de Jong JJ, van der Berg H, Tunagel JMH, Krone WJA, Goudsmit J. Evolution of sequences encoding the principal neutralization epitope of human immunodeficiency virus 1 in host dependent, rapid and continuous. Proc Natl Acad Sci 1990; 87: 9938-9942. 78.Michael N, Davis KE, Loomis-Price LD, VanCott TC, Burke DS, Redfield RR et al. V3 seroreactivity and sequence variation: tracking the emergence of V3 genotypic variation in HIV-1 infected patients. AIDS 1996; 10: 121-129. 79.Watkins B, Ritz M, Wilson C, Aldrich K, Davis A, Robert-Guroff M. Immune escape by human immunodeficiency virus type 1 from neutralizing antibodies: Evidence for multiple pathways. J Virol 1993; 67(12): 7493-7500. 80.Nara PL, Smit L, Dunlop N, Hatch W, Merges M, Waters D et al. Evidence for rapid selection and deletion of HIV-1 subpopulations in vivo by V3 specific neutralizing antibody: a model of humoral-associated selection. Dev Biol Stand 1990; 72: 315-341. 81.Kuiken CL, Lukasov V, Baan E, Dekker J, Leunissen JAM, Goudsmit J. Evidence for limited within person evolution of the V3 domain of the HIV-1 envelope in the Amsterdam population. AIDS 1995; 10: 31-37. 82.Carrow EN, Vujcic LK, Glass WL, Seamon KB, Rastog SC, Hendry RM et al. High prevalence of antibodies to the gp120 V3 region principal neutralizing determinant of HIV-1 MN in sera from Africa and the Americas. AIDS Res Hum Retroviruses 1991; 7(10): 831-838. 83.Baillou A, Brand D, Denis F, Boup SM, Chout R, Goudeau A et al. High antigenic cross-reactivity of the V3 consensus sequences of HIV-1 gp120. AIDS Res Hum Retroviruses 1993; 9(12): 1209-1215. 84.Cheingsong-Popov R, Lister S, Callow D, Kaleebu P, Beddows S, Weber J et al. Serotyping HIV type 1 by antibody binding to the V3 loop. Relationship to viral genotype. AIDS Res Hum Retroviruses 1994; 10(11): 1379-1386. 85.Lopez-Campistrous AE, Reyes O, Garay H, Diaz H, Duarte CA. Antibodies against HIV-1 V3 loop synthetic peptides in the plasma of Cuban seropositives recognize more frequently African sequences and are inversely related to disease progression. Sent to AIDS for publication. 86.Sherefa K, Sallberg M, Sonnerborg A. Evidence of no change in V3 loop antibody recognition pattern in HIV type 1-infected Ethiopians between 1988 and 1933. AIDS Res Hum Retroviruses 1994; 10(11): 1551-1556. 87.Holmback K, Kusk P, Hulgaard EF, Bugge TH, Scheibel E et al. Autologous antibody response against the principal neutralizing domain of human immunodeficiency virus type 1 isolated from infected humans. J Virol 1993; 67(3): 1612-1619. 88.Pau CP, Lee-Thomas S, Auwanit W, George JR, Ou CY, Parekh BS et al. Highly specific V3 peptide enzyme immunoassay for serotyping HIV-1 specimens from Tailand. AIDS 1993; 7: 337-340. 89.Wasi C, Herring B, Raktham S, Vanichsen, Mastro TD, Young NL et al. Determination of HIV subtypes in injecting drug users in Bangkok, Thailand using peptide enzyme immunoassay and the heteroduplex mobility assay: Evidence of increasing infection with HIV-1 subtype E. AIDS 1995; 9(8): 843-851. 90.Cheingsong-Popov R, Lister S, Callow D, Kaleebu P, Beddows S, Weber J et al. Serotyping HIV type 1 by antibody binding to the V3 loop. Relationship to viral genotype. AIDS Res Hum Retroviruses 1994; 10(11): 1379-1386. 91.Pau CP, Kai M, Holloman-Candal DL, Luo CC, Kalish M, Schochetman G et al. Antigenic variation and serotyping of HIV type 1 from four World Health Organization-sponsored HIV vaccine sites. AIDS Res Hum Retroviruses 1994; 10(11): 1369-1377. 92.Zwart G, Langedijk H, van der Hoek L, de Jong JJ, Wolfs TF, Ramautarsing C et al. Immunodominance and antigenic variation of the principal neutralization domain of HIV-1. Virol 1991; 181: 481-489. 93.Nara P, Goudsmit J. Clonal dominance of the neutralizing response to the HIV-1 V3 epitope evidence for "Original Antigenic Sin" during vaccination and infection in animals, including humans. Vaccines 1991; 91. Cold Spring Harbor Lab press. 0-87969-367-391. 94.Fenouillet E, Blanes N, Benjuad A, Gluckman JC. Anti-V3 antibody reactivity correlates with clinical stage of HIV-1 infection and with serum neutralizing activity. Clin Exp Immunol 1995; 99: 419-424. 95.Rossi P, Moschese V, Broliden PA, Fundaro C, Quinti I, Plebani A et al. Presence of maternal antibodies to human immunodeficiency virus type 1 envelope protein gp120 epitopes correlates with the uninfected status of children born to seropositive mothers. Proc Natl Acad Sci USA 1989; 86: 8055-8058. 