 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Influence of the stabilising sequence on the expression of multi-epitope polypeptides of HIV-1 in Escherichia coli
*Dania Vazquez, Marinieve Montero, Carlos A Duarte and Alfredo Menendez Vaccine Division, Center for Genetic Engineering and Biotechnology. Ave. 31 e/ 158 and 190, P.O. Box 6162, CP 10600, Havana, Cuba. *Corresponding author.
Received in August 1996. Accepted for publication in November 1996.
Code Number:BA97022
Size of Files:
Text: 37.3K
Graphics: Line drawings (gif) - 2.3K
Photographs (jpg) - 102.7K
ABSTRACT The gene tab3 contains the DNA sequence for 15 aminoacids (aa) of the V3 loop from six different isolates of HIV-1, joined by the sequence AGGGA. This gene was fused to a DNA fragment encoding the first 26 aa of the human interleukin 2 (hu-IL2) and cloned in an expression plasmid. The fusion protein (TAB4) was expressed at high levels in different Escherichia coli strains. The removal of the hu-IL2 fragment from the plasmid abolished the expression of the protein. Similar results were obtained for plasmids pTAB7 and pTAB7SE, containing two copies of tab3 with and without the hu-IL2 stabilising sequence, respectively. Plasmids pVB1 and pVB2, in which the gene tab3 was fused to the bacterial signal peptides OmpA and PelB, were also evaluated. Proteins VB1 and VB2 were expressed at low levels and were not translocated to the E. coli periplasm. Our results indicate that the lack of expression of proteins without hu-IL2 was neither a consequence of the stalling of ribosomes at rare AGA codons, nor a product of a more complex secondary structure around the Shine-Delgarno region or the AUG triplet. The analysis of steady-state levels of specific transcripts showed that differential availability of functional messenger ribonucleic acids determined these differences in the expression of the multi-epitope polypeptides.
Key words: V3 loop, functional mRNA, secondary structure, dna Y, argU gene RESUMEN El gen tab3 contiene la secuencia de ADN para 15 aminoacidos (aa) del lazo V3 de seis aislamientos distintos de VIH-1, unidos por la secuencia AGGGA. Este gen se fusiono en el 5 a un segmento de ADN que codifica los primeros 26 aa de la interleuquina 2 humana (IL2-h). La proteina de fusion (TAB4) se expreso a altos niveles en varias cepas de Escherichia coli; sin embargo, la expresion fue abolida al eliminar la region de IL2-h del plasmidio. Se construyeron plasmidios con dos copias del gen tab3 con y sin el segmento de IL2-h pero los resultados fueron similares. Se obtuvieron los plasmidios pVB1 y pVB2 con el gen tab3 fusionado a las secuecias para los peptidos senales de OmpA y PelB. Las proteinas VB1 y VB2 se expresaron a bajos niveles y no fueron translocadas al periplasma de E. coli. Nuestros resultados indican que la falta de expresion de las proteinas sin la IL2-h no es debido a la aparicion de codones raros en el gen, como AGA para arginina, ni a la formacion de estructuras secundarias en las que esten involucradas la region de Shine-Delgarno o el codon de iniciacion. El analisis de los niveles de los transcriptos especificos sugiere que la disponibilidad de acido ribonucleico mensajero funcional es la causa de estas diferencias en la expresion.
Palabras claves: V3, ARNm funcional, estructura secundaria, dna Y, gen argU Introduction The majority of the scientific community recognizes the human immunodeficiency virus type 1 (HIV-1) as the etiologic agent of AIDS (1). The search for a vaccine against this virus is a great challenge due to the distinctive characteristics of HIV-1. A rational approach in vaccine design is the identification of the viral components with the ability to induce protection. In the case of HIV-1 the external glycoprotein (gp120) of the virus has been considered the central target of the protective immune response. This protein contains the principal neutralizing domain (PND) (2) which is located on the third variable region between the aminoacids (aa) 296-331, at the so called V3 loop. High titers of neutralizing antibodies against this region are induced upon immunizations with gp120, gp160 or synthetic V3 peptides (3, 4).
