 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
TRANSFECCION DE ADN A CELULAS DE MAMIFEROS
Fidel Ovidio Castro y Yangtse Portelles
Division de Genetica de Celulas de Organismos Superiores. Centro de
Ingenieria Genetica y Biotecnologia. Apartado postal 6162. La Habana,
Cuba.
Recibido en enero de 1996. Aprobado en abril de 1997.
Code Number:BA97037
Size of Files:
Text: 72.7K
Graphics: Line drawings (gif) - 10K
Photographs (jpg) -47K
ABSTRACT Transfection of foreign DNA into living cells in vitro is a powerful research tool in both molecular and cell biology. By means of the different transfection techniques available at present, one can stably change the genetic make-up of a cell as to produce recombinant proteins from it, or test the strenght of a gene promoter coupled to a reporter gene, among many other applications. In this review are discussed the most important transfection techniques with special emphasis in advantages and disadvantages of each one. Reporter gene systems for use in eukaryotic cells are also presented and discussed. As an important conclusion, it is noted that any given transfection protocol needs to be optimized for a given cell line. Key words: transfection, GFP, polyethilenimine, mammalian cells, transgenesis RESUMEN La transfeccion o introduccion de material genetico exogeno a celulas en cultivo es una eficaz herramienta en los estudios de biologia molecular y celular. Mediante las diferentes tecnicas de transfeccion se puede alterar establemente el genoma de una celula para que produzca una proteina recombinante o evaluar de forma rapida y transitoria la fortaleza de un promotor acoplado a un gen reportero, entre otras muchas aplicaciones. Las distintas tecnicas de transfeccion empleadas para celulas de mamiferos son discutidas en esta revision, sobre la base de sus atributos y desventajas. Tambien son abordados los sistemas de genes reporteros mas utilizados en transfecciones transientes. Se concluye que cada metodo de transfeccion debe ser optimizado para una linea celular dada. Palabras claves: transfeccion, GFP, polietilenimina, celulas de mamiferos, transgenesis
Aspectos generales
La posibilidad de introducir ADN en celulas en cultivo ha permitido estudiar mas profundamente las funciones y el control de la expresion de los genes de mamiferos. Existen diferentes metodos para la introduccion de ADN foraneo en celulas de cultivo. Algunos de estos metodos se basan en producir un ambiente quimico adecuado que provoca la adhesion del ADN a la superficie de las celulas de donde es endocitado por mecanismos aun no claramente establecidos; otros implican el uso de un campo electrico para abrir poros en las celulas, a traves de los cuales presumiblemente pasa el ADN al interior de la celula y en los ultimos anos se han generalizado metodos que aprovechan, por una parte, la capacidad de los policationes de condensar sobre si al ADN y por otra la de los liposomas de proteger esta union de la degradacion lisosomal y elevar la capacidad fusogenica de estos complejos. Otras tecnicas como el microbombardeo con particulas cubiertas con ADN, la microinyeccion de genes en los nucleos de celulas y algunas menos comunes se han utilizado para introducir genes en celulas no solo de mamiferos sino en organismos superiores de forma general.
El empleo de uno u otro metodo depende en gran medida de la linea celular que se emplee, el objetivo de la transfeccion, el gen a introducir, la economia del laboratorio y la disponibilidad de equipamiento adecuado. Cada metodo, ademas, debe ser optimizado para cada linea celular antes de ser empleado de forma rutinaria en las transfecciones. La via mas simple para optimizar los parametros de las transfecciones es utilizar un gen reportero , o sea aquel cuyo producto sea facilmente detectable por metodos analiticos no complejos y poco costosos. Los genes reporteros permiten una cuantificacion indirecta de la fortaleza de los promotores, asi como de la eficiencia de los sistemas de transfeccion.
El procedimiento que brinde los resultados mejores en una linea celular dada no necesariamente va a ser optimo para otra, por lo que deben ser ensayados varios y escoger aquel que mayor eficiencia ofrezca. En la Tabla 1 se ofrece una comparacion simplificada de los beneficios de cada metodo de transfeccion. Como se aprecia no existe un metodo que reuna en si todos los atributos optimos tales como: util para transfecciones estables y transientes, facil de realizar, de bajo costo y lo suficientemente versatil como para transfectar cualquier tipo de celula. Tabla 1. Metodos de transfeccion de celulas de mamiferos.