96.Devash Y, Clavelli A, Wood K, Reagan KJ, Rubinstein A. Vertical transmission of human Immunodeficiency Virus is correlated with the absense of high-affinity/avidity maternal antibodies to the gp120 principal neutralizing determinant. Proc Natl Acad Sci USA 1990; 87: 3445-3449. 97.Parekh BS, Shaffer MN, Pau CP, Abrams E, Thomas EP, Pollack H et al. Lack of correlation between maternal antibodies to V3 peptides of gp120 and perinatal HIV-1 transmission. AIDS 1991; 5: 1179-1184. 98.Halsey NA, Markham R, Wahren B, Boulos R, Rossi P, Wigzell H. Lack of association between maternal antibodies to the V3 loop peptides and maternal-infant HIV-1 transmission. J Acquired Immune Deficiency Syndrome 1992; 5: 153-157. 99.Rubinstein A, Goldstein H, Calvelli T, Devash Y, Rubinstein R, Soeiro R et al. Maternofetal transmission of human immunodeficiency virus-1: The role of antibodies to the V3 primary neutralizing domain. Pediatric Research 1992; 33(1): 76-79. 100.Moore JP. The reactivities of HIV-1+ human sera with solid phase V3 loop peptides can be poor predictors of their reactivities with the V3 loops on native gp120 molecules. AIDS Res Hum Retroviruses 1993; 9(3): 209-218. 101.Tschacher E, Buchow H, Gallo RC, Reitz MS. Funtional contribution of cysteine residues to the human immunodeficiency virus type 1 envelope. J Virol 1990; 64: 2250-2259. 102.Freed EO, Myers DJ, Risser R. Identification of the principal neutralizing determinant of human immunodeficiency virus type 1 as a fusion domain. J Virol 1991; 65: 190-194. 103.Chiou SH, Freed EO, Paganiban AT, Keneally WR. Studies on the role of the V3 region in human immunodeficiency virus type 1 envelope glycoprotein function. AIDS Res Hum Retroviruses 1992; 8(9): 1611-1617. 104.Bergeron L, Sullivan N, Sodrowski J. Target cell specific determinants of membrane fusion within the human immunodeficiency virus type 1 gp120 third variable region and gp41 amino terminus. J Virol 1992; 66(4): 2389-2397. 105.Kowalski M, Potz J, Basiripour L Dorfman T, ChunGoh T, Terwillinger E et al. Funtional regions of the envelope glycoprotein of the human immunodeficiency virus type 1. Science 1987; 237: 1351-1355. 106.Helseth E, Kowalsky M, Gabudza D, Olshevsky U, Haseltine D, Sodrowski J. Rapid complementation assays measuring replicative potential of HIV-1 envelope glycoprotein mutants. J Virol 1990; 64: 2416-2420. 107.Freed EO, Myers DJ, Risser R. Characterization of the fusion domain of the human immunodeficiency virus type 1. Proc Natl Acad Sci USA 1990; 87: 4650-4654. 108.Freed EO, Risser R. Identification of conserved residues in the human immunodeficiency virus type 1 PND that are involved in fusion. AIDS Res Hum Retroviruses 1991; 7(10): 807-811. 109.Ivanoff L, Looney D, McDanal C, Morris J, Wong-Staal F, Langlois A et al. Alteration of HIV-1 infectivity and neutralization by a single amino acid replacement in the V3 loop domain. AIDS Res Hum Retroviruses 1991; 7(7): 595-603. 110.Page KA, Stearns S, Littman D. Analysis of mutations in the V3 domain of gp160 that affect fusion and infectivity. J Virol 1992; 66(1): 524-533. 111.Schulz T, Reeves JD, Head JG, Tailor C, Stephens P, Clements G, et al. Effect of mutations in the V3 loop of HIV-1 gp120 on infectivity and susceptibility to proteolytic cleavage. AIDS Res Hum Retroviruses 1993; 9(2): 159-166. 112.Tersmette M, Lange JMA, de Goede REY, de Wolf F, Eeftinck Schattenkerk JKM, Schellekens PTA et al. Association between biological properties of human immunodeficiency virus type 1 variants and risk for AIDS and AIDS mortality. Lancet 1989; 1: 983-985. 113.Gruters RA, Terpstra FG, de Goede REY, Mulder JW, de Wolf F, Schellekens PTA, van Lier RAW et al. Immunologicla and virological markers in individuals progressing to AIDS. AIDS 1991; 5: 837-844. 114.Shuitemaker H, Koot M, Kootstra NA, Dercksen MW, de Goede RY, Steenwijk RP et al. Biological phenotype of human immunodeficiency virus type 1 clones at different stages of infection: Progression of disease is associated with a shift from monocytotropic to T-cell tropic virus populations. Virol 1992; 66: 1354-1360. 115.Koot M, Keet IPM, Vos AHV, de Goede REY, Roos MTL, Coutinho RA et al. Prognostic value pf HIV-1 syncytium inducing phenotype for rate of CD4+ cell depletion and progression to AIDS. Ann Intern Med 1993; 118: 681-688. 116.de Jong JJ, de Ronde A, Keulen W, Tersmette M, Goudsmit J. Human immunodeficiency virus type 1 clones chimeric for the envelope V3 domain differ in syncytium formation and replication capacity. J Virol 1992; 66(11): 6777-6780. 