 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Genetic characterization of HIV-1 strains isolated from Cuban seropositive patients Carmen E Gomez,^1 Enrique Iglesias,^1 Julio R Fernandez,^1 Felipe M Rolo,^2 Leonor Lobaina,^2 Madelin Blanco,^2 Enrique Noa,^2 Antonieta M Herrera^1 and Carlos A Duarte^1
^1 Division de Vacunas, Centro de Ingenieria Genetica y Biotecnologia,
apartado postal 6162, Ciudad Habana 10600, Cuba.
Code Number:BA97028
Size of Files:
Text: 14.2K
Graphics: Line drawings (gif) - 8.6K
Introduction Soon after the discovery of Human Immunodeficiency Virus type 1 (HIV-1) as the etiologic agent of AIDS, the significant genetic heterogeneity of this virus evoked intense interest. Many studies have shown the existence of inter and intrapatient variability (1) and that mutations are not evenly distributed throughout the genome (2). Variation in the env gene is likely to be the consequence of the selective pressures from either B or T cell responses, and it yields virus strains that escape the immune system surveillance. The analysis of the major structural genes of the virus, including gag and env, has highlighted the existence of multiple HIV-1 genetic subtypes (3). Some of them appears to be widely distributed geographically, and several regions harbour a mixture of subtypes. HIV-1 vaccine development faces many formidable challenges, one of them is the variability of the viral genome. Given the extreme degree of genetic divergence, the immunogens based on a single viral subtype may not elicit an immune response broad enough to be protective against the entire spectrum of viruses that exist world-wide, or even within a single geographic region. Several groups, including ours, have based their vaccine strategies in the V3 loop of gp120. This region contains the Principal Neutralising Domain of HIV-1 and is known to mediate the fusion of the viral and cell membranes (4). A survey of the V3 variability in the HIV-1 isolates circulating in the region is therefore very important. This paper describes the genetic heterogeneity of the V3 region of HIV-1 gp120 from 12 Cuban HIV-1 infected patients. Materials and Methods
Collection and preparation of DNA
Whole blood samples were obtained from 12 HIV-1 seropositive subjects infected mainly in Cuba. The clinical features of these patients are illustrated in Table 1. Peripheral blood mononuclear cells (PBMC) were isolated by ficoll-hypaque density gradient centrifugation of a heparinized whole blood specimen, and DNA was prepared through non-ionic detergent lysis and proteinase K digestion followed by organic extraction method from 10^6 PBMCs. Table 1. Clinical features of HIV-1 infected individuals. --------------------------------------------------------------------------- Patient Sequence Sex Transmission CDC Sexual number code route classification orientation --------------------------------------------------------------------------- 216 93CU051 M Sexual IV C2 HB 371 93CUPL16 M IVDU IV C2 HB 421 93CUPL09 M Sexual IV C1 HB 434 93CUPL08 F Sexual IV C1 HT 465 93CUPL11 M Sexual IV C1 HB 480 93CUPL18 M Sexual II HB 519 93CUUSPL42 M Sexual II HB 632 93CUPL10 M Sexual IV C1 HB 666 93CU044 M Sexual II HB 711 93CUPL05 F Sexual IV C2 HT 757 93CUUSPL01 M Sexual IV C1 HB 909 93CU053 M IVDU IV C1 HB ---------------------------------------------------------------------------CDC: Centre for Disease Control and Prevention, IVDU: intravenous drug user, HB: heterobisexual, HT: heterosexual. Virus isolation Virus isolation was attempted by co-cultivation of patient peripheral blood mononuclear cells (PBMCs) with seronegative donor PBMCs previously stimulated