---------------------------------------------------------------------------
Metodo Utilidad Costo Facilidad^a Versatilidad^b Referencias
---------------------------------------------------------------------------
Fosfato de calcio Transiente + + ++ 1-4; 7-10
y estable
DEAE-dextrana Transiente + + ++ 19-21
Electroporacion Transiente +++ + +++ 47-66
y estable
Microinyeccion Estable ++++ +++ + 67
Bombardeo de ADN Transiente +++ ++ + 78-87
y estable
PEI Transiente, + + ? 32
estable?
Lipofeccion Transiente ++ + +++ 35-46
y estable
---------------------------------------------------------------------------
^a Facilidad, se refiere a la complejidad de la tecnica; mayor numero de
cruces implica mayor complejidad^b Versatilidad, se refiere a la aplicabilidad de la tecnica para distintos tipos de celulas en cultivo; mayor numero de cruces indica mayor versatilidad PEI, polietilenimina --------------------------------------------------------------------------- El advenimiento de la tecnologia de los liposomas y mas recientemente el uso de polietilenimina como agente de transfeccion, han representado avances hacia el reactivo universal de transfeccion, pero aun quedan problemas por resolver, como es el costo en el caso de los sistemas comerciales de liposomas y verificar la versatilidad de la polietilenimina para transfecciones estables en una amplia gama de celulas.
De forma general los metodos para introducir ADN en celulas de mamiferos se pueden dividir en tres grupos:
2. Transfeccion mediada por polimeros cationicos acoplados o no a grupos lipidicos (DEAE-dextrana, liposomas, polietilenimina y otros).
3. Transfeccion mediada por efectos fisicos o mecanicos (fusion de protoplastos, electroporacion, microinyeccion, bombardeo de microproyectiles recubiertos con ADN).
Transfeccion mediada por agregados inorganicos (fosfato de calcio) Los coprecipitados de fosfato de calcio (hidroxiapatita) y de ADN purificado se han empleado por mas de 20 anos para la transferencia y expresion de informacion genetica en celulas de mamiferos en cultivo. Esta tecnica fue descrita por vez primera para introducir ADN de adenovirus en celulas de mamiferos por Graham y van der Eb en 1973 (1). Posteriormente se demostro que era posible integrar este ADN exogeno a los cromosomas (2). Esta tecnica ha devenido una de las mas populares para la transfeccion estable o transiente de celulas de mamiferos y ha sido estudiada en detalle por varios grupos en el mundo (3-8).
La precipitacion por fosfato de calcio se ha empleado para el analisis de la replicacion del ADN y la funcionabilidad de los promotores en experimentos transientes en los cuales las celulas son procesadas 48 a 60 h despues de la transfeccion. Es ademas la tecnica mas difundida para la produccion de lineas celulares establemente transformadas. Su superioridad en ese sentido con respecto a la transfeccion mediada por DEAE-dextrana es reconocida. Se cree que una celula transfectada por fosfato de calcio captura mas ADN que una transfectada por DEAE-dextrana (9).
El principio de precipitacion del ADN por el fosfato de calcio es simple, pero la formacion de un coprecipitado optimo de ADN y fosfato de calcio es dificil de repetir debido fundamentalmente a una serie de parametros que afectan la solubilidad del calcio y del fosfato. La mayoria de los autores ha encontrado gran variabilidad en la eficiencia con la cual distintas lineas celulares toman el ADN mediante la precipitacion con fosfato de calcio (8) y normalmente recomiendan protocolos optimizados . Sin embargo se ha estudiado poco la naturaleza de las complejas relaciones entre los distintos componentes que originan la formacion del estado sobresaturado imprescindible para la precipitacion del ADN. Factores que afectan la formacion del precipitado calcio-fosfato- ADN Segun Jordan y colaboradores (8) hay tres aspectos esenciales que afectan la formacion del precipitado, a saber, la concentracion de ADN, la temperatura y el tiempo de la reaccion de precipitacion.
Un exceso de ADN soluble en la solucion sobresaturada de calcio y fosfato puede prolongar el periodo durante el cual se forma el precipitado. Se ha demostrado que a altas concentraciones, el ADN puede incluso evitar la formacion de dicho precipitado (8). Es por esto que reviste especial importancia la preparacion del ADN que se transfectara, ya que errores en la determinacion de la concentracion de los plasmidos o contaminantes presentes en la preparacion, que absorban a 260 nm, van a influir sustancialmente en la naturaleza del precipitado. Es de vital importancia por ende el disponer de una preparacion de ADN de alta pureza y con concentracion minuciosamente corroborada para las transfecciones.
La solubilidad de la hidroxiapatita es mayor a bajas temperaturas y este parametro influyo decisivamente en la eficiencia de la transfeccion de celulas de mosquito segun los autores (8). A pesar de que la hidroxiapatita es considerