 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
MOLECULAR ANALYSIS OF Neisseria meningitidis CLASS 3 OUTER MEMBRANE PROTEIN IN STRAINS RECOGNIZED BY THE MONOCLONAL ANTIBODY CB-Nm.2 Gerardo Guillen*, Anabel lvarez, Consuelo Nazabal, Maria de Jesus Leal, Luis Manuel Alonso, Alexis Musacchio, Maite Delgado, Osvaldo Reyes and Ricardo Silva
Vaccines Division, Center for Genetic Engineering and Biotechnology, P.O.
Box 6162, Havana, Cuba. Telephone: (53-7) 21 8008. Fax: (53-7) 21 8070.
Code Number:BA97002
Sizes of files:
Text: 48.8K
Graphics: Line drawings (gif) -93K
Photographs (jpg) - 22.8K
ABSTRACT Bactericidal antibodies against outer membrane proteins are crucial to protect against Meningococcal Meningitis. The bactericidal monoclonal antibody (MAb) CB-Nm.2, specific for the class 3 outer membrane protein of Neisseria meningitidis, was assayed in an enzyme linked immuno- sorbent assay (ELISA) with a panel of 86 N. meningitidis strains. Fifty six strains belonging to seven serogroups: A, B, 29E, L, X, Y, Z, and five serotypes: 1, 4, 5, 12, 13, reacted with CB-Nm.2. The porB genes coding four such proteins were cloned and sequenced, and the translated amino acid sequences were compared with five previously published sequences. Sequence alignment revealed a five amino acid region (S/T)VETG located in the main variable region (VR) VR1 which was present in all N. meningitidis strains recognized by CB-Nm.2 and not in the strains which were negative in ELISA. Two synthetic peptides were designed on the basis of the predicted antigenic determinant for strains B385 and H355. Mouse antiserum obtained against the synthetic peptides recognized Neisseria strains in whole cell Dot-blot, but synthetic peptides failed to react with the MAb. The results show that the (S/T)VETG region is present among different serotypes of N. meningitidis and it is probably involved in antigenic recognition by the bactericidal MAb CB- Nm.2. Key words: class 3 protein, porB gene, DNA sequence RESUMEN Los anticuerpos bactericidas contra las proteinas de la membrana externa son decisivos en la proteccion contra la meningitis meningococcica. El anticuerpo monoclonal (AcM) CB-Nm.2 que es bactericida contra Neisseria meningitidis y especifico para la proteina de clase 3 de esta bacteria, se evaluo en ELISA contra un panel de 86 cepas de N. meningitidis. Cincuenta y seis de las cepas, pertenecientes a siete serogrupos: A, B, 29E, L, X, Y, Z, y cinco serotipos: 1, 4, 5, 12, 13, fueron reconocidas por el CB-Nm.2. Los genes porB codificantes para cuatro de estas proteinas se clonaron y secuenciaron. Las secuencias de ADN fueron traducidas a aminoacidos y comparadas con cinco secuencias publicadas con anterioridad. El alineamiento de secuencias mostro una region de cinco aminoacidos (S/T)VETG, localizada en la principal region variable (VR) VR1 que se encontraba presente en todas las cepas de N. meningitidis reconocidas por CB-Nm.2, y no en las cepas que resultaron negativas en el ELISA. Se sintetizaron peptidos que contenian esta region de las cepas B385 y H355. Los sueros de raton obtenidos contra los peptidos sinteticos reconocieron las cepas de Neisseria en Dot de celulas totales, pero los peptidos no fueron reconocidos por el AcM. Los resultados obtenidos muestran que la region (S/T)VETG es el posible sitio de reconocimiento antigenico del AcM bactericida CB-Nm.2 y esta presente en diferentes serotipos de N. meningitidis. Palabras claves: proteina de clase 3, gen porB, secuencia de ADN
Introduction Neisseria meningitidis is a major etiological agent of bacterial meningitis and septicemia, causing one third of the epidemic and endemic bacterial meningitis cases throughout the world (1, 2). The disease produces a fulminant effect with a high mortality rate (3).
There are at least 13 serogroups of N. meningitidis (4, 5), more than 20 different serotypes (6), 17 subtypes, and 8 immunotypes (7). Meningococci are classified into serogroups, serotypes (class 2 and class 3 proteins) and subtypes (class 1 protein) by capsular polysaccharides and by the differences in outer membrane protein (OMP) composition, respectively. The same serotypes are found in groups B, C, Y and W135; the class 3 proteins of serogroup A were found to be antigenically homogeneous and are designated serotype 21 (8).
The epidemiology of the disease shows the coexistence, at the same time, of different serogroups and serotypes in the same geographic area and a high variability of the prevalence of the strains. Nevertheless, strains expressing the class 3 protein are currently the major cause of Meningococcal Meningitis due to serogroup B around the world.
Commercially available polysaccharide vaccines provide limited protection against infection caused by serogroups A and C. The group B polysaccharide is poorly immunogenic in humans (6, 9, 10), due to antigenic similarities with human brain components (11). Several approaches to provide protection against serogroup B using OMP have failed. Only the Cuban vaccine (VA-MENGOC-BC) has proven its efficacy against serogroups B and C (12). So far, attempts to obtain an antimeningococcal vaccine based on the use of a cloned OMP have been hampered by the antigenic variability of these proteins (13) and the fact that most of these antigens fail to induce bactericidal antibodies. To obtain a wider range of protection, more than one antigen will probably have to be included in a vaccine preparation.
It has been shown that class 2 and class 3 OMPs, coded by the porB locus (14), elicit bactericidal antibodies (15), protect against challenge to meningococci in the infant rat infection model (16) and induce high antibody titers after natural infection in humans (17). At the same time, the class 3 antigen is one of the major components of the Cuban and the Norwegian vaccines (12, 18).
In contrast with the class 1 protein, where the subtype specificity of the sera is directed against a well-defined antigenic region (loop 2 or 4), the serotype specificity, in the class 3 protein, changes among different surface-exposed loops in different strains (19).
Because of the variability of the class 3 protein among the N. meningitidis strains, extensive characterization of the specific antigenic determinant involved in the bactericidal reaction should be done in order to select the epidemiologically most representative protein to be included in vaccine preparations. A similar approach has already been used for the class 1 proteins (20, 21). The aim of this study was to show that, in spite of the high variability among the porB genes belonging to different serotypes of N. meningitidis, these genes share antigenic determinants that may be involved in the cross-protective response against N. meningitidis. Materials and Methods Bacterial strains
The N. meningitidis strains used in this study are listed in Table 1. The Meningococcal strain B385 (B:4:P1.15) came originally from a patient with the Meningococcal disease (20). Escherichia coli strains used were XL-1-Blue (22) and HB-101 (23).
--------------------------------------------------------------------------- Table.1. Immunoidetification of class 3 protein in selected strains of Neisseria meningitidis by whole cell ELISA using the MAb CB-Nm.2. ND (non determined), NT (non typeable). --------------------------------------------------------------------------- CV^a Strain number Source Serogroup Subtype Serotype MAb CB-Nm.2 --------------------------------------------------------------------------- 11 IHN2312 NPHI^b W-135 ND ND - 12 IHN36157 NPHI B 15 4 +++ 13 IHN5385 NPHI B 1 4 +++ 16 IHN36117 NPHI B 7 14 - 17 IHN5433 NPHI NT 16 NT ++ 18 IHN5421 NPHI B 16 4 +++ 19 IHN36152 NPHI NT 16 NT - 20 IHN5435 NPHI NT NT NT - 21 IHN5428 NPHI B NT 4 +++ 27 118/89 CPHE^c B 15 ND +++ 29 B4 CPHE B ND ND +++ 44 H355 CPHE B 15 15 - 45 C11 WR CPHE C ND ND - 46 V1-77 CPHE 29E ND ND +++ 54 RHN871 NPHI N. subflava ND ND - 57 RHN869 NPHI N. mucosa ND ND - 60 52 CPHE B. catarralisND ND + 61 I-81 CPHE A ND ND +++ 63 V-75 CPHE Y ND ND ++ 73 H44/76 NPHI B 16 15 - 74 - CPHE X ND ND +++ 75 - CPHE Z ND ND +++ 76 - CPHE H ND ND - 77 - CPHE L ND ND +++ 79 B:14:CPHE CPHE B ND 14 - 81 B:12:CPHE CPHE B ND 12 +++ 89 B:8:CPHE CPHE B ND 8 +/- 92 43-31-1 CPHE B ND 13 + 93 B:4:CPHE CPHE B ND 4 +++ 101 31C2 CPHE B ND ND +++ 102 150C2 CPHE B ND ND +++ 109 M986 CPHE B ND 2.7 - 111 B:11:CPHE CPHE B ND 11 - 113 B385 CPAV^d B 15 4 +++ 125 B:6:CPHE CPHE B ND 6 - 127 B:1:CPHE CPHE B ND 1 - 181 Z90 Achtman^e B ND 9 - 182 882066 Achtman - 4 NT - 183 C2241 Achtman C ND ND - 184 2802 Achtman A ND ND ++ 185 M992 Achtman B 1 5 +++ 186 S3446 Achtman B ND 14 - 191 2959 Achtman B 15 4 +++ 194 84077 Achtman A 3 21 + 195 2996 Achtman B 2 2b - 196 B506 Achtman A ND ND + 197 51 Achtman B 2 2a - 198 S3032 Achtman B 16 12 +++ 199 M982 Achtman B 9 9 - 200 Z222 Achtman I ND ND - 201 Z3756 Achtman A ND ND + 204 02019002 Achtman C ND ND - 205 Z21 Achtman B 16 15 - 208 Z4754 Achtman A ND ND +/- 210 MC19 Achtman C ND ND -+ 212 SBLNK338 Achtman A ND ND + 214 8529 Achtman B 3 15 - 215 190I Achtman B 6 18 - 217 6940 Achtman B 6 19 - 218 B16B6 Achtman B 2 2a - 220 Z14 Achtman B 15 15 - 221 M990 Achtman B 6 6 - 222 M1080 Achtman B 1.7 1 +++ 223 870227 Achtman B 10 4 +++ 224 2802 Achtman A ND ND ++ 225 J117 Achtman A ND ND + 226 C1419 Achtman A ND ND ++ 227 B95 Achtman A ND ND ++ 233 88 Achtman C ND ND - ^a CV : identification number in the Center for Genetic Engineering and Biotechnology collection. ^b NPHI : National Public Health Institute, Helsinki, Finland. ^c CPHE : Provincial Center for Hygiene and Epidemiology, Havana, Cuba. ^d CPAV : Center for Production of Anti-meningococcal Vaccine. Finlay Institute, Havana, Cuba. ^e Dr. Mark Achtman. Max Planck Institute for Molecular Genetic, Berlin, Germany. --------------------------------------------------------------------------- Genomic DNA preparation N. meningitidis strains were grown in Muller-Hinton Broth (OXOID, UK) or brain heart infusion broth (OXOID, UK). The cells were harvested from the culture by low speed centrifugation, resuspended in 8 mL of TE [10 mM Tris-hydroxymethylaminomethane, 1 mM ethylene diamine tetraacetic acid (EDTA) pH 8.0] containing 10 mg/mL lysozyme (Sigma, UK), 0.5 mg/mL pronase E (Merck, FRG) and 1 % sodium duodecil sulphate (SDS) (BDH, UK) and incubated for 1 h at 37 C, followed by an extraction with phenol- chloroform-isoamyl alcohol (25:24:1), the addition of an equal volume of 2- butanol, and a precipitation with 2.5 volumes of absolute ethanol. The tRNA was removed by incubation with 100 ug/mL of RNAse A (Sigma, UK).
Polymerase chain reaction and electrophoresis The polymerase chain reactions (PCRs) were performed with Taq DNA polymerase (Enzibiot, Cuba) using two units per reaction. A reaction mixture containing 25 mM Tris-HCL pH 9.0, 50 mM KCl, 10 mM MgCl, 0.1 % gelatin, 1 mM dithiothreitol (DTT), 200 uM of each dNTP, primers at 1 mM and 1 ug of genomic DNA was incubated in a programmable heat block HYBAID (Cera Labo SA, France) during 30 cycles for 1 min at 95 C (denaturation), 1 min at 55 C (annealing) and 1 min at 72 C (extension reaction) and, the last cycle of extension was carried out for 3 min at 72 C. Then, the reaction mixture was extracted with 100 uL of phenol-chloroform-isoamyl alcohol (25:24:1) and the aqueous phase was washed with diethyl ether saturated with TE. After centrifuging for 1 min, the ether was discarded and the remainder was removed by heating at 50 C for 5 min. The gene amplification product was checked in 0.8 % agarose gel electrophoresis in TA (40 mM Tris-acetate pH 8.0, 1 mM EDTA) at 120 v using 5 uL of each sample. The rest of the samples were separated in 0.8 % low melting temperature (LMT) agarose gels as described by Sambrook et al., (24) and the amplified genes were isolated by the phenol extraction method (24). DNA cloning and sequencing DNA fragments isolated from the LMT agarose gels were ligated within the EcoR V cut pSK vector (BlueScript II SK+, Stratagene, USA) and used to transform the E. coli strain XL-1-Blue. Positive phagemide plaques were selected by alpha-complementation in LB media plates containing 100 ug/mL X-gal and 20 ug/mL IPTG, followed by DNA hybridization using the oligonucleotide P1371 labelled with alpha-dATP^32 as the hybridization probe and the Nco I - BamH I restriction analysis. DNA from each clone was sequenced using the sequenase version 2.0 kit (USB, USA). Restriction enzymes and primers used for PCR amplification and DNA sequencing were from Enzibiot (Cuba). All the procedures were carried out as described by Sambrook et al., (24) and following the instructions given by the manufacturers.
Sequence alignment was done using the CLUSTAL V software (25). Whole cell ELISA Strains were grown overnight at 37 C on chocolate agar plates and then, for a further 7 h at 37 C in 5 mL cultures of a brain heart infusion broth. The bacteria were harvested and suspended in PBS with 0.02 % sodium azide. The optical density (OD) of the suspension, measured at 620 nm, was adjusted to 0.1. One hundred microliters of this suspension (3 x 10^7 cells) were added to individual wells in polystyrene microtiter plates and allowed to evaporate overnight at 37 C. The plates were tested with standard methods (26) using the IgG2b MAb CB-Nm.2 (27) diluted 1:500 in PBS as a primary antibody and the IgG anti-mouse HRPO conjugate (Amersham, UK) as a secondary antibody. The E. coli strain HB 101 and the N. meningitidis strain B385 were used as negative and positive controls, respectively. Peptide synthesis Peptides were synthesized according to the solid-phase method (28) on a 1 mmol/g HPLC resin (4-methylbenzhydrylamine; tert-butyloxycarbonyl, t-Boc; hydrogen fluoride, HF; Fluka) using the t-Boc/Benzyl strategy. Peptide- resin was cleaved with HF using the "Low-High" procedure (29) in the presence of appropriate scavengers, and washed three times with ether. Peptides were extracted with 30 % acetic acid and purified on reverse phase HPLC (Vydac C18, 10 x 250 mm). Immunization and production of antiserum Synthetic peptides were conjugated to the carrier protein keyhole limpet haemocyanin (KLH) as described in Carter (30).
The conjugate was adjuvated with Freund complete adjuvant (SIGMA, USA) and used to immunize, subcutaneosly with one dose of 50 ug, 10 female six week old Balb/c mice. Mice were then boosted with four doses of the same antigen adjuvated in Freund incomplete adjuvant at two week intervals. The animals were bled two weeks after the last immunization and the sera stored at 20 C until required. Dot-blotting One microlitre of the bacterial suspension, equivalent to 0.1 DO620, was spotted onto a hybond-C nitrocellulose membrane filter (Amersham, UK). The membrane was blocked with 5 % low fat milk in PBS for 1 h at 37 C, then washed and incubated for 3 h at 37 C with antipeptide antibodies diluted 1:100 in PBS. After washing, antibody binding was detected using an antimouse peroxidase conjugate (1:1000) (Amersham) for 1 h at room temperature and the chromogen 4-chloro-1-naphtol. All washing steps were performed with PBS containing 0.05 % Tween 20. Results and Discussion The murine bactericidal monoclonal antibody CB-Nm.2 obtained against the N. menigitidis strain B385 (27, 31) was assayed in ELISA with a panel of 86 N. meningitidis strains. The MAb CB-Nm.2 recognized the class 3 protein in 56 strains belonging to seven serogroups: A, B, 29E, L, X, Y and Z and five serotypes: 1, 4, 5, 12 and 13 but did not recognize the serotypes 2, 6, 9, 11, 14, 15, 18 and 19. Included in the ELISA, there were also two strains of serotype 1 from which only one (CV222) was recognized by the MAb CB-Nm.2 (Table 1). Seventeen strains isolated in Cuba were not included in Table 1 due to their similarities to the strain CV113.
For vaccine design, the recognition by the bactericidal MAb CB-Nm.2 of strains belonging to seven different serogroups is quite important, especially for the 11 strains of serogroup A, one of the most frequently isolated strains around the world. Among these strains were isolates from 9 different countries: China (CV194 and CV208), India (CV184 and CV224), England (CV225), Finland (CV227), Sweden (CV212), Sudan (CV201), Gambia (CV226), Brazil (CV196) and the former Soviet Union (CV61). A large amount of strains within the serogroup B (21 of 56 positive strains), with a non determined serotype, were recognized by the MAb CB-Nm.2. This was not unexpected, since they were isolated in Cuba during the 1980-1990 period, as well as the strain B385 used in immunization schedules to obtain the MAb CB-Nm.2; around 95 % of the isolates of this period were classified as B:4:P1.715.
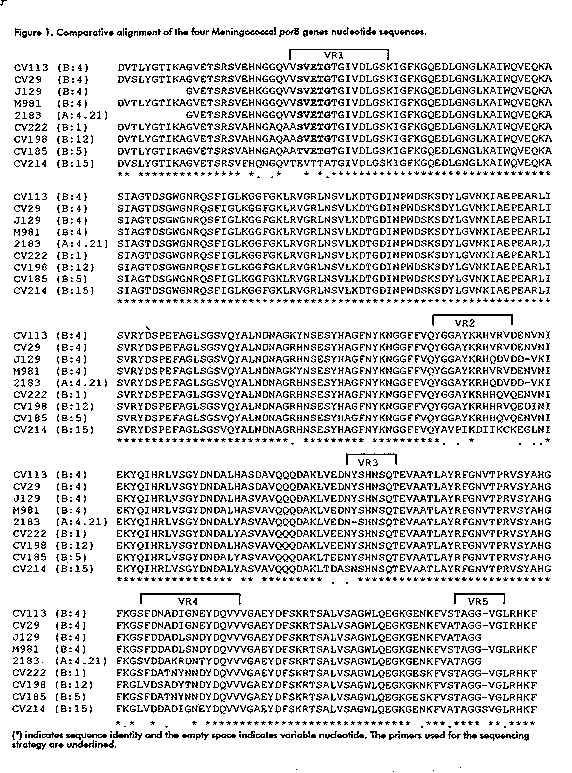
To locate the epitope recognized by the MAb using the comparison of the amino acid sequence, four porB genes (three recognized and one unrecognized by the MAb CB-Nm.2), representative of different serotypes, were amplified and sequenced (Figure 1). The DNA was translated to amino acids for sequence alignment along with previously published DNA sequences from serotypes 1, 4, and 12 (32). Differences between the porB sequences of each serotype and a consensus sequence were determined with the CLUSTAL V software (Figure 2). Figure 1. Comparative alignment of the four Meningococcal porB genes nucleotide sequences.
--------------------------------------------------------------------------
VR1
---------------
CV113 (B:4)
DVTLYGTIKAGVETSRSVEHNGGQVVSVETGTGIVDLGSKIGFKGQEDLGNGLKAIWQVEQKA
CV29 (B:4)
DVSLYGTIKAGVETSRSVEHNGGQVVSVETGTGIVDLGSKIGFKGQEDLGNGLKAIWQVEQKA
J129 (B:4)
GVETSRSVEHKGGQVVSVETGTGIVDLGSKIGFKGQEDLGNGLKAIWQVEQKA
M981 (B:4)
DVTLYGTIKAGVETSRSVEHNGGQVVSVETGTGIVDLGSKIGFKGQEDLGNGLKAIWQVEQKA
2183 (A:4.21)
GVETSRSVEHNGGQVVSVETGTGIVDLGSKIGFKGQEDLGNGLKAIWQVEQKA
CV222 (B:1)
DVTLYGTIKAGVETSRSVAHNGAQAASVETGTGIVDLGSKIGFKGQEDLGNGLKAIWQVEQKA
CV198
(B:12)DVTLYGTIKAGVETSRSVAHNGAQAASVETGTGIVDLGSKIGFKGQEDLGNGLKAIWQVEQK
A
CV185 (B:5)
DVTLYGTIKAGVETSRSVAHNGAQAATVETGTGIVDLGSKIGFKGQEDLGNGLKAIWQVEQKA
CV214 (B:15)DVSLYGTIKAGVETSRSVFHQNGQVTEVTTATGIVDLGSKIGFKGQEDLGNGLKAIWQVEQKA
** *************** *. .* * *.********************************
CV113 (B:4) SIAGTDSGWGNRQSFIGLKGGFGKLRVGRLNSVLKDTGDINPWDSKSDYLGVNKIAEPEARLI
CV29 (B:4) SIAGTDSGWGNRQSFIGLKGGFGKLRVGRLNSVLKDTGDINPWDSKSDYLGVNKIAEPEARLI
J129 (B:4) SIAGTDSGWGNRQSFIGLKGGFGKLRVGRLNSVLKDTGDINPWDSKSDYLGVNKIAEPEARLI
M981 (B:4) SIAGTDSGWGNRQSFIGLKGGFGKLRVGRLNSVLKDTGDINPWDSKSDYLGVNKIAEPEARLI
2183(A:4.21)SIAGTDSGWGNRQSFIGLKGGFGKLRVGRLNSVLKDTGDINPWDSKSDYLGVNKIAEPEARLI
CV222 (B:1) SIAGTDSGWGNRQSFIGLKGGFGKLRVGRLNSVLKDTGDINPWDSKSDYLGVNKIAEPEARLI
CV198 (B:12)SIAGTDSGWGNRQSFIGLKGGFGKLRVGRLNSVLKDTGDINPWDSKSDYLGVNKIAEPEARLI
CV185 (B:5) SIAGTDSGWGNRQSFIGLKGGFGKLRVGRLNSVLKDTGDINPWDSKSDYLGVNKIAEPEARLI
CV214 (B:15)SIAGTDSGWGNRQSFIGLKGGFGKLRVGRLNSVLKDTGDINPWDSKSDYLGVNKIAEPEARLI
***************************************************************
VR2
-----------
CV113 (B:4) SVRYDSPEFAGLSGSVQYALNDNAGKYNSESYHAGFNYKNGGFFVQYGGAYKRHVRVDENVNI
CV29 (B:4) SVRYDSPEFAGLSGSVQYALNDNAGRHNSESYHAGFNYKNGGFFVQYGGAYKRHVRVDENVNI
J129 (B:4) SVRYDSPEFAGLSGSVQYALNDNAGRHNSESYHAGFNYKNGGFFVQYGGAYKRHQDVDD-VKI
M981(B:4) SVRYDSPEFAGLSGSVQYALNDNAGKYNSESYHAGFNYKNGGFFVQYGGAYKRHVRVDENVNI
2183(A:4.21)SVRYDSPEFAGLSGSVQYALNDNAGRHNSESYHAGFNYKNGGFFVQYGGAYKRHQDVDD-VKI
CV222 (B:1) SVRYDSPEFAGLSGSVQYALNDNAGRHNSESYHAGFNYKNGGFFVQYGGAYKRHHQVQENVNI
CV198 (B:12)SVRYDSPEFAGLSGSVQYALNDNAGRHNSESYHAGFNYKNGGFFVQYGGAYKRHHRVQEDINI
CV185 (B:5) SVRYDSPEFAGLSGSVQYALNDNAGRHNSESYHAGFNYKNGGFFVQYGGAYKRHHQVQENVNI
CV214 (B:15)SVRYDSPEFAGLSGSVQYALNDNAGRHNSESYHAGFNYKNGGFFVQYAVPIKDIIKCKEGLNI
*************************. ********* **********. . * . ..*
VR3
--------
CV113 (B:4) EKYQIHRLVSGYDNDALHASDAVQQQDAKLVEDNYSHNSQTEVAATLAYRFGNVTPRVSYAHG
CV29 (B:4) EKYQIHRLVSGYDNDALHASDAVQQQDAKLVEDNYSHNSQTEVAATLAYRFGNVTPRVSYAHG
J129 (B:4) EKYQIHRLVSGYDNDALHASVAVQQQDAKLVEDNYSHNSQTEVAATLAYRFGNVTPRVSYAHG
M981 (B:4) EKYQIHRLVSGYDNDALHASVAVQQQDAKLVEDNYSHNSQTEVAATLAYRFGNVTPRVSYAHG
2183(A:4.21)EKYQIHRLVSGYDNDALYASVAVQQQDAKLVEDN-SHNSQTEVAATLAYRFGNVTPRVSYAHG
CV222 (B:1) EKYQIHRLVSGYDNDALYASVAVQQQDAKLVEENYSHNSQTEVAATLAYRFGNVTPRVSYAHG
CV198 (B:12)EKYQIHRLVSGYDNDALHASVAVQQQDAKLVEENYSHNSQTEVAATLAYRFGNVTPRVSYAHG
CV185 (B:5) EKYQIHRLVSGYDNDALYASVAVQQQDAKLTEENYSHNSQTEVAATLAYRFGNVTPRVSYAHG
CV214 (B:15)EKYQIHRLVSGYDNDALYASVAVQQQDAKLTDASNSHNSQTEVAATLAYRFGNVTPRVSYAHG
***************** ** ********* . . ****************************
VR4 VR5
--------------- --------
CV113 (B:4) FKGSFDNADIGNEYDQVVVGAEYDFSKRTSALVSAGWLQEGKGENKFVSTAGG-VGLRHKF
CV29 (B:4) FKGSFDNADIGNEYDQVVVGAEYDFSKRTSALVSAGWLQEGKGENKFVATAGG-VGIRHKF
J129 (B:4) FKGSFDDADLSNDYDQVVVGAEYDFSKRTSALVSAGWLQEGKGENKFVATAGG
M981 (B:4) FKGSFDDADLSNDYDQVVVGAEYDFSKRTSALVSAGWLQEGKGENKFVSTAGG-VGLRHKF
2183(A:4.21)FKGSVDDAKRDNTYDQVVVGAEYDFSKRTSALVSAGWLQEGKGENKFVATAGG
CV222 (B:1) FKGSFDATNYNNDYDQVVVGAEYDFSKRTSALVSAGWLQEGKGESKFVSTAGG-VGLRHKF
CV198 (B:12)FRGLVDSADYTNDYDQVVVGAEYDFSKRTSALVSAGWLQEGKGKNKFVSTAGG-VGLRHKF
CV185 (B:5) FKGSFDATNYNNDYDQVVVGAEYDFSKRTSALVSAGWLQEGKGESKFVSTAGG-VGLRHKF
CV214 (B:15)FKGLVDDADIGNEYDQVVVGAEYDFSKRTSALVSAGWLQEGKGENKFVATAGGSVGLRHKF
*.* * . * ****************************** .***.**** **.****
(*) indicates sequence identity and the empty space indicates variable
nucleotide. The primers used for the sequencing strategy are underlined.
---------------------------------------------------------------------------
Figure 2a and 2B. Alignment of the amino acid sequences of the class 3 proteins from nine selected strains. The serogroup and serotype are indicated in parenthesis. The brackets show the VR and the conserved amino acid in the VR1 of the strains recognized by the MAb CB-Nm.2 in ELISA is indicated in bold-face letters. The N- and C-terminal sequences of strains J129 and 2183 were not available from the original reference. These sequence data appear in the EMBL Nucleotide Sequence Data Library under the access numbers: X79464 (CV113), X78579 (CV29), X67933 (J129), X65531 (M981), X67934 (2183), X65530 (M1080), X65534 (S3032), X96496 (CV185), X81048 (CV214).
---------------------------------------------------------------------------
1 10 20 30 40 50 60 70
CV113
GACGTTACCCTGTACGGCACCATCAAAGCCGGCGTAGAAACTTCCCGCTCTGTAGAGCACAATGGAGGTCAGGTG
G
CV029
GACGTTACCCTGTACGGCACCATCAAAGCCGGCGTAGAAACTTCCCGCTCTGTAGAGCACAATGGAGGTCAGGTG
G
CV185
GACGTTACCCTGTACGGCACCATCAAAGCCGGCGTAGAAACTTCCCGCTCCGTAGCTCACAATGGAGCTCAGGCG
G
CV214
GATGTCAGCCTGTACGGCACCATCAAAGCCGGCGTAGAAACTTCCCGCTCTGTATTTCACCAGAACGGCCAAGTT
A
** ** * ****************************************** *** *** * * ** *
80 90 100 110 120 130 140 150
CV113
TTAGCGTTGAAACCGGTACCGGCATCGTTGATTTGGGTTCAAAAATCGGCTTCAAAGGCCAAGAAGACCTCGGTA
A
CV029
TTAGCGTTGAAACCGGTACCGGCATCGTTGATTTGGGTTCAAAAATCGGCTTCAAAGGCCAAGAAGACCTCGGTA
A
CV185
CTACGGTTGAAACCGGTACCGGCATCGTTGATTTGGGTTCGAAAATCGGCTTCAAAGGCCAAGAAGACCTCGGTA
A
CV214
CTGAAGTTACAACCGCTACCGGCATCGTTGATTTGGGTTCGAAAATCGGCTTCAAAGGCCAAGAAGACCTCGGTA
A
* *** ***** ************************ *********************************
160 170 180 190 200 210 220
CV113
CGGTCTGAAAGCCATTTGGCAGGTTGAGCAAAAGGCATCTATCGCCGGTACTGACTCCGGTTGGGGCAACCGCCA
A
CV029
CGGTCTGAAAGCCATTTGGCAGGTTGAGCAAAAGGCATCTATCGCCGGTACTGACTCCGGTTGGGGCAACCGCCA
A
CV185
CGGCCTGAAAGCCATTTGGCAGGTTGAGCAAAAGGCATCTATCGCCGGTACTGACTCCGGTTGGGGCAACCGCCA
A
CV214
CGGCCTGAAAGCCATTTGGCAGGTTGAGCAAAAAGCATCTATCGCCGGTACTGACTCCGGTTGGGGCAACCGCCA
A
*** ** ********************************************************************
230 240 250 260 270 280 290 300
CV113
TCCTTCATCGGTTTGAAAGGCGGCTTCGGTAAATTGCGCGTCGGCCGTTTGAACAGCGTCCTGAAAGACACCGGC
G
CV029
TCCTTCATCGGTTTGAAAGGCGGCTTCGGTAAATTGCGCGTCGGCCGTTTGAACAGCGTCCTGAAAGACACCGGC
G
CV185
TCCTTCATCGGTTTGAAAGGCGGCTTCGGTAAATTGCGCGTCGGCCGTTTGAACAGCGTCCTGAAAGACACCGGC
G
CV214
TCCTTCATCGGCTTGAAAGGCGGCTTCGGTAAATTGCGCGTCGGTCGTTTGAACAGCGTCCTGAAAGACACCGGC
G
***************************************************************************
310 320 330 340 350 360 370 380
CV113
ACATCAATCCTTGGGATAGCAAAAGCGACTATTTGGGTGTAAACAAAATTGCCGAACCCGAAGCACGCCTCATTT
C
CV029
ACATCAATCCTTGGGATAGCAAAAGCGACTATTTGGGTGTAAACAAAATTGCCGAACCCGAGGCACGCCTCATTT
C
CV185
ACATCAATCCTTGGGATAGCAAAAGCGACTATTTGGGTGTAAACAAAATTGCCGAACCCGAAGCACGCCTCATTT
C
CV214
ACATCAATCCTTGGGATAGCAAAAGCGACTATTTGGGTGTAAACAAAATTGCCGAACCCGAGGCACGCCTCATTT
C
***************************************************************************
390 400 410 420 430 440 450
CV113
CGTACGCTACGATTCTCCCGAATTTGCCGGCCTCAGCGGCAGCGTACAATACGCGCTTAACGACAATGCAGGCAA
A
CV029
CGTACGCTACGATTCTCCCGAATTTGCCGGCCTCAGCGGCAGCGTACAATACGCGCTTAACGACAATGCAGGCAG
A
CV185
CGTACGCTACGATTCTCCCGAATTTGCCGGCCTCAGCGGCAGCGTACAATACGCGCTTAACGACAATGCAGGCAG
A
CV214
CGTACGCTACGATTCTCCCGAATTTGCCGGCCTCAGCGGCAGCGTACAATACGCGCTTAACGACAATGCAGGCAG
A
***************************************************************************
460 470 480 490 500 510 520 530
CV113
TATAACAGCGAATCTTACCACGCCGGCTTCAACTACAAAAACGGCGGCTTCTTCGTGCAATATGGCGGTGCCTAT
A
CV029
CATAACAGCGAATCTTACCACGCCGGCTTCAACTACAAAAACGGTGGCTTCTTCGTGCAATATGGCGGTGCCTAT
A
CV185
CATAACAGCGAATCTTACCACGCCGGCTTCAACTACAAAAACGGCGGCTTCTTCGTGCAATATGGCGGTGCCTAT
A
CV214
CATAACAGCGAATCTTACCACGCCGGCTTCAACTACAAAAACGGTGGCTTCTTCGTGCAATATGGCGGTGCCTAT
A
******************************************* *******************************
540 550 560 570 580 590 600
CV113
AAAGACATGTGCGGGTGGATGAGAACGTGAATATTGAGAAATACCAGATTCACCGTTTGGTCAGCGGTTACGACA
A
CV029
AAAGACATGTGCGGGTGGATGAGAACGTGAATATTGAGAAATACCAGATTCACCGTTTGGTCAGCGGTTACGACA
A
CV185
AAAGACATCATCAAGTGCAAGAGAACGTGAATATTGAGAAATACCAGATTCACCGTTTGGTCAGCGGTTACGACA
A
CV214
AAAGACATCATCAAGTGCAAGAGGGCTTGAATATTGAGAAATACCAGATTCACCGTTTGGTCAGCGGTTACGACA
A
********** * *** * *** ***************************************************
610 620 630 640 650 660 670 680
CV113
TGATGCCCTGCACGCTTCCGATGCCGTACAGCAACAAGATGCCAAATTGGTTGAAGACAATTATTCGCACAACTC
T
CV029
TGATGCCCTGCACGCTTCCGATGCCGTACAGCAACAAGATGCCAAATTGGTTGAAGACAATTATTCGCACAACTC
T
CV185
TGATGCCCTGTACGCTTCCGTAGCCGTACAGCAACAAGACGCGAAACTGACTGAAGAAAATTATTCGCACAACTC
T
CV214
TGATGCCCTGTACGCTTCCGTAGCCGTACAGCAACAAGACGCGAAACTGACTGATGCTTCCAATTCGCACAACTC
T
********* ********* ***************** ** *** ** *** * **************
690 700 710 720 730 740 750 760
CV113
CAAACCGAAGTTGCCGCTACCTTGGCATACCGCTTCGGCAACGTAACGCCCCGCGTTTCTTACGCCCACGGCTTC
A
CV029
CAAACCGAAGTTGCCGCTACCTTGGCATACCGCTTCGGCAACGTAACGCCCCGCGTTTCTTACGCCCACGGCTTC
A
CV185
CAAACCGAAGTTGCCGCTACCTTGGCATACCGCTTCGGCAACGTAACGCCCCGCGTTTCTTACGCCCACGGCTTC
A
CV214
CAAACCGAAGTTGCCGCTACCTTGGCATACCGCTTCGGCAACGTAACGCCCCGAGTTTCTTACGCCCACGGCTTC
A
***************************************************************************
770 780 790 800 810 820 830
CV113
AAGGCTCGTTTGATAATGCAGACATAGGCAACGAATACGACCAAGTGGTTGTCGGTGCGGAATACGACTTCTCCA
A
CV029
AAGGCTCGTTTGATGATGCAGACTTAAGCAACGATTACGACCAAGTGGTTGTCGGTGCGGAATACGACTTCTCCA
A
CV185
AAGGCTCGTTTGATGCTACAAACTACAACAACGATTACGACCAAGTGGTTGTCGGTGCGGAATACGACTTCTCCA
A
CV214
AAGGTTTGGTTGATGATGCAGACATAGGCAACGAATACGACCAAGTGGTTGTCGGTGCGGAATACGACTTCTCCA
A
**** * * ***** * ** ** ***********************************************
840 850 860 870 880 890 900 910
CV113
ACGCACTTCTGCCTTGGTTTCTGCCGGTTGGTTGCAAGAAGGCAAAGGCGAAAACAAATTCGTATCGACTGCCGG
C
CV029
ACGCACTTCTGCCTTGGTTTCTGCCGGTTGGTTGCAAGAAGGCAAAGGCGAAAACAAATTCGTATCGACTGCCGG
C
CV185
ACGCACTTCTGCCTTGGTTTCTGCCGGTTGGTTGCAAGAAGGCAAAGGCGAAAGCAAATTCGTATCGACTGCCGG
C
CV214
ACGCACTTCTGCCTTGGTTTCTGCCGGTTGGTTGCAAGAAGGCAAAGGCGAAAACAAATTCGTAGCGACTGCCGG
C
***************************************************************************
920 930 940
CV113 GGT---GTCGGTTTGCGCCACAAATTCTAA
CV029 GGT---GTCGGTATTCGCCACAAATTCTAA
CV185 GGT---GTCGGTTTGCGCCACAAATTCTAA
CV214 GGTTCCGTCGGTCTGCGCCACAAATTCTAA
*** ****** * ***************
---------------------------------------------------------------------------
The genomic DNA was isolated for PCR amplification, as described in Materials and Methods. The porB genes were amplified using primers from the N-terminal and the C-Terminal constant regions, selected on the basis of the porB gene sequence published previously (33). The primers designed for PCR amplification were:
P1371 N-Terminal
5' TTCCATGGACGTTACCCTGTACGGC 3' Nco I
------
P1372 C-Terminal
5' ATGGATCCTTAGAATTTGTGGCGCAGACC 3' BamH I
------
Primers 1371 and 1372 were designed to include the Nco I and BamH I restriction sites with the aim of cloning the isolated genes in an expression vector. The Nco I and BamH I sites were also used for restriction analysis.
Amplified DNA fragments were ligated within the EcoR V cut pSK vector used to transform the E. coli strain XL-1-Blue and screened for positive clones as described in Materials and Methods.
The following set of primers, located in the constant regions within the gene were constructed for DNA sequencing.
P1494 5' TTGAAAGGCGGCTTCGG 3' P1495 5' CAGGGCATCATTGTCGT 3' The location of these primers is shown in Figure 1. Primer P1494 is oriented to the 3' end and primer P1495 to the 5' end. The primers SK and KS located in the vector, flanking the cloning site, were also used.
The topological model for the class 3 protein proposed by van der Ley et al., (34), based on the model of porin proteins, shows eight exposed loops (19). The five VRs identified from our sequence alignment coincide with the loops I, V, VI, VII, and VIII, as previously shown by others (19, 35). There are some minor amino acid changes, before the VR2 and after the VR5, not included within the VRs, because they are located in the transmembrane region without any antigenic relevance. Comparing the VRs 1 to 5, we identified the amino acid sequence (S/T)VETG, where S/T is a conservative substitution, as the only sequence present in all the strains recognized by the MAb CB-Nm.2 and not present in the strains of serotype 15, which did not react with this MAb. To confirm this finding, two peptides were synthesized:
C12 GQVVSVETGTGIVDC C13 GQVTEVTTATGIVDC These peptides were designed following the serotype 4 (C12) and serotype 15 (C13) amino acid sequences, as determined for strains B385 and H355, respectively. The proposed recognition sequence for the MAb CB-Nm.2 is shown in bold-face letters.
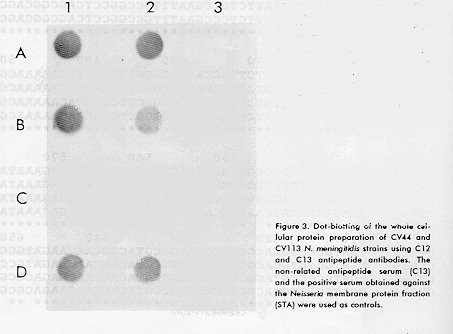
This epitope is not present in serotype 15. Mouse antiserum obtained against the synthetic peptides recognized Neisseria strains serotype 4 and serotype 15 in whole cell Dot-blot (Figure 3), confirming that this region is exposed in the outer membrane of N. meningitidis. The cross-reactivity of the antipeptide sera with both strains is probably due to the conservative amino acids present in C12 and C13. The serum cross-reactivity was also observed against the synthetic peptides in ELISA (data not shown), but synthetic peptides failed to react with the MAb. In Figure 2, the sequence homology shows a high variability rate even within the VRs of serotype 4, but the epitope for the MAb CB-Nm.2 is widely conserved among the 29 serotype 4 strains tested. The high variability within the VR2 of serotype 4, was previously observed by Zapata et al., (35), concluding that this region is not involved in forming the serotype 4 determinant. We also found a wide variability in other VRs.
Figure 3. Dot-blotting of the whole cellular protein preparation of CV44 and CV113 N. meningitidis strains using C12 and C13 antipeptide antibodies. The non-related antipeptide serum (C13) and the positive serum obtained against the Neisseria membrane protein fraction (STA) were used as controls.
It is worth notice that the sequence reported by Bash et al., (19) for strain Cu385 differs in VR3 for the strain B385 reported here. We can add that the serotype determinant for serotype 4 is neither on VR3 or VR4; coinciding with Zapata et al., (35) in VR1 as the most probable region to locate the serotype determinant.
Delvig et al., (36) have also shown that most of the human antibodies developed against the class 3 protein after vaccination with the Norwegian group B vesicle vaccine are directed against the VR1 continuous epitope (36).
We did not find bactericidal activity of the antipeptide antiserum against the homologous strain; only one out of five mice sera assayed showed bactericidal activity at a low dilution (data not shown). The peptide C12 even failed to react with the MAb in ELISA (data not shown). The same situation has been observed with the class 1 OMP, where antibodies against the linear peptide reacted poorly with the native protein in outer membranes and were non-bactericidal. Nevertheless, the same epitope presented on a cyclic peptide elicited bactericidal antibodies (37, 38). The same result was obtained by Christodoulides and Heckels (39), presenting the linear epitopes in multiple antigen peptides with a defined Th-cell epitope from the tetanus toxin.
Then, the bactericidal epitope recognized by the MAb CB-Nm.2 on the class 3 protein as well as the epitopes of the class 1 protein are located in loops exposed in the outer membrane, and in spite of their sequential character, they should have some conformational requirement to elicit bactericidal antibodies. Another possibility is that, in contrast to the class 1 protein, the bactericidal epitopes in the class 3 protein are not sequential but share amino acid determinants located in different regions of the protein, having strong conformational components, and the (S/T)VETG region shown here might interact with another part of the protein to conform the antigenic determinant. This could be supported by the fact that the SVETG region is present in the strains S3446, 190I, and 6940 belonging to serotypes 14, 18, and 19, respectively (19); such serotypes, even from different strains, did not react with the MAb CB-Nm.2 in the whole cell ELISA.
In conclusion, we have shown by sequence comparison the high heterogeneity among the strains recognized by the bactericidal MAb CB-Nm.2, suggesting that the epitope for the bactericidal MAb should have a strong conformational component. We also predicted the determinant that may be involved in this epitope in the strains belonging to different serotypes of N. meningitidis. Further characterization, structural analysis, and evaluation of the bactericidal activity of CB-Nm.2 against the other strains recognized in ELISA should be done to define the bactericidal antigenic determinant of the class 3 protein recognized by the MAb CB- Nm.2.
References 1. Peltola H. Meningococal disease: still with us. Rev Infect Dis 1983; 5:71-86. 2. Schwartz B, Moore PS, Broome CV. Global epidemiology of Meningococcal disease. Clin Microbiol Rev 1989; 2: S118-S124.
3. Broome CV. Group A meningococcus: epidemiology and development of a protein polysaccharide conjugate vaccine. In: Achtman M. et al., editor. Neisseriae 1990. Seventh International Conference on Pathogenic Neisseriaceae, Berlin, GFR, September 9-14, Berlin; New York: de Gruyter, 1991: 17-24.
4. Fredlund H. Serum factors and polymorphonuclear leukocytes in human host defense against Neisseria meningitidis. Studies of interactions with special reference to a chemiluminometric technique. Scan J Infect Dis 1993; suppl. 87:72.
5. Verheul AFM, Snippe H, Poolman JT. Meningococcal lipopolysaccharide: virulence factor and potential vaccine component. Microb Rev 1993;57:34- 49.
6. Frasch CE. Status of a group B Neisseria meningitidis vaccine. Eur J Clin Microb 1985;4:533.
7. Hart CA, Rogers TRF. Meningococcal disease. J Med Microbiol 1993;39:3- 25.
8. Frasch CE, Zollinger WD, Poolman JT. Serotype antigens of Neisseria meningitidis and a proposed scheme for designation of serotypes. Rev Infect Dis 1985;7:504-10.
9. Wyle FA, Artenstein MS, Brandt BL, Tramont EC, Kasper DL, Altieri PL et al. Immunologic response of man to group B meningococcal polysaccharide vaccines. J Infect Dis 1972;120:514-22.
10. Frasch CE. Vaccines for prevention of meningococcal disease. Clin Microbiol Rev 1989; suppl. 2:1345-85.
11. Finne J. Polysialic acid: a glycoprotein carbohydrate involved in neural adhesion and bacterial meningitis. Trends Biochem Soc 1985;10:129- 32.