with phytohemaglutinin A (PHA) for three days. Cultures were supplemented with fresh stimulated donor cells every 3-4 days and monitored for the production of p24 antigen using a commercially available ELISA (DAVIH-p24, DAVIHLAB, La Habana, Cuba). Culture supernatants positive for p24 were harvested and DNA was purified by RNase A and proteinase K digestion, phenol extracted and quantified by optical density at 260 nm. Polymerase chain reaction Nested polymerase chain reaction (PCR) was used to amplify 0.36 Kb of the C2-V3 region from viral DNA. The first round used the following flanking primers:
5'GCGCCCATAGTGCTTCCTGCTGCTCCCAAG 3' - Reverse: 5'TCCTATGGCAGGAAGAAGCGGAGACA 3'
5'ATGAATTCTGGGTCCCCTCCTGAGGA 3'
- Reverse: 5'ATAAGCTTGCAGTCTAGCAGAAGAAGA 3'
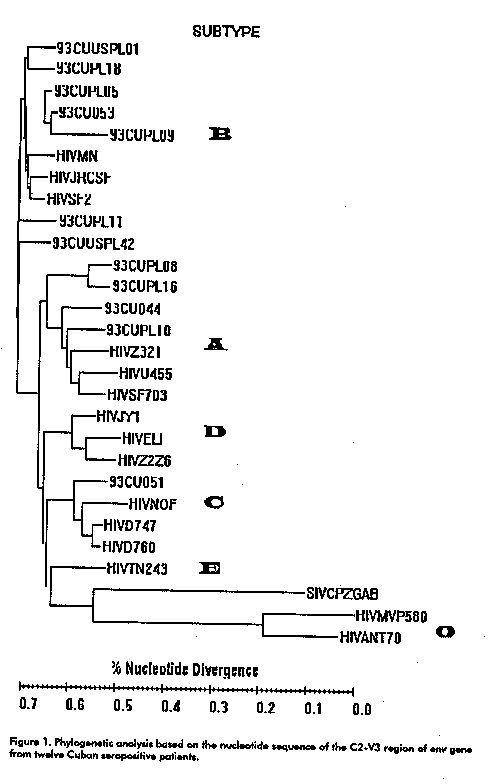
The amplified C2V3 products were cloned in a T-vector (Amersham). Plasmids were purified from positive clones and the DNA sequence determined with Sequenase 2.0 kit (Amersham). The DNA sequence of each of the five V3 clones was subjected to sequential pairwise alignment. Results The sequence of 360 nucleotides from the C2-V3 region was determined for five clones from each individual, and used to generate a consensus. These consensus sequences were subjected to sequential pairwise alignment versus prototype sequences representing the six env subtypes (A-F) within the major HIV-1 cluster (group M) and the group O. Figure 1 shows a phylogenetic tree based on the nucleotide sequences of the C2-V3 region of env including the 12 specimens analysed in this study.
The majority of the viral isolates (7/12) were classified as subtype B. The intrapatient variability ranged between 0 and 10.8 % for DNA, and from 0 to 18.6 % at the amino acid level. Three samples had the typical GPGR motif at the crown of the V3 loop, as in most subtype B sequences, while the others had the unusual GWGR, TPGR and GRGR motifs. The former of these three tetrapeptides was prevalent mainly in Brazilian isolates (5) and the later was reported by us for the first time in a Cuban isolate (6). The V3 consensus from isolates with the GRGR motif displayed several differences with regard to the sequences of the B subtype, specially within the principal neutralising domain (PND). The highly positive charge associated with the loop (+ 5) correlated with the rapid-high, syncytium-inducing (SI) phenotype exhibited by these isolates (Lobaina et al, manuscript in preparation) as has been previously reported (7).
Two samples (93CU044 and 93CUPL10) clustered with subtype A. The intrapatient diversity in these samples ranged between 0 and 5.7 % while the amino acid variability ranged between 0 and 14.3 %. The sequence 93CU051 belonged to subtype C. It lacks the potential N-linked glycosylation site immediately preceding the V3 loop, a typical feature of this subtype.
Finally the other two isolates (93CUPL08 and 93CUPL16) could not be subtyped because they exhibited more than 20 % DNA divergence with all prototype sequences. They were more related to subtype B (26.8 % of divergence) and also lack the potential N-linked glycosylation site immediately preceding the V3 loop. The intrapatient divergence in these samples ranged between 0 and 4.6 % while the tetrapeptide in the crown of the V3 loop were APGQ and GPGQ.
Table 2 shows the intra and interpatient variability, the subtype and the octamer found at the tip of the loop for each sample analysed in the study. In all cases transitions accounted for 70 % of the nucleotides changes and more than 60 % of them were substitutions of G for A. Table 2. Geetic divergence and remarkable features exhibited by the Cuban isolates.
