 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Glycosylation pattern characterization of natural and recombinant glycoproteins using a two-dimensional mapping technique Omar Quintero, Raquel Montesino, Rossana Garcia, Marlen Mata and Jose A Cremata GlycoLab, Bio-Industry Division, Center for Genetic Engineering and Biotechnology, P.O. Box 6162, Havana, Cuba.
Code Number:BA97034
Size of Files:
Text: 11.6K
Graphics: Line drawings (gif) - 15.7K
Photographs (jpg) - 40.6k
Introduction Several glycan profiling methods have been recently published. They are based on measurement of the oligosaccharide effective size on gel filtration chromatography (1); High Performance Anion Exchange Chromatography retention times and electrophoretic mobilities on High Performance Capillary Electrophoresis (2, 3); and 2-3 dimensional (2D-3D) carbohydrate mapping database of pyridylaminated-oligosaccharide derivatives using C18 and amide-silica HPLC and an extra anion exchange column, respectively (4, 5).
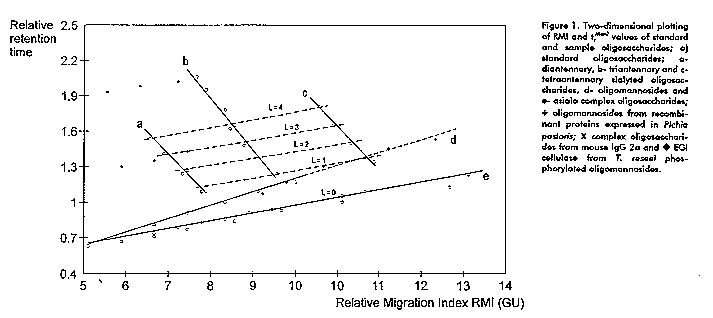
The methodology that will be described is based on the separation of derivatized oligosaccharides on Fluorophore Assisted Carbohydrate Electrophoresis (FACE) (6) and Amine Adsorption-HPLC (7). The first report about the suitability of separation of ANTS (8-amine-1,3,6-naphthalene trisulfonic acid) oligosaccharide derivatives using NH2-HPLC under ion suppression conditions is advanced. We propose a two-dimensional (2D) sugar-mapping technique for (ANTS-derivatives) of neutral and sialyl oligosaccharides as a simple and sensitive technique for structural characterization of N-linked oligosaccharides from natural and recombinant glycoproteins using only picomoles of samples. In addition, the contribution of each monosaccharide residue was determined which facilitate the understanding of the behavior of asialo and sialo complex oligosaccharides. The proposed methodology includes: i) reductive amination with ANTS of enzymaticaly released oligosaccharides, ii) simultaneous separation of derivatized oligosaccharides by FACE and NH2-HPLC column under ion suppression conditions, iii) plotting of the relative migration indexes (RMI with respect to a mixture of malto-oligosaccharides of different degree of polymerization) (X-axis) and relative retention times (tr^Man7 relative to Man7GlcNAc2 oligosaccharide) (Y-axis) in a two coordinates graphic. This methodology fulfill almost all the requirements for a complete characterization of neutral and charged oligosaccharides released from N-glycosylated glycoprotein as it is demonstrated with several examples of natural and DNA-recombinant glycoproteins. Results and Discussion The analysis of oligomannosides and complex oligosaccharides using FACE and NH2-HPLC gave structural complementary information. Individual monosaccharide contributions to RMI and tr^Man7 were determined. Oligomannoside series shows a constant increment per mannose residue added of 0.7 GU and 0.09 on RMI and tr^Man7 respectively. The deletion of a Gal residue in asialo complex oligosaccharide decreases the RMI and tr^Man7 in 1 GU and 0.08 respectively. Furthermore, a larger effect was observed when asialo and sialo complex oligosaccharides are compared. The introduction of a sialic acid determines faster migration on FACE while the same sialic acid produces a remarkable increase on HPLC retention time. Changes on sialic acid linkage configuration are also detected on the RMI and tr^Man7. A two-dimensional plotting of RMI vs tr^Man7
More than 40 different standard oligosaccharides were studied in terms of electrophoresis and HPLC behavior. All these compounds were plotted into a 2D graphic of RMI referred as GU and retention time relative to Man7 (tr^Man7). Figure 1 shows a family of straight lines corresponding to oligomannosides, asialo and sialylated di, tri and tetraantennas. Incorporation of a new member of the oligomannosides or asialo complex series will meet their own curves due to the constant contribution of the corresponding monosaccharides. Analysis of sialo oligosaccharides, showed that, to each antennary structure corresponds one line e.g. the di, tri or tetraantennary lines are composed by the elements of the same structural motif but varying on the number and configuration linkages of the attached sialic acids. All this elements determines straight lines with negative slopes. Since the addition of a Fuc unit to the inner core is depicted by a positive shift of the corresponding point, then, the fucosylated isomers generate a new set of points and thus a new line parallel to the unfucosylated structures. As well as Fuc addition generates a new line, some other factors does, e.g. the deletion of galactose or N-Acetyl glucosamine units and the introduction of lactosamine extensions (data not shown).