Several laboratories have developed vaccine strategies based on V3 synthetic peptides (5). The approach we have followed is the use of multi- epitope polypeptides (MEPs) which include peptides with immunological relevance for protection. We have demonstrated the feasibility of employing those MEPs as immunogens to induce HIV-1 neutralizing monoclonal antibodies in mice (6) and neutralizing polyclonal antibodies in rabbits (7).
The design of an efficient expression system is an important aspect in the production of recombinant proteins intended for vaccine applications. We have previously designed a DNA fragment encoding the central region of the V3 loop (15 aa) of gp120 from isolates LR150, JY1, RF, MN, BRVA, and IIIB of HIV-1 (7). The fragment was expressed as a fusion product with the amino terminus of the human interleukin 2 (hu-IL2). This sequence has been successfully used as a stabiliser by other authors (8), although its mechanism of action is not clear. In the present work we studied the role of the IL2 gene fragment at the transcription/translation level. We also explored the use of bacterial signal peptides from outer membrane protein A (OmpA) and pectate lyase B (PelB) (9) as alternative stabilisers. This approach may also allow the production of the MEP in the periplasm of Escherichia coli devoid of foreign N-terminal sequences. Materials and Methods Genetic constructions The gene tab3 (7) is composed of the coding sequences for the central 15 aa of the V3 loop from six different isolates of HIV-1 (LR150, JY1, RF, MN, BRVA and IIIB). The peptides are separated by a sequence of 5 aa (AGGGA) (6).
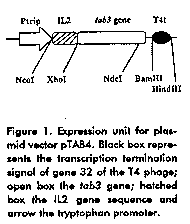
The expression plasmid (pTAB4) (Figure 1) carries a gene encoding a fusion protein of 143 aa, with 26 residues from the amino terminal part of hu-IL2 at the amino terminus. This protein was called TAB4.
We made five genetic constructions derived from pTAB4 (Table 1). For the construction of pTAB7, the plasmid pTAB4 was restricted NdeI-HindIII and the large fragment was purified and ligated to an in-frame synthetic adapter carrying cohesive ends NdeI-XhoI and the DNA fragment XhoI-HindIII from pTAB4. The resulting plasmid had two copies of the gene tab3 fused to the coding sequence for the first 26 aa of the hu-IL2 at the amino terminus. It was named pTAB7. Plasmids pTAB4 and pTAB7 were cleaved with NcoI-XhoI and the ends filled using the Klenow fragment of DNA polimerase I. These preparations were further ligated. The results of these genetic manipulations were two plasmids encoding the proteins TAB4 and TAB7 without the 26 aa of hu-IL2. These variants were named pTAB4SE and pTAB7SE, respectively. Table 1. Plasmids used in the study.
---------------------------------------------------------------------------
Plasmids Description Expected products
---------------------------------------------------------------------------
pTAB4 One copy of the tab3 gene fused TAB4, 16 kDa with the
to the sequence encoding the first N-terminal part of hu-IL2.
26 residues of IL2 at the 5'end,
trp promoter.
pTAB4SE One copy of the tab3 gene, trp TAB4SE, 13 kDa without the N-
promoter. terminal part of hu-IL2.
pTAB7 Two copies of the tab3 gene fused TAB7, 27 kDa with the N-
to the sequence encoding the first terminal part of hu-IL2.
26 residues of IL2 at the 5'end,
trp promoter.
pTAB7SE Two copies of the tab3 gene, trp TAB7SE, 24 kDa without the N-
promoter. terminal part of hu-IL2.
pVB1 One copy of the tab3 gene, fused VB1, 13 kDa in the periplasm
to the sequence encoding OmpA of E. coli
leader at the 5' end, trp promoter.
pVB2 One copy of the tab3 gene, fused VB2, 13 kDa in the periplasm
to the sequence encoding PelB of E. coli
leader at the 5'end, trp promoter.
pACYD6 Kan^R, argU gene in the same tRNA^AGA for arginin
orientation as the bla gene.
---------------------------------------------------------------------------
Plasmids pAS15 and pAS16 (9) are derivatives of pBR322 with the tryptophan promoter and the terminator sequence of T4 phages. They have the complete hu-IL2 gene sequence fused to the sequences encoding bacterial signal peptides from the OmpA and from the PelB, respectively. They were digested with XhoI-BamHI and the large fragments were purified and ligated with the DNA fragment XhoI-BamHI from pTAB4. The resultant plasmids pVB1 and pVB2 correspond to the fusion of gene tab3 to sequences encoding bacterial signal peptides from OmpA nd PelB, respectively.