---------------------------------------------------------------------------
Sequence Intra-patient Intra-patient Divergence Subtype Octamer
code divergence (DNA) divergence (aa) with prototype
sequences (DNA)
---------------------------------------------------------------------------
93CU051 0.9 - 4.8 % 2.8 - 9.6 % 17.1 % C KIGPGQAF
93CUPL16 0.6 - 10.8 % 0 - 3.9 % 26.8 % NT HIGPGQAF
93CUPL09 2.4 - 10.8 % 4.6 - 18.6 % 14.4 % B HLGRGRSV
93CUPL08 0 - 2.4 % 0 - 4.6 % 26.8 % NT HIAPGQAL
93CUPL11 0.3 - 4.9 % 0 - 8.4 % 17 % B HIGPGRAF
93CUPL18 0 - 5.6 % 0 - 8.2 % 10.2 % B NIGPGRAF
93CUUSPL42 0 - 9.8 % 0 - 17.3 % 17.9 % B HITPGRAF
93CUPL10 1.5 - 5.7 % 4.6 - 14.3 % 15.8 % A HIGPGQVF
93CU044 0 - 0.5 % 0 - 0.9 % 14.8 % A RIGPGQAF
93CUPL05 1.2 - 4.9 % 2.8 - 9.9 % 7.7 % B HLGWGRSV
93CUUSPL01 1.5 - 9.3 % 3.7 - 13.2 % 10.2 % B PIGPGRAF
93CU053 0 - 2.8 % 1.1 - 5.2 % 4.8 % B YLGRGRSV
---------------------------------------------------------------------------
Discussion
Seven out of 12 isolates from Cuban patients were classified as subtype B confirming the predominance of this subtype in the Caribbean. Subtype B strains are also prevalent in South America, North America and western Europe, as well as in Australia, New Zealand and Japan. However, the other subjects were identified as subtype A and C, mainly prevalent in Africa. These data correlate with the epidemiological characteristics of the HIV-1 infection in Cuba, where the virus has been mainly introduced from America and Africa.
The atypical amino acid motifs detected in this study in the V3 loop can lead to important changes in the antigenicity of the viruses. It has been demonstrated that several monoclonal antibodies against the V3 region lost their recognition capacity when amino acid changes are introduced in the PND (4). In our group we have been studying the reactivity of human serum against peptides including these motifs. In every case, these peptides have been poorly recognised and most of them have reacted exclusively with the homologous serum (Gomez et al, unpublished data).
The most frequent changes occurring during replication are G to A hypermutations, where 15 to 80 % of Gs are changed to As over a stretch of sequence. Such errors were first detected in the chicken retrovirus, Spleen Necrosis Virus and subsequently with HIV-1 and SIV. The number of cases included in this study are small and not representative of the HIV quasispecies circulating in Cuba. This work is part of a study which includes the analysis of a higher number of isolates (Rolo et al, unpublished data). However, further studies should be implemented with randomly selected samples that could be more representative of the HIV-1 epidemic in Cuba. These and others studies will aid the design of appropriate vaccines to slow the spread of HIV-1. References 1. Simmonds P, Bulfe P, Judlom CA, Bishop IO, Brown AJ. Analysis of sequence diversity in hypervariable regions of the external glycoprotein of human immunodeficiency virus type 1. J Virol 1990;64:5840-5850. 2. Pedroza Martins L, Chenciner N, Wain Hobson S. Complex intrapatient sequence variation in the V1 and V2 hypervariable regions of the HIV-1 gp120 envelope sequence. Virology 1992;191:837-845. 3. Myers G, Korber BT, Smith RF, Wain HS, Pavlaxis GN. Human Retroviruses and AIDS. 1993. 4. Duarte CA.The third variable region of the external envelope glycoprotein of the human immunodeficiency virus type 1. Structure, biological properties and immunogenicity. Biotecnologia Aplicada 1997;14(1):1-22. 5. Potts K, Kalish ML, Loot T, Orloff G, Luo CC, Bernard MA et al. Genetic heterogeneity of the V3 region of the HIV-1 envelope glycoprotein in Brazil. AIDS 1993;7:1191-1197. 6. Gomez CE, Fernandez JR, Iglesias E, Lopez AE, Lobaina L, Noa E et al. Complete DNA sequence of the gene encoding the external glycoprotein (gp120) from a Cuban HIV type 1 isolate. AIDS Research and Human Retroviruses 1996;12:553-555. 7. Fouchier RAM, Groenink M, Kootstra NA, Tersmette M, Huisman HG, Miedema F, Schuitemaker H. Phenotype associated sequence variation in the third variable domain of the HIV type 1 gp120 molecule. J Virol 1992; 66(5):3183- 3187. Copyright 1997 Elfos Scientiae The following images related to this document are available:Line drawing images[ba97028a.gif] |
| |||||||||

{kind=link}