117.Callahan L. HIV-1 vrion-cell interactions: An electrostatic model of pathogenicity and syncytium formation. AIDS Res Hum Retroviruses 1994; 10(3): 231-233. 118.Callahan LN, Phelan P, Mallinson M, Norcross MA. Dextran sulfate blocks antibody binding to the principal neutralizing domain of human immunodeficiency virus type 1 without interfering with gp120-CD4 interactions, J Virol 1991; 65: 581. 119.Patel M, Yanagishita M, Roderiquez G, Bou-Habib DC, Oravecz T, Hascall VC and Norcross MA: Cell surface heparan sulfate proteoglycan mediates HIV-1 infection of T cell lines. AIDS Res Hum Retroviruses 1993; 9: 167-174. 120.Mbemba E, Czyrski JA, Gattegno L. The interaction of a glycosaminoglycan, Heparin with HIV-1 major envelope glycoprotein. Biochimica and Biophysica Acta 1992; 1180(2): 123-129. 121.Fouchier RAM, Groenink M, Kootstra NA, Tersmette M, Huisman HG, Miedema F et al. Phenotype associated sequence variation in the third variable region of the human immunodeficiency virus type 1. Proc Natl Acad Sci USA 1992; 89: 9434-9438. 122.Cheng-Mayer C, Shioda T, Levy JA. Host range, replicative and cytophatic properties of human immunodeficiency virus type 1 are determined by very few amino acid changes in Tat and gp120. J Virol 1991; 65: 6931-6941. 123.Kuiken Vl, de Jong J, Baan E, Keulen W, Termette M, Goudsmit J. Evolution of the V3 envelope domain in proviral sequences and isolates of human immunodeficiency virus type 1 during transition of the viral biologial phenotype. J Virol 1992; 66: 4622-4627. 124.Grimaila RJ, Fuller BA, Rennert PD, Nelson MB, Hammarskjold ML, Potts B et al. Mutations in the principal neutralization determinant of the human immunodeficiency virus type 1 affect syncytium formation, virus infectivity, growth kinetics and neutralization. J Virol 1992; 66: 1875-1883. 125.Conner RI, Notermans DW, Mohri H, Cao Y, Ho DD. Biological cloning of functionally diverse quasispecies of HIV-1. AIDS Res Hum Retroviruses 1993; 9: 541-546. 126.Oka S, Ida S, Shioda T, Takebe Y, Kobayashi N, Shibuya Y et al. Genetic analysis of HIV-1 during rapid progression to AIDS in an apparently healthy man. AIDS Res Hum Retroviruses 1994; 10(3): 271-277. 127.O'Brien WA, Koyanag Y, Namazie A, Zhao JQ, Diagne A, Idler K et al. HIV-1 tropism for mononuclear phagocytes can be determined by regions of gp120 outside the CD4 binding domain. Nature 1990; 342: 69-73. 128.Cann AJ, Churcher MJ, Boyd M, O'Brien W, Zhao JQ, Zack J et al. The region of the envelope gene of human immunodeficiency virus type 1 responsible for determination of cell tropism. J Virol 1992; 66(1): 305-309. 129.Hwang SS, Boyle TJ, Lyerly K, Cullen BR. Identification of the envelope V3 loop as the primary determinant of cell tropism in HIV-1. Science 1991; 253: 71-74. 130.York-Higgins D, Cheng-Mayer C, Baue D, Levy JA, Dina D. Human immunodeficiency virus type 1 cellular host range, replication and cytopathicity are linked to the envelope region of viral genome. J Virol 1990; 64: 4016-4020. 131.Liu ZQ, Wood C, Levy J, Cheng-Mayer C. The viral envelope gene is involved in macrophage tropism of a human immunodeficiency virus type 1 strain isolated from brain tissue. J Virol 1990; 64: 6148-6153. 132.Shioda T, Levy J, Cheng-Mayer. Macrophage and T cell line tropism can be determined by specific regions of the envelope gp120 gene. Nature 1991; 349: 167-169. 133.Westervelt P, Gendelman H, Ratner L. Identification of a determinant within the human immunodeficiency virus type 1 surface envelope protein critical for productive infection of primary monocytes. Proc Natl Acad Sci USA 1991; 88: 3097-3101. 134.Takeuchi Y, Akutsu M, Murayama K, Shimuzu N, Hoshino H. Host range mutant human immunodeficiency virus type 1: modification of cell tropism by a single point mutation at the neutralization epitope in the env gene. J Virol 1991; 65: 1710-1718. 135.Sharpless NE, O'Brien WA, Verdin E, Kufta CV, Chen ISY, Dubois-Dalcq M. Human immunodeficiency virus type 1 tropism for brain microglial cells is determined by a region of the env glycoprotein that also controls macrophage tropism. J Virol 1992; 66(4): 2588-2593. 136.Hattori TA, Kaito K, Takatsuki H, Katunuma N. Involvement of tryptase-related cellular protease(s) in HIV-1 infections. FEBS Letters 1989; 248: 48-52. 137.Kido H, Fukutomi A, Katunuma N. A novel membrane-bound serine esterase in human CD4 lymphocytes immunologically reactive with antibody inhibiting syncytia induced by HIV. Purification and characterization J Biol Chem 1990; 265: 21979-21985. 