Apart from the already described straight lines another set of parallel lines with positive slopes can be detected. All these ones are characterized by a parameter "L" which is a function of the "effective charge" disclosed by the molecule. Thus, this parameter "L" is, among other factors, dependent of the number of functional carboxylic groups and on the configuration linkages of the sialic acid at the non-reducing end of the sialo oligosaccharide. Each new line is constituted by the RMI and tr^Man7 values of oligosaccharides which display the same "effective charge" determined by the equation:
L = 2 x # Sialic Acid (alpha 2,6 linked) +1 x # Sialic Acid (alpha 2,3 linked)
N-linked glycan profiling of glycoproteins
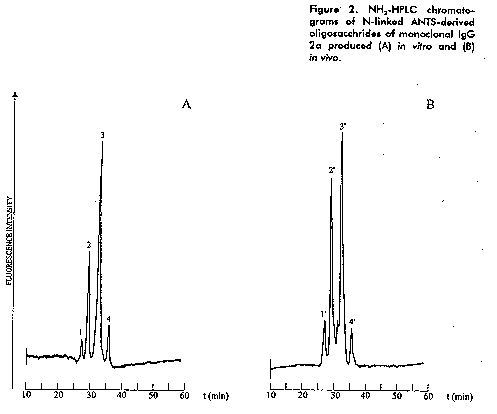
IgG 2a produced in vivo and in vitro gave a typical N-linked glycosylation profile of the IgG family showing small asialo oligosaccharides as has been reported before. The major structures corresponds to the monogalacto-core fucosylated dianntena (in two positional isomers) followed by the agalacto - core fucosylated diantenna in proximal ratios of 0.5:1 and 0.8:1. Other two minor species were detected, the digalacto - core fucosylated and the unfucosylated agalacto dianntenas (Figure 2-4).
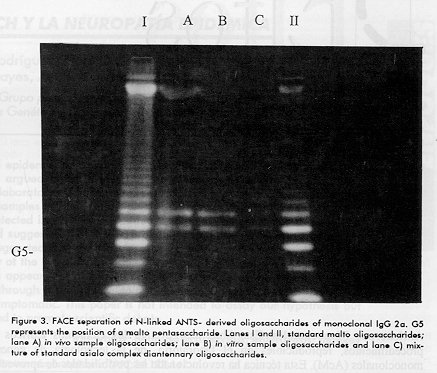
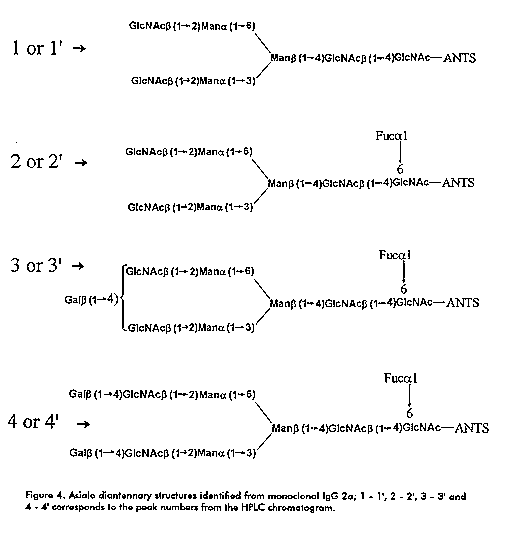
Figure 3. FACE separation of N-linked ANTS- derived oligosaccharides of monoclonal IgG 2a. G5 represents the position of a malto pentasaccharide. Lanes I and II, standard malto oligosaccharides; lane A) in vivo sample oligosaccharides; lane B) in vitro sample oligosaccharides and lane C) mixture of standard asialo complex diantennary oligosaccharides. Figure 4. Asialo diantennary structures identified from monoclonal IgG 2a; 1 - 1', 2 - 2', 3 - 3' and 4 - 4' corresponds to the peak numbers from the HPLC chromatograms.