12. Sierra GV, Campa C, Garcia L, Sotolongo F, Izquierdo L. Efficacy evaluation of the Cuban vaccine VA-MENGOC-BC against disease caused by serogroup B Neisseria meningitidis. In: Achtman M et al., editor. Neisseriae 1990. Seventh International Conference on Pathogenic Neisseriae, Berlin, GFR, September 9-14, Berlin; New York: de Gruyter, 1991:129-34.
13. Meyer TF. Pathogenic Neisseriae - a model of bacterial virulence and genetic flexibility. Zbl BaKt 1990;274:135-54.
14. Hitchcock PJ. Unified nomenclature for pathogenic Neisseria species. Clinical Microb Rev 1989; 2: S64-S65.
15. Poolman JT, Hopman CTP, Zanen HC. Immunogenicity of meningococcal antigens as detected in patient sera. Infect Imm 1983;40:398-406.
16. Saukkonen K, Abdillahi H, Poolman JT, Leinonen M. Protective efficacy of monoclonal antibodies to class 1 and class 3 outer membrane proteins of Neisseria meningitidis B:15:P1.16 in infant rat infection model; new prospects for vaccine development. Microbial Pathogen 1987;3:261-7.
17. Guttorsen HK, Wetzler LM, Naess A. Humoral immune response to class 3 outer membrane protein during the course of meningococcal disease. Infect Immun 1993;61:4734-42.
18. Bjune G, Gronnesby JK, Hoiby EA, Closs O, Nokleby H. Results of an efficacy trial with an outer membrane vesicle vaccine against systemic serogroup B meningococcal disease in Norway. NIPH Ann 1991;14:125-32.
19. Bash MC, Lesiak KB, Banks S, Frasch CE. Analysis of Neisseria meningitidis class 3 outer membrane protein gene variable regions and type identification using genetic techniques. Infect Imm 1995;63:1484- 90.
20. Guillen G, lvarez A, Lemos G, Paredes T, Silva R, Martin A. Comparison of the DNA sequence of nine different genes for the class 1 outer membrane protein from Neisseria meningitidis. Biotecnologia Aplicada 1993; 10: 108-13.
21. van der Ley P, van der Biezen J, Poolman JT. Construction of Neisseria meningitidis strains carrying multiple chromosomal copies of the porA gene for use in the production of a multivalent outer membrane vesicle vaccine. Vaccine 1995; 13: 401-7.
22.Bullock WO, Fernandez JM, Short JM. XL1-Blue: a high efficiency plasmid transforming recA Escherichia coli strain with beta- galactosidase selection. BioTechniques 1987;5:376-9.
23. Boyer HW, Roulland-Dussoix D. A complementation analysis of the restriction and modification of DNA in Escherichia coli. J Mol Biol 1969;41:459.
24. Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual, 2^nd ed. Cold Spring Harbor Laboratory Press, 1989.
25. Higgins DG, Bleasby AJ, Fuchs R. CLUSTAL V: improved software for multiple sequence alignment. CABIOS 1991;5:151-3.
26. Rosenqvist E, Wedege E, Hoiby A, Froholm O. Serogroup determination of Neisseria meningitidis by whole-cell ELISA, dot-blotting, and agglutination. APMIS 1990;98:501-6.
27. Cruz S, Fernandez de Cossio ME, del Valle J, Nazabal C, Ohlin M, Gavilondo JV. Monoclonal antibodies against P1, P3, and 31 Kda outer membrane proteins of Neisseria meningitidis. In: Gavilondo et al., editors. Advances in Modern Biotechnology, Book of Short Reports of the Congress Biotecnologia Habana 92; 1992 June 8-12; Cuba: CIGB. Havana 1992;1:13.26.
28. Houghten RA, DeGraw ST, Bray MK, Hoffmann SR, Frizzell ND. Simultaneous multiple peptide synthesis: the rapid preparation of large numbers of discrete peptides for biological, immunological, and methodological studies. BioTechniques 1986;4:522-6.
29. Tam JP, Heath WF, Merrifield RB. SN2 Deprotection of synthetic peptides with a low concentration of HF in dimethyl sulfide: evidence and application in peptide synthesis. J Am Chem Soc 1983;105:6442-55.
30. Carter JM. Techniques for conjugation of synthetic peptides to carrier molecules. In: Methods in Molecular Biology. Eds. B.M. Dunn and M.W. Pennington - Humana Press Inc 1994;36:155.
31. Vazquez EJ, Ayala M, Danielson L, Fernandez ME, Cruz S, Nazabal C et al. DNA sequences of two bactericidal IgG2B monoclonal antibodies specific for class 3 outer membrane protein of Neisseria meningitidis B:4:P1.15. Biotecnologia Aplicada 1993;10:119-24.
32. Feavers IM, Suker J, McKenna AJ, Heath AB, Maiden MCJ. Molecular analysis of the serotyping antigens of Neisseria meningitidis. Infect Imm 1992; 60:3620-9.
33. Wolff K, Stern A. The class 3 outer membrane protein (PorB) of Neisseria meningitidis: gene sequence and homology to the gonococcal porin PIA. FEMS Microbiol Lett 1991;83179-86.
34. van der Ley P, Heckels JE, Virji M, Hoogerhout P, Poolman JT. Topology of Outer Membrane Porins in Pathogenic Neisseria spp. Infect Imm 1991;59: 2963-71.
35. Zapata GA, Vann WF, Rubinstein Y, Frasch CE. Identification of variable region differences in Neisseria meningitidis class 3 protein sequences among five group B serotypes. Mol Microbiol 1992; 6:3493-9.
36. Delvig A, Wedege E, Caugant DA, Dalseg R, Kolberg J, Achtman M, Rosenqvist E. A linear B-cell epitope on the class 3 outer-membrane protein of Neisseria meningitidis recognized after vaccination with the Norwegian group B outer-membrane vesicle vaccine. Microbiology 1995;141:1593-600.
37. Christodoulides M, McGuinness BT, Heckels JE. Immunization with a synthetic peptide containing epitopes of the class 1 outer-membrane protein of Neisseria meningitidis: production of bactericidal antibodies on immunization with a cyclic peptide. J Gen Microbiol 1993; 139:1729-38.
38. Hoogerhout P, Donders EMLM, van Gaans-van den Brink JAM, Kuipers B, Brugghe HF, van Unen LMA et al. Conjugates of synthetic cyclic peptides elicit bactericidal antibodies against a conformational epitope on a class 1 outer membrane protein of Neisseria meningitidis. Infect Imm 1995;63:3473-8.
39. Christodoulides M, Heckels JE. Immunization with a multiple antigen peptide containing defined B- and T-cell epitopes: production of bactericidal antibodies against group B Neisseria meningitidis. Microbiology 1994;140: 2951-60.
Copyright 1997 Elfos Scientiae The following images related to this document are available:Photo images[ba97002d.jpg]Line drawing images[ba97002a.gif] [ba97002c.gif] [ba97002b.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}