138.Clements GL, Price-Jones MJ, Stephens PE, Sutton C, Schultz TF, Clapham PR et al. The V3 loops of the HIV-1 and HIV-2 surface glycoproteins contains proteolytic cleavage sites: A possible function in viral fusion? AIDS Res Hum Retroviruses 1991; 7: 3-16. 139.Murakami T, Hattori T, Takatsuki KA. Principal neutralizing domain of human immunodeficiency virus type 1 interacts with proteinase-like molecule(s) at the surface of Molt4 clone 8 cells. Biochimica et Biophysica Acta 1991; 1079: 279-284. 140.Avril LE, Di Martino-Ferrer M, Barin F, Gauthier F. Interaction between a membrane-associated serine proteinase of U-937 monocytes and peptides from the V3 loop of the human immunodeficiency virus type 1 (HIV-1) gp120 envelope glycoprotein. FEBS 1993; 317(12): 167-172. 141.Callebaut C, Krust B, Jacotot E, Hovanessian A. T cell activation antigen, CD26, as a cofactor for entry of HIV in CD4+ cells. Science 1993; 262: 2045-2050. 142.Autiero M, Abrescia P, Dettin M, Di Bello C, Guardiola J. Binding to CD4 of synthetic peptides patterned on the principal neutralizing domain of HIV-1 envelope protein. Virology 1991; 185: 820-828. 143.Batinic D, Robey FA. The V3 region of the envelope glycoprotein of human immunodeficiency virus type 1 binds sulfated polysaccharides and CD4-derived synthetic peptides. J Biol Chem 1992; 267: 6664-6671. 144.Camerini D, Seed B. A CD4 domain important for HIV mediated syncytium formation lies outside the virus binding site. Cell 1990; 60: 747-754. 145.Truneh A, Buck D, Cassat DR, Juszczak, R, Kassis S, Ryu SE et al. A region in domain 1 of CD4 distinct from the primary gp120 binding site is involved on HIV infection and virus mediated fusion. J Biol Chem 1991; 266: 5942-5948. 146.Langedijk JPM, Puijk WC, Hoorn WPN, Meloen RH. Location of CD4 dimerization site explains critical role of CDR3-like in HIV-1 infection and T cell activation and implies a model for complex of coreceptor-MHC. J Biol Chem 1993; 268: 16875-16878. 147.Hwang SS, Boyle TJ, Lyerly K, Cullen BR. Identification of the envelope V3 loop as the mayor determinant of CD4 neutralization sensitivity. Science 1991; 257: 535-537. 148.De Rossi A, Pasta M, Mammano F, Panozzo M, Dettin M, Di Bello C et al. Synthetic peptides from the principal neutraliizing domain of human immunodeficiency virus type 1 (HIV-1) enhance HIV-1 infection through a CD-dependent mechanism. Virology 1991; 184: 187-196. 149.Nehete RN, Arlinghaus RB, Sastry KJ. Inhibition of human immunodeficiency virus type 1 infection and syncytium formation in human cells by V3 loop synthetic peptides from gp120. J Virol 1993; 67: 6841-6846. 150.Fantini J, Yahi N, Mabrouk K, van Rietschoten J, Rochat H, Sabatier JM. CR Acad Sci Paris 1993; 316: 1381-1387. 151.Yahi N, Fantini J, Mabrouk K, Tamalet K, de Micco P, van Rietschoten J et al. Multibranched V3 peptides inhibit HIV-1 infection in human lymphocytes and macrophages. J Virol 1994; 68(9): 5714-5720. 152.Benjouad A, Fenouillet E, Gluckman JC, Sabatier JM. Multi-branched peptide constructs (MBPC) of the V3 loop of envelope glycoprotein gp120 inhibit human immunodeficiency virus induced syncytium formation. Antiviral Chem Chemother 1994; 5: 195-196. 153.Benjouad A, Chapuis F, Fenouillet E, Gluckman JC. Multibranched peptide constructs derived from the V3 loop of envelope glycoprotein gp120 inhibit Human Immunoideficiency Virus type 1 infection through interaction with CD4. Virology 1995; 206: 457-464. 154.Lu S, Putney S, Robinson H. Human immunodeficiency virus type 1 entry to T cell: More rapid escape from an Anti V3 loop than from an antireceptor antibody. J Virol 1992; 66(4): 2547-2550. 155.Yahi N, Baghdiguian S, Moreau H, Fantini J. Galactosyl ceramide (or a closely related molecule) is the receptor for human immunodeficiency type 1 on human colon epithelial cells. J Virol 1992; 66: 4448-4454. 156.Harouse JM, Bhat S, Spitalnik SL, Lughlin M, Stefano K, Silberberg DH et al. Inhibition of entry of HIV-1 in neural cell lines by antibodies against galactosyl ceramide, Science 1991; 253: 320-323. 157.Bhat S, Spitalnik S, Gonzalez Scarano F, Silbergerg DH: Galactosyl ceramide or a derivative is an essential component of the neural receptor for HIV-1 envelope glycoprotein gp120, Proc Natl Acad Sci USA 1991; 88: 7131-7134. 158.Yahi N, Fantini J, Baghdiguian S, Mabrouk K, Tamalet C, Rochat H et al. SPC3, a synthetic peptide derived from the V3 domain of human immunodeficiency virus type 1 (HIV-1) gp120, inhibits HIV-1 entry into CD4+ and CD4- cells by two distinct mechanisms. Proc Natl Acad Sci USA 1995; 92: 4867-4871. 