Other examples were also studied and the corresponding pair of values of each oligosaccharide were plotted in the 2D graphic (Figure 1). These include the glycosylation profiles of several heterologous proteins expressed in the methylotrophic yeast Pichia pastoris, where Man9GlcNAc2 was the common oligosaccharide. An structural modification of this oligosaccharides was possible to be characterized in conjunction with exoglycosidase digestion. Fungal proteins were also analyzed as was the case of Trichoderma reseei endoglucanase 1 "glycoforms". The presence of charged species was considered due to low RMI values and strong retention in HPLC. When plotting the RMI and tr^Man7 in the 2D graphic, the corresponding points lay on parallel lines to the oligomannosides standards and proximal to L = 3 and L = 7 lines of sialylated oligosaccharides. This behavior suggested phosphorylation of oligomannosides oligosaccharides. This was further confirmed by ESI-MS in combination with mild acid hydrolysis and alkaline phosphatase digestion. Conclusions
- The existence of a complex network which in conjunction with the defined contribution of each monosaccharide gives also a predictive character to this web. - Linear dependence within each group of compounds (asialo, mono, di, tri or tetrasialylated oligosaccharides), greatly facilitate the structural characterization of an isolated oligosaccharide. - The developed methodology serve to analyze N-linked glycosylation pattern of natural and recombinant glycoproteins.
1. Kobata A, Yamashita K, Takasaki S. BioGel P-4 column chromatography of oligosaccharides: effective size of oligosaccharides expressed in glucose units. In: Methods in Enzymology. Academic Press, San Diego 1987;138:84- 93. 2. Hermentin P, Witzel R, Vliegenthart JFG, Kamerling JP, Nimtz M, Conradt HS. A strategy for the mapping of N-glycans by high-pH anion-exchange chromatography with pulsed amperometric detection. Anal Biochem 1992;203:281-289. 3. Hermentin P, Doenges R, Witzel R, Kokke CH, Vliegenthart JFG, Kamerling JP, Conradt HS, Nimtz M, Brazel D. A strategy for mapping of N-Glycans by High-performance capillary electrophoresis. Anal Biochem 1994;221:29-41. 4. Tomiya N, Awaya J, Kurono M, Endo S, Arata Y, Takahashi N. Analyses of N-linked oligosaccharides using a two-dimentional mapping technique. Anal Biochem 1988;171:73-90. 5. Takahashi N, Nakagawa H, Fujikawa K, Kawamura Y, Tomiya N. Three-dimentional elution mapping of pyridylaminated N-linked neutral and sialyl oligosaccharides. Anal Biochem 1995;226:139-146. 6. Jackson P. The analysis of fluorophore labeled saccharides by high- resolution polyacrylamide gel electrophoresis. Advances in Electrophoresis 1994;7:225-262. 7. Baenziger JU. High-performance liquid chromatography of oligosaccharides. In: Methods in Enzymology. Academic Press San Diego 1994;230:237-249. Copyright 1997 Elfos Scientiae The following images related to this document are available:Photo images[ba97034c.jpg]Line drawing images[ba97034a.gif] [ba97034d.gif] [ba97034b.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}