The plasmid pACYD6 (10) was used in a co-transformation experiment with pTAB4SE. It contains the argU gene in the same orientation as that of the gene that confersampicillin resistance. The argU gene encodes tRNA^AGA for arginin.
All genetic manipulations were carried out following the methods of Sambrook J et al. (11). The restriction and modifying enzymes were obtained from New England Biolabs (MA, USA) and used according to the manufacturer s recommendations. The DNA sequence of the genes in the plasmids was verified by the Sanger method (12) using the Sequenase 2.0 reaction kit (USB, Cleveland, OH, USA). Bacterial strains and growth conditions The E. coli strain MC1061 (F^- araD139 delta (ara-leu)7696 galE15 galK16 delta (lac)X74 rpsl (Str^r) hsdR2 (rK^-mK^+) mcrAmcrB1) was used for plasmid propagation and strains MM294 (F^- endA1 hsdR17 (rK^-mK^+) supE44 thi-1 relA1? rfbD1? spoT1?) and W3110trpA^905 (F^- mcrAmcrB IN (rrnD-rrnE)1 ^- trpA905) were used for expression experiments. W3110trpA905 was used for messenger ribonucleic acid (mRNA) analysis and co-transformation experiments.
Cells harbouring plasmids were grown overnight at 37 C in Luria broth (LB) supplemented with ampicillin (50 ug/mL) and tryptophan (100 ug/mL). Cells transformed with pTAB4, pTAB4SE, pTAB7 and pTAB7SE were transferred from LB overnight cultures to LB medium supplemented with ampicillin (50 ug/mL) with an initial OD 530nm of 0.1. They were grown at 37 C until OD 530nm of 0.8-1.0. The expression of the proteins was induced with beta indolacrilic acid to a final concentration of 20 ug/mL and the growth was continued for 10-12 h. Cells transformed with pVB1 and pVB2 were transferred to M9 medium supplemented with 2 % casein hydrolysate, 0.2 % glucose, ampicillin (50 ug/mL) and tryptophan (10 ug/mL) with an initial OD 530nm of 0.1. The growth and induction conditions were the same, except that incubations were at 28 C (13).
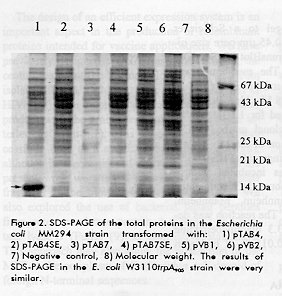
For the analysis of mRNAs from pTAB4, pTAB7, pTAB4SE and pTAB7SE the culture samples were harvested at 4, 8 and 12 h after the addition of the beta indolacrilic acid. Co-transformation experiments The E. coli W3110trpA905 strain was co-transformed with 1 ug of plasmids pTAB4SE and pACYD6. After shock at 42 C the cells were grown in LB medium for 20 min and plated on LB medium supplemented with kanamicine (50 ug/mL) and ampicillin (50 ug/mL). SDS-PAGE and western blot Cell pellets (20 ug of total protein) were suspended in 300 uL of Laemmli sample buffer (14), incubated at 98 C for 5 min and subjected to 15 % SDS- PAGE. For Western blot, the proteins were electroblotted from acrylamide gel to a Schleicher & Schuell (Keene, NH, USA) 0.45 um pore size nitro- cellulose membrane in a TransBlot SD SemiDry (BioRad, California, USA). The membrane was blocked with 5 % non-fat dry milk in PBS (0.14 M NaCl, 0.003 M KCl, 0.008 M Na2HPO4 and 0.0015 M KH2PO4), and treated for 1 h, at 37 §C, with 5 ug/mL of the monoclonal antibody 2C4, which reacts with the V3 region of the MN isolate of HIV-1 (6). After three washings with PBS/0.05 % Tween20, the membrane was incubated with a 1:2000 dilution of an anti-mouse IgG conjugated with HRPO (Boheringer-Mannheim, Mannheim, Germany) for 30 min at 37 C. The reaction was developed with 0.5 mg/mL of 3.3 diaminebenzidine (Sigma, St. Louis, MO) and 0.015 % of hydrogen peroxide. Secondary structure of mRNA The secondary structure analysis of mRNAs by the computer program PCFOLD (15) was restricted to regions of approximately 