159.Moore JP, Trkola A, Korber B, Boots LJ, Kessler JA, McCutchan FE et al. A human monoclonal antibody to a complex epitope in the V3 region of gp120 of a human immunodeficiency virus type 1 has broad reactivity within and outside Clade B. J Virol 1995; 69(1): 122-130. 160.VanCott TC, Bethke FR, Polonis VR, Gorny M, Zolla-Pazner S, Redfield RR et al. Dissociation rate of antibody gp120 binding interactions is predictive of V3 mediated neutralization of HIV-1. J Immunol 1994; 153(1): 449-459. 161.Spear GT, Takefman DM, Sullivan EK, Landay AL, Zolla-Pazner S. Complement activation by human monoclonal antibodies to human immunodeficiency virus. J Virol 1993; 67(1): 53-59. 162.Nakamura M, Sasaki H, Terada M, Ohno T. Complement-dependent virolysis of HIV-1 with monoclonal antibody NM-01. AIDS Res Hum Retroviruses 1993; 9(7): 619-626. 163.Tilley SA, Honnen WJ, Racho ME, Chou TC, Pinter A. Synergistic neutralization of HIV-1 by human monoclonal antibodies against the V3 loop and the CD4 binding site of gp120. AIDS Res Hum Retroviruses 1992; 8(4): 461-467. 164.Montefiori DC, Stewart K, Aheam JM, Zhou J. Complement mediated binding of naturally glycosilated and glycosilation-modified HIV-1 to human CR2 (CD21). J Virol 1993; 67: 2699-2706. 165.Matsushita S, Maeda H, Kimachi K. Characterization of a mouse/human chimeric monoclonal antibody (C beta 1) to a principal neutralizing domain of the human immunodeficiency virus type 1 envelope protein. AIDS Res Hum Retroviruses 1992; 8(6): 1107-1115. 166.Duarte CA., Perez L, Vazquez J, Duenas M, Vilarubia OL, Navea L et al. Epitope mapping, V region DNA sequence, and neutralizing Fab fragments of two monoclonal antibodies against the HIV-1 V3 loop. Immunotechnology 1996; 2: 11-20. 167.Levi M, Sallberg M, Ruden U, Herlyn D, Maruyama H, Wigzell H et al. A complementarity determining region synthetic peptide acts as a miniantibody and neutralizes human immunodeficiency virus type 1 in vitro. Proc Natl Acad Sci USA 1993; 90: 4374-4378. 168.Burton D, Barbas C, Persson M, Koenig S, Chanock R, Lerner R. A large array of human monoclonal antibodies to HIV-1 from combinatorial libraries of asymptomatic seropositive individuals. Proc Natl Acad Sci USA 1991; 88: 10134-10137. 169.Barbas CF, Collet TA, Roben P, Binley J, Amberg W, Hoekstra D et al. Molecular profile of the antibody response to HIV-1 as probed by combinatorial libraries. J Mol Biol 1993; 230: 812-823. 170.Barbas CA, Hu D, Dunlop N, Sawyer L, Cababa D, Hendry M et al. In vitro evolution of a neutralizing human antibody to human immunodeficiency virus type 1 to enhance affinity and broaden strain cross- reactivity. Proc Natl Acad Sci USA 1994; 91: 3809-3813. 171.Vancott TC, Polonis VR, Loomis LD, Michael NL, Nara PL, Birx DL. Differential role of V3-Specific antibodies in neutralization assays involving primary and laboratory-adapted isolates of HIV type 1. AIDS Res Hum Retroviruses 1995; 11(12): 1379-1390. 172.D'Zouza MP, Milman G, Bradac JA, McPhee D, Hanson CV, Hendry RM et al. Neutralization of primary HIV-1 isolates by anti-envelope monoclonal antibodies. AIDS 1995; 9(8): 867-874. 173.Conley AJ, Gorny M, Kessler JA, Boots LJ, Osorio-Castro M, Koenig S et al. Neutralization of primary human immunodeficiency virus type 1 isolates by the broadly reactive antibody 447-52D. J Virol 1994; 68(1): 6994-7000. 174.Bou-Habib DC, Roderiquez G, Gravecz T, Berman P, Lusso P, Norcross MA. Cryptic Nature of the envelope V3 region epitopes protects primary monocytotropic human immunodeficiency virus type 1 from antibody neutralization. J Virol 1994; 68(9): 6006-6013. 175.Zolla-Pazner S, Sharpe S. A resting cell assay for improved detection of antibody mediated neutralization of HIV type 1 primary isolates. AIDS Res Hum Retroviruses 1995; 11(12): 1449-1457. 176.Francis DP, Fast P, Harkonen S, McElrath ML, Belshe R, Berman P et al. X International Conference on AIDS, Yokohama, Japan, 1995, Abstract Book 1, Abs 314: 90. 177.Takeda A, Tauzon CU, Ennis FA. Antibody mediated infection by HIV via Fc receptors-mediated entry. Science 1988; 242: 580-583. 178.Robinson W, Montefiori DC, Mitchell WM. Antibody dependent enhancement of human immunodeficiency virus type 1 infection. Lancet 1988; 1: 790-794. 179.Haubrich R, Takeda A, Koff W, Smith G, Ennis F. Studies of antibody-dependent enhancement of human immunodeficiency virus type 1 infection mediated by Fc receptors using sera from recipients of recombinant gp160 experimental HIV-1 vaccine. J Infec Dis 1992; 165: 545-548. 180.Robinson, WE, Gorny MK, Xu JY, Mitchell WM, Zolla-Pazner S. Two immunodominant domains of gp41 bind antibodies which enhance human immunodeficiency virus type 1 infection in vitro. J Virol 1991; 65: 44169-4176. 181.Eaton AM, Ugen KE, Weiner DB, Wildes T, Levy J. An anti gp41 human monoclonal antibody that enhances HIV-1 infection in the absence of complement. AIDS Res Hum Retroviruses 1994; 10(1): 13-18. 182.Jiang S, Lin K, Neurath AR. Enhancement of human immunodeficiency virus type 1 infection by antisera to peptides from the envelope glycoproteins gp120/g41. J Exp Med 1991; 174: 1557-1563. 183.Jiang S, Neurath AR. Potencial risks of eliciting antibodies enhancing HIV-1 infection of monocytic cells by vaccination with V3 loops of unmatched HIV-1 isolates. AIDS 1992; 6: 331-342. 184.Takeda A, Ennis FA, Roninson JE, Ho DD, Debouck C. Characterization of enhancement of HIV-1 infection by human monoclonal antibodies to HIV-1. In: Vaccines 91, 1991, Cold Spring Harbor Laboratory Press: 23-27. 185.Takeda A, Robinson JE, Ho DD, Debouck C, Haigwood NL, Ennis FA. Distinction of human immunodeficiency virus type 1 neutralization and infection enhancement by human monoclonal antibodies to glycoprotein 120. J Clin Invest 1992; 89: 1952-1957. 186.Emini E, Nara PO, Schleif WA, Lewis JA, Davide JP, Lee DR et al. Antibody mediated in vitro neutralization of human immunodeficiency virus type 1 abolishes infectivity for chimpanzees. J Virol 1990; 64(8): 3674-3678. 187.Emini E, Schleif WA, Numberg JH, Conley AJ, Eda Y, Tokiyoshi S et al. Prevention of HIV-1 infection in chimpanzees by gp120 V3 domain specific monoclonal antibody. Nature 1992; 355: 728-730. 188.Safrit J, Fung M, Andrews CA, Braun DG, Sun W, Chang T et al. hu-PBL-SCID mice can be protected from HIV-1 infection by passive transfer of monoclonal antibody to the principal neutralizing determinant of envelope gp120. AIDS 1993; 7: 15-21. 189.Hinkula J, Bratt G, Gilljam G, Nordlund S, Broliden PA, Holmberg V et al. Immunological and virological interactions in patients receiving passive immunotherapy with HIV-1 neutralizing monoclonal antibodies. In: Functionally Important HIV-1 B cell determinants mapped with monoclonal antibodies. Phd Tesis, Stockholm, 1993. 190.Gunthard HF, Gowland, Schupbach, Fung M, Boni J, Liou RS et al. A phase I/IIA clinical study with a chimeric mouse-human monoclonal antibody to V3 loop of human inmunodeficiency virus Type 1 gp120. J Infect Dis 1994; 170: 1384-93. 191.Wahren B, Bratt G, Hinkula J et al. Monoclonal antibodies given as passive treatment to HIV infected individuals. In: Girard M, Valette L, eds. Retroviruses of Human AIDS and related animal diseases. 5e Colloque des Cent Gardes. Paris, 1990, Pasteur Merieux, Abstracts Book: 263-267. 192.Morell A, Terry WD, Waldman TA. Metabolic properties of IgG subclasses in man. J Clin Invest 1970; 49: 673-680. 193.Metlas R, Skerl V, Pongor S, Colombatti A, Veljkovic V. Reactivity of AIDS patient sera with a peptide derived from HIV type 1NY5 glycoprotein 120 V3 loop consensus sequence of collagens.AIDS Res Hum Retroviruses 1994; 10(11): 1425-1426. 194.Grant MD, Weaver MS, Tsoukas C, Hoffmann GW. Distribution of antibodies against denatured collagen in AIDS patients suggest a link between autoimmunity and the immunopathogenesis of AIDS.J Immunol 1990; 144: 1241-1250. 195.Trujillo JR, McLane MF, Lee TH, Essex M. Molecular mimicry between the human immunodeficiency virus type 1 gp120 V3 loop and human brain proteins. J Virol 1993; 67(12): 7711-7715. 196.Douvas A, Takehana Y. Cross-reactivity between autoimmune anti U1 snRNP antibodies and neutralizing epitopes of HIV-1 gp120/1. AIDS Res Hum Retroviruses 1994; 10(3): 253-262. 197.Rakowicz-Szulczynska, Kaczmarski W, Raso V, Steimer K, Durda P. Internalization of anti-gp120 monoclonal antibody and human antibodies by HIV-1 infected lymphocytes. Antibody, Immunoconjugates and Radiopharmaceuticals 1993; 6(4): 209-219. 198.Berberian L, Goodglick L, Kipps TJ, Braun J. Immunoglobulin VH3 gene products: Natural ligands for HIV gp120. Science 1993; 261: 1588-1591. 199.Berberian L, Valles-Ayoub, Sun N, Martinez-Maza, Braun J. A VH clonal deficit in HIV-positive individuals reflects a B-cell maturational arrest. Blood 1991; 78: 175. 200.Kohler H, Muller S, Nara P. Deceptive imprinting in the immune response against HIV-1. Immunol Today 1994; 15(10): 475-478. 201.Metlas R, Veljkovic V. Does the HIV-1 manipulate immune network via gp120 immunoglobulin-like domain involving V3 loop? Vaccine 1995; 13(4): 355-359. 202.Goodglick L, Zevit N, Neshat M, Braun J. Mapping the Ig Superantigen-binding site of HIV-1 gp120. J Immunol 1995; 155(11): 5151-5159. 203.Takahashi H, Cohen J, Hosmalin A, Cease KB, Houghten R, Cornette JL et al. An immunodominant epitope of the human immunodeficiency virus envelope glycoprotein gp160 recognized by class 1 major histocompatibility complex molecule-restricted immune cytotoxic T lymphocytes. Proc Nat Acad Sci USA 1988; 85: 3105. 204.Lasarte J, Sarobe P, Gullon A, Prieto J, Borras-Cuesta F. Induction of cytotoxic lymphocytes in mice against the PND of HIV-1 by immunization with an engineered T-cytotoxic-T-helper synthetic peptide construct. Cell Immunol 1992; 141: 211-218. 205.Martinon F, Grasmasse H, Boutillon C, Chirat F, Deprez B, Guillet JG, Gomard E, Tartar A, Levy JP. Immunization of mice with lipopeptides bypasses the prerequisite for adjuvant immune response of Balb/c mice to HIV-1 envelope glyoprotein. J Immunol 1993; 149(10): 3416-3422. 206.Clerici M, Lucey DR, Zajac RA, Boswell RN, Gebel HM, Takahashi H, Berzofsky JA, and Shearer GM. Detection of CTL specific for synthetic peptides of gp160 in HIV-seropositive individuals. J Immunol 1991; 146: 2214-2219. 207.Achour A, Picard O, M bika JP, Willer A, Snart R, Bizini B et al. Envelope protein and P18IIIB peptide recognized by cytotoxic T-lymphocytes from human immunized with AIDS virus envelope. Vaccine 1993; 11: 699-701. 208.Achour A, Lemhammedi S, Picard O, M bika JP, Zagury JF,Moukrim Z et al. Cytotoxic T lymphocytes specific for HIV-1 gp160 antigen and synthetic peptide in an HLA-A11 immunized individual. AIDS Res Hum Retroviruses 1994; 10(1): 19-25. 209.Takahashi H, Houghten R, Putney SD, Margulies Dh, Moss B, Germain RN et al. Structural requirements for class I MHC molecule-mediated antigen presentation and cytotoxic T-cell recognition of an immunodominant of the human immunodeficiency virus envelope protein. J Exp Med 1989; 170: 2023-2035. 210.Takahashi H, Germain RN, Moss B, Berzofsky JA. An immunodominant class I-restricted cytotoxic Y-lymphocyte determinant of human immunodeficiency virus type 1 induces CD4 class II restricted help for itself. J Exp Med 1990; 17: 571-576. 211.Safrit JY, Lee AY, Andrews CA, Koup RA. A region of the third variable loop of HIV-1 is recognized by HLAB7 restricted CTL from two acute seroconvertors. J Immunol 1994; 153: 3822-3830. 212.Takahashi H, Merli S, Putney SC, Houghten R, Moss B, Germain RN et al. A single amino acid interchange yield reciprocal CTL specificities for HIV-1 gp160. Science 1989; 246: 118-121. 123.Philips RE, Rowland-Jones S, Nixon DF, Gotch FM, Edwards JP, Ogulnesi AO et al. human immunodeficiency virus genetic variation that can escape cytotoxic T cell recognition. Nature 1991; 354: 453-459. 214.Takahashi H, Nakagawa Y, Pendleton CD, Houghten RA, Yokomuro K, Germain RN et al. Induction of broadly cross-reactive cytotoxic T cells recognizing and HIV-1 envelope determinant. Science 1992; 255: 333-336 215.Harris SJ, Gearing AJH, Layton GT, Adams SE, Kingsman AJ. Enhanced proliferative cellular response to HIV-1 V3 peptide and gp120 following immunization with V3-Ty virus like particles. Immunology 1993; 77(3): 315-321. 216.Moore RL, Fox B. Immunization of mice with human immunodeficiency virus glycoprotein gp160 peptide 315-329 induces both class I and class II restricted T cells: Not all cells can respond to whole molecule stimulation. AIDS Res Hum Retroviruses 1993; 9(1): 51-59. 217.Clerici M, Stocks NI, Zajac RA, Boswell RN, Berstein DC, Mann DL et al. Interleukin-2 production used to detect antigenic peptide recognition by T-helper lymphocytes from a symptomatic HIV-seropositive individuals. Nature 1989; 339: 383-385. 218.Clerici M, Tacket CO, Via CS, Lucey DR, Muluk SC, Zajac RA et al. Immunization with subunit human immunodeficiency virus vaccine generates stronger T helper cell immunity than natural infection. Eur J Immunol 1991; 21: 1345-1349. 219.Gorse J, Belshe RB, Newman FK, Frey SE, the NIAID AIDS vaccine clinical trials network. Lymphocyte proliferative responses following immunization with human immunodeficiency virus recombinant gp160. Vaccine 1992; 10(6): 1992. 220.Neurath R, Jiang S, Strick N, Kolbe H, Kieny M, Muchmore E et al. Antibody responses of chimpanzees immunized with synthetic peptides corresponding to full length V3 hypervariable loops of HIV-1 envelope glycoproteins. AIDS Res Hum Retroviruses 1991; 7(10): 813-823. 221.Tolman RL, Bednarek MA, Johnson BA, Leanza WJ, Marburg S, Underwood DJ et al. Cyclic V3 Loop related HIV-1 conjugate vaccines. Synthesis, conformation and immunological properties. Int J Peptide Protein Res 1993; 41: 455-466. 222.Javaherian K, Langlois AJ, La Rosa GJ, Profy AT, Bolognesi DP, Herlihy WC et al. Broadly Neutralizing Antibodies Elicited by the Hipervariable Neutralizing Determinant of HIV-1. Science 1990; 250: 1590. 223.Putney SD, La Rosa GJ, Profy AT, Silver S, Scott CF, Javaherian K et al. HIV-1 principal neutralization determinant elicits broadly neutralizing antibodies. In: Vaccines 91, 1991, Cold Spring Harbor Laboratory. Edited by Brown F, Chanock and RM and Lerner RA. 224.Palker TJ, Matthews TJ, Langlois A, Tanner ME, Martin ME, Scearce RM et al. Polyvalent human immunodeficiency virus synthetic immunogen comprised of envelope gp120 T helper cell sites and B cell neutralization epitopes. J Immunol 1989; 142(10): 612-3619. 225.Hart MK, Palker TJ, Matthews TJ, Langlois AJ, Lerche NW, Martin ME et al. Synthetic peptides containing T and B cell epitopes from human immunodeficiency virus envelope gp120 induce anti-HIV proliferative responses and high titers of neutralizing antibodies in rhesus monkeys. J Immunol 1990; 146(8): 2677-2685. 226.Nardelli B, Lu YA, Shiou D, Delpierre-Defoort C, Profy A, Tam JP. A chemically defined synthetic vaccine model for HIV-1. J Immunol 1992; 148: 914-920. 227.Profy A, Salinas P, Khiroya R, Sax S, Hanham C, Boyd J. Immunogenicity of synthetic peptides corresponding to the V3 region of gp120. IVth Int Conf on AIDS, Berlin, 1993. PO-A29-0688, Abstract Book 1: 249. 228.Cryz SJ, Goldstein H, Pettoello-Mantovani M, Kim A, Furer E, Que JU, Hasler T et al. Human Immunodeficiency virus -1 Principal neutralizing domain peptide-toxin A conjugate vaccine. Vaccine 1995; 13(1): 67-71. 229.Mitsuda S, Nakagawa T, Osada T, Shimamoto T, Nakazato H, Ikai A. A receptor mediated delivery of an HIV-1 derived peptide vaccine. Biochem and Biophysical Res Communications 1993; 194(3): 1155-1160. 230.Defoort JP, Nardelli B, Huang W, Ho D, Tam JP. Macromolecular assemblage in the design of a synthetic AIDS vaccine. Proc Natl Acad Sci USA 1992; 89: 3879-3883. 231.Montero M, Menendez A, Dominguez MC, Navea L, Vilarubia OL, Quintana D et al. Broadly reactive antibodies against a gp120 V3-loop Multi-Epitope Polypeptide neutralize different isolates of human immunodeficiency virus type 1 (HIV-1). Sent to Vaccine for publication. 232.Wagner R, Fliessbach H, Modrow S, vonBrunn A, Wolf H, Boltz T et al. Expression of HIV-1 autologous p55v and p55/gp120-V3 core particles: A new approach in HIV vaccine development. Vaccines 91, 1991: 109-113, Cold Spring Harbor Laboratory Press. 233.Luo LZ, Li Y, Cannon PM, Kim SY, Kang CY. Chimeric gag-V3 virus-like particles of human immunodeficiency virus Induce virus neutralizing antibodies. Proc Natl Acad Sci USA 1992; 89: 21. 234.Fung MS, Sun CRY, Gordon W, Chang NT, Chang TW, Sun NC. Monoclonal anti-idiotypic antibody mimicking the principal neutralizing site in HIV-1 gp120 induces HIV-1 neutralizing antibodies in rabbits. J Immunol 1990; 145(7): 2199-2206.Boudet F, Theze J, Zouali M. Anti-idiotypic antibodies to the third variable domain of gp120 induce an anti HIV-1 antibody response in mice. Virology 1994; 200: 176-188. 235.Kang CY, Nara P, Chamat S, Caralli V, Chen A, Nguyen ML et al. Anti-idiotype antibody elicits broadly neutralizing anti-gp120 antibodies in monkeys. Proc Natl Acad Sci USA 1992; 89: 2546-2550. 236.Fontenot JD, Gatewood JM, Mariappan S, Pau CP, Parekh B, George R et al. human immunodeficiency virus (HIV) antigens: Structure and serology of multivalent human mucin MUC1-HIV V3 chimeric proteins. Proc Natl Acad Sci USA 1995; 92: 315-319. 237.Gorse G, Keefer MC, Belshe RB, Matthews TJ, Forrest BD, Hsieh RH, Koff WC, Hanson CV, Dolin R, Weinhold KJ, Frey SE, Ketler N, Fast PE. A dose ranging study of a prototype synthetic HIV (MN) branched peptide vaccine. J Infect Dis 1996; 173(2): 330-339. 238.Salmon-Ceron D, Excler JL, Sicard D, Blanche P, Finkielstzjen, Gluckman JC et al. Safety and immunogenicity of a recombinant HIV type 1 glycoprotein 160 boosted by a V3 synthetic peptide in HIV-negative volunteers. AIDS Res Hum Retroviruses 1995; 11(12): 1479-1486. Copyright 1997 Elfos Scientiae The following images related to this document are available:Line drawing images[ba97001a.gif] |
| |||||||||

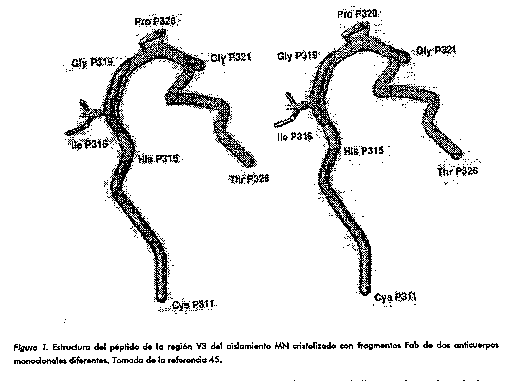{kind=link}