100 nucleotides (16). Northern blot analysis To ensure that the quantities of total cells were the same in all the samples, the culture volume was harvested following the formula 2/OD 530nm, and the total RNA was extracted (17). Samples (20 % of the total RNA) were subjected to 1.2 % agarose/formaldehyde gel at 100 V and 4 C according to Sambrook et al. (11). For Northern blot, the RNAs were transferred from the gel to a Schleicher & Schuell (Keene, NH, USA) 0.45 um pore size nitro-cellulose membrane by capillarity in SSC (3 M NaCl, 0.3 M sodium citrate, pH 7) overnight at room temperature. The membrane was processed and hybridised as described by Sambrook et al. (11). Results Expression of the proteins in two bacterial strains E. coli W3110trpA905 and MM294 strains were transformed with the plasmids described in Table 1, and the recombinant strains were analysed for their ability to produce the expected proteins. A predominant band of 16 kDa was observed in SDS-PAGE of the sample from pTAB4- transformed cells of the MM294 strain (Figure 2). A similar result was obtained in the W3110trpA905 strain (data not shown). In contrast, it was not detected in samples from either the negative control or the other plasmids.
However, the results of Western blot with anti-V3 monoclonal antibody showed that pTAB7 and pVB2 produced low levels of proteins that were specifically recognised in both strains (Figure 3). These bands were not observed in any of the lanes corresponding to transformed cells with pTAB4SE, pTAB7SE and the negative control. The protein VB1 was only detected when higher quantities of proteins were loaded (data not shown). Proteins VB1 and VB2 showed an electrophoretic migration very similar to TAB4 as verified in Western blot. After an osmotic shock, the proteins remained in the intracellular fraction (data not shown).
Figure 3. Western blot of the proteins in the Escherichia coli strain A)-MM294 and B)-W3110trpA905 transformed with: 1) pTAB4, 2) pTAB4SE, 3) pTAB7, 4) pTAB7SE, 5) pVB1, 6) pVB2, 7) Negative control.
We examined whether the limiting quantities of the tRNA^AGA for arginin in E. coli were influencing the expression of TAB4SE. No positive effect was observed on the expression of TAB4SE when this tRNA is supplied in trans by co-transformation with plasmids pACYD6 and pTAB4SE (Figure 4).
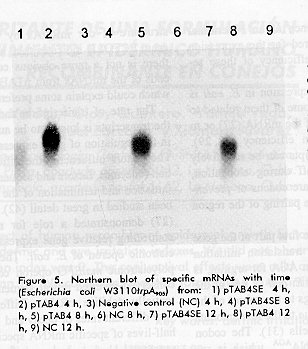
Analysis of the secondary structure of our mRNAs by the computer program PCFOLD did not reveal any base pairing involving the Shine-Delgarno (SD) sequence or the AUG triplet (data not shown) Availability of functional mRNA is the cause of differences in expression The levels of specific mRNAs were analysed for plasmids pTAB4, pTAB4SE, pTAB7 and pTAB7SE in the E. coli W3110trpA905 strain. Total RNA was extracted at 4, 8 and 12 h after the addition of beta indolacrylic acid. Figure 5 shows the results of Northern blot. The levels of specific mRNA from pTAB4SE were lower than the levels from pTAB4. Moreover, the quantities of mRNA from pTAB4SE decreased faster than the mRNA from pTAB4. Similar results were found for the pair pTAB7 / pTAB7SE. This disparity is not due to differences in the plasmid copy number or cell viability on the cultures (results not shown).
We previously described the expression of the HIV-1 MEP TAB4 in E. coli W3110trpA905 (7). Here we constructed several MEP variants and studied their expression in E. coli, as well as the influence of the N-terminal IL2 stabilising sequence on the expression of the proteins.
E. coli strains with different genetic backgrounds were transformed with pTAB4 and the expression of TAB4 was evaluated. Both strains showed high levels of expression. These results indicate that there is no sensitive influence of the genotype of those strains on the expression of TAB4. In another set of experiments, the fragment encoding the N-terminal IL2 stabilising sequence was removed from pTAB4, resulting in the abolishment of the expression, undetectable either by SDS-PAGE or Western blot. We observed this effect with the MEP TAB1 (6) and with other proteins (unpublished), but the reasons are unknown. It has been reported that some recombinant proteins are rapidly degraded when expressed intracelullarly in E. coli and that this problem could be particularly marked with proteins of low molecular weight (18). One of the most popular solutions to it, is the construction of fused genes which somehow enhance the intracellular stability of the protein products (19, 20). To investigate if the loss of expression of the pTAB4SE was related to the size of the protein, we evaluated the expression of the plasmids pTAB7 having two copies of the gene tab3 and the IL2 stabilising sequence and of the pTAB7SE, without the stabilising sequence. It was possible to obtain a certain expression from the pTAB7 but not from pTAB7SE. This result suggests that, whatever the reason for the lack of expression of the series of SE plasmids, it is not directly related to the size of the protein. This, together with the absence of bands of degradation products in any of Western blots, makes the degradation of the proteins a very unlikely explanation for the lack of expression.
Two additional N-terminal regions from bacterial protein were tested. Plasmids pVB1 and pVB2 carried the gene tab3 fused to the sequences encoding signal peptides from OmpA and PelB, respectively. In addition, this strategy would allow the recovery of the products in the periplasm of E. coli without any stabilising sequence at the amino terminus of the protein in case they were properly processed.
The expression of pVB1 and pVB2 was detectable only by Western blot, indicating the presence of very low amounts of both proteins. Furthermore, our results indicate that VB1 and VB2 were not translocated across the cytoplasmic membrane, in spite of the recognised ability of those signal peptides to direct the efficient translocation of eukaryotic and prokaryotic proteins (13, 21).
The choice of prokaryotic signal sequences should not be critical for the translocation process, since signal peptides from OmpA, PelB, alkaline phosphatase, beta lactamase and OmpF have been successfully used in several expression systems (9, 22, 23). However, depending on the protein sequence, some negative influence could be expected. Several authors have pointed out that the presence of positively charged aminoacids within the 20-30 residues downstream of the signal peptide can inhibit the protein translocation to the periplasm (24-26). They demonstrated (24) that a single point mutation immediately after the signal sequence, that changes any aminoacid to arginin, is enough to block the translocation of polypeptides. The insertion of six consecutive lysines among the residues 14-21 downstream from the signal sequence could also block the translocation.
The gene tab3 encodes a protein that has an arginin as third aminoacid, and three more within the first 25 residues of the polypeptide (positions 6, 10 and 21). The blocking of the translocation to the periplasm could probably be a consequence of the presence of positively charged aminoacids just downstream of the signal peptide.
The fact that the fusion of tab3 to OmpA and PelB allows the expression of the protein, is in agreement with the expression from pTAB4 and contrasts with the results from pTAB4SE. This demonstrated that the presence of an N-terminal stabilising sequence is required for the expression of this MEP, although the efficiency of these sequences could be different.
It is known that protein expression in E. coli is influenced by many factors, some of them related to the processing and stability of the mRNA (27) or to differences in the translation efficiency (28, 29). The translation of the transcripts can be negatively affected by ribosome drop- off during elongation, stalling of the ribosomes at rare codons or prevention of ribosome entry, by base pairing of the region spanning the SD sequence.
Stalling at rare codons in the first part of the gene has been suggested to affect translation initiation and elongation (30). The codon AGA for arginin, is one of the least used codons in E. coli (1.7 % of the total codon content) and belongs to the subset of the most slowly translated ones, due to the low amounts of its cognate tRNA in the cells (31). The codon AGA is recognised by the tRNA^AGA, which is encoded in E. coli by the argU gene (32). Some authors have found that recombinant gene expression in E. coli can be severely affected by the limitation of minor tRNA^AGA if its AGA/AGG content is higher than 3 % (33). Varenne S et al. (34) established that consecutiveness rather than the number of rare arginin codons is critical for the protein expression. Others have postulated the "Minor Codon Modulator Hypothesis", which correlates the presence of AGA or AGG codons in the first 25 aa of the proteins with a poor expression of those proteins. These two codons are able to limit the expression levels either by their position in the polypeptide or by their abundance (35, 36). Brinkman U et al. (33), Garcia OL et al. (10) and Schenk PM et al. (37) used the overexpression of the argU gene in trans as a successful strategy to increase the expression of some proteins with a high content of AGA/AGG (human alpha-2 interferon, tPA, uPA, alpha-Gluc, alpha-Gal, 28 kDa protein of barley yellow mosaic virus and HIV-1gp41). In the present study, the W3110trpA^905 strain was co-transformed with plasmids pACYD6 (10) and pTAB4SE in order to learn if the AGA codon in position 3 in this protein was somehow influencing the expression. We did not detect any expression in cells harbouring both plasmids either by SDS-PAGE or Western blot, suggesting that this rare codon is not limiting the expression of the genes without the stabilising sequence.
Different authors have reported the influence of the mRNA secondary structure in the protein expression (38-40). The direct engagement of the initiating AUG codon and/or the SD sequence in any hydrogen bonded structure could affect the efficiency of translation. Analysis of the secondary structure of our mRNAs by the computer program PCFOLD indicates that, at least theoretically, there is not a more obvious complexity at the 5' ends of the mRNAs from pTAB4SE and pTAB7SE which could explain some preferential interferences.
The rate of transcription and/or the stability of the transcripts is known to be an important element in the regulation of gene expression in E. coli (41). The factors influencing the efficiency of transcription (enzymes, factors and sequences involved in the initiation and termination of the transcription) have been studied in great detail (42). Newbury SF et al. (27) demonstrated a role for mRNA stability in controlling relative gene expression within a polycistronic operon of E. coli. They showed that the polycistronic malEFG operon contains two highly conserved inverted repeat sequences that could potentially stabilise mRNAs. Different mRNAs have very different rates of decay and in many cases, the half-lives of specific mRNA species vary with environmental changes in a manner that is consistent with a role for mRNA stability in the control of gene expression (43). Current understanding suggests that mRNAs are generally stabilised by structures at the termini, and that the high susceptibility to decay of individual species may be determined by special features that provide targets for specific enzymes (44).
Analysis of the steady-state levels of specific transcripts was done for some of the plasmids. The results showed that there was a marked difference between them. The fact that the serie of SE plasmids have less specific mRNA and that its relative amounts decrease faster suggests either that they are produced at a slower rate or that they are more rapidly degraded. Although the amounts of SE-mRNAs observed in the Northern blot could seem to be sufficient to support some protein expression, there is no guaranty that these mRNAs are functionally active. It is established that the functional inactivation of mRNA could be observed with very little structural degradation (for instance, the removal of the 5' end translation signals) and it is not dependent on total chemical instability (42).
In summary, we strongly support the idea that the differential availability of functional mRNA determines the differences of expression from the plasmids studied. Whether these differences are supported by a higher rate of transcription favoured by the presence of the stabiliser at the 5' end or by a higher functional and /or chemical stability, needs to be determined. Acknowledgements We thank Jesus Seoane for the pictures and Ricardo Bringas for the analysis of the secondary structure of the mRNA molecules. References 1. Barre-Sinoussi F, Chermann JC, Rey F, Nuguyere MT, Chamaret S, Gruest J et al. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science 1983;220:868- 871. 2. Javaherian K, Langlois AJ, McDanal C, Ross KL, Eckler LI, Jellis CL et al. Principal neutralizing domain of the human immunodeficiency virus type 1 envelope glycoprotein. Proc Nat Acad Sci USA 1989;86:6768- 6772. 3. Putney SD, LaRosa GJ, Profy AT, Silver S, Scott CF, Javaherian K et al. HIV-1 principal neutralizing determinant elicits broadly neutralizing antibodies. In: Vaccine 91. 1991. Eds. Brown F, Chanock RM and Lerner RA. Cold Spring Harbor Laboratory. 4. Javaherian K, Langlois AJ, LaRosa GJ, Profy AT, Bolognesi DP, Herlihy WC et al. Broadly neutralizing antibodies elicited by the hypervariable neutralizing determinant of HIV-1. Science 1990;250:1590- 1593. 5. Hart MK, Palker TJ, Matthews TJ, Langlois AJ, Lerche NW, Martin ME et al. Synthetic peptides containing T and B cell epitopes from HIV envelope gp120 induce anti-HIV proliferative responses and high titers of neutralizing antibodies in Rhesus monkeys. The Journal of Immunology 1990;145:2677-2685. 6. Duarte CA, Montero M, Seralena A, Valdez R, Jimenez V, Benitez J et al. Multiepitope polypeptide of HIV-1 envelope induces neutralizing monoclonal antibodies against the V3 loop. AIDS Res Hum Retrov 1994;10:235- 243. 7. Montero M, Duarte C, Menendez A, Navea L et al. Immunogenicity of an HIV-1 multiepitope peptide (MEP). X International Conference on AIDS, Yokohama. 1994. Abstract No. PA:0371. 8. Novoa LI, Madrazo J, Fernandez JR, Benitez J, Narciandi E, Rodriguez JC et al. Method for the expression of heterologous proteins produced in fused form in Escherichia coli, use thereof, expression vectors and recombinant strains. European Patent Application 416673 A1. 1991. Bulletin 91/11. 9. Ayala M, Duenas M, Santos A, Vazquez J, Menendez A, Silva A et al. Bacterial single chain antibody fragments, specific for carcinoembryonic antigen. BioTechniques 1992;13:790-799. 10. Garcia OL, Gonzalez B, Menendez A, Sosa AE, Fernandez JR, Santana H et al. The argU gene product enhances expression of the recombinant human alpha2-interferon in Escherichia coli. Annals of New York Acad Sci 1996;782:79-86. 11. Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning. A Laboratory Manual. 1989. Sec. Ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. 12. Sanger F, Nicklen S, Coulson AR. DNA sequencing with chain-terminating inhibitors. Proc Nat Acad Sci USA 1977;74:5463-5467. 13. Duenas M, Vazquez J, Ayala M, S”derlind E, Ohlin M, Perez L et al. Intra and extracellular expression of a scFv antibody fragment in Escherichia coli: Effect of bacterial strain and pathway engeneering using GroES/L chaperonins. BioTechniques 1994;16:476-483. 14. Laemmli NY. Cleavage of structural proteins during the assembly of the head of the bacteriophage T4. Nature 1970;227:680-685. 15. Zucker M, Stiegler P. Optimal computer folding of large mRNA sequences using thermodynamic and auxiliary information. Nucl Acids Res 1981;9:133- 149. 16. Schauder B, McCarthy JEG. The role of bases upstream the Shine-Delgarno region and in the coding sequence in the control of gene expression in Escherichia coli: translation and stability of mRNAs in vivo. Gene 1989;78:59-72. 17. Miller JH. Experiment in Molecular Genetics. 1972. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. 18. Georgiou G. Optimizing the production of recombinant proteins in microorganisms. AlChE Journal 1988; 34: 1233-1248. 19. Lin Y, Liu Y-X, Cisio T, Mason LB, Yu M-YW. Expression and characterization of the preS1 peptide of hepatitis B surface antigen in Escherichia coli. J Med Virol 1991;33:181-187. 20. Shen S-H. Multiple joined genes prevent product degradation in Escherichia coli. 1984. Proc Nat Acad Sci USA 1984;81:4627-4631. 21. Ayala M, Balin RF, Fernandez de Cossio ME, Canaan-Haden L, Larrick JW, Gavilondo JV. Variable region sequence modulates periplasmic export of a single-chain Fv antibody fragment in Escherichia coli. BioTechniques 1995;18 (5).
22. Batra JK, Chaudhary VK, Fitzgerald D, Patsan I. TGF alpha-antitac (Fv)- PE40: A functional toxin cytotoxic for cells with EGF or IL2 receptors. Biochem Biophys Res Commun 1990;171:1-5. 23. Buchner J, Rudolf R. Renaturation, purification, and characterization of recombinant Fab-fragment produced in Escherichia coli. Bio/Technology 1991; 9: 157-162. 24. Johansson M. Positively charged aminoacids placed next to a signal sequence block protein translocation more efficiently in Escherichia coli than in mammalian microsomes. Mol Gen Genet 1993;239:251-256. 25. Yamane K, Misushima S. Introduction of basic amino acid residues after the signal peptide inhibits protein translocation across the citoplasmic membrane of Escherichia coli. J Biol Chem 1988;263:19690-19696. 26. Boyd D, Beckwith J. The role of charged aminoacids in the localization of secreted and membrane proteins. Cell 1990;62:1031-1033. 27. Newbury SF, Smith NH, Higgins CF. Differential mRNA stability control relative gene expression within a polycistronic operon. Cell 1987;51:1131- 1143. 28. McCarthy JE, Bokelman C. Determinants of translational initiation efficiency in the atp operon of E. coli. Mol Microbiol 1988;2:455-465. 29. Sampson LL, Hendrix RW, Huang WM, Casjens SR. Translational initiation controls to the relative rates of the expression of the bacteriophage late genes. Proc Nat Acad Sci USA 1988; 85: 5439-5443. 30. Bergmann JE, Lodish HR. A kinetic model of protein synthesis. Application to hemoglobin synthesis and translation control. J Biol Chem 1979;254:11927-11937. 31. Bonekamp F, Jensen KF. The AGG codon is translated slowly in E. coli even at very low expression levels. Nucl Acids Res 1988;16:3013- 3024. 32. Garcia GM, Mar PK, Mullin DA, Walker JR, Prather NE. The Escherichia coli dnaY encodes an arginin transfer RNA. Cell 1985;45:453-459. 33. Brinkman U, Mattes RE, Buckel P. High level expression of recombinant genes in Escherichia coli is dependent on the availability of the dna Y gene product Gene 1989;85:109-114. 34. Varennes S, Baty D, Verheij H, Shire D, Lazdunski C. The maximum rate of gene expression is dependent on the downstream context of unfavourable codons. Biochimie 1989;71:1222-1229. 35. Inouye M, Chen G. Regulation of gene expression by minor codon in E. coli; minor codon modulator hypothesis. In: NATO ASI Series. Post transcriptional control of gene expression. 1990. Eds. McCarthy JEG and Tuite MF. Springer Verlag, H49. 36. Chen GT, Inouye M. Suppression of the negative effect of minor arginin codons on gene expression; preferential usage of minor codons within the first 25 codons of the Escherichia coli genes. Nucleic Acids Res 1990;18:1465-1473. 37. Shenk PM, Baumann S, Mattes R, Steinbibeta HH. Improved high-level expression system for eukaryotic genes in Escherichia coli using T7 RNA Polymerase and rare ^ArgtRNAs. Biotechniques 1995;19:196-200. 38. Iserentant D, Fiers W. Secondary structure of mRNA and the efficiency of translation initiation. Gene 1980;9:1-12. 39. Kubo M, Imanaka T. mRNA secondary structure in an open reading frame reduces translation efficiency in Bacillus subtilis. J Bacteriol 1989;171:4080-4082.
40. Wikstrom PM, Lind LK, Berg DE, Bjork GR. Importance of mRNA folding and start codon accessibility in the expression of genes in a ribosomal protein operon of Escherichia coli. J Mol Biol 1992;224:949-966. 41. Higgins CF, Smith NH. Messenger mRNA processing, degradation and the control of gene expression. In: Regulation of gene expression, S.G.M. 1986; Symposium Vol 39: 179-198. Eds. Higgins CF and Booth IR. Cambridge University Press. Cambridge. 42. Lewin B. Gene IV. 1990. Cell Press, Cambridge, Mass. Oxford University Press, Walton Steet, Oxford OX2 6DP. 43. vonGabain A, Belasco JG, Schottel JL, Chang ACY, Cohen SN. Decay of mRNA in E. coli: investigation of the fate of specific segments of transcripts. Proc Natl Acad Sci USA 1983;80:653-657. 44. Brawerman G. Mechanisms of mRNA decay. TIBTECH 1990;8:171-174. Copyright 1997 Elfos Scientiae The following images related to this document are available:Photo images[ba97022d.jpg] [ba97022c.jpg] [ba97022e.jpg] [ba97022b.jpg]Line drawing images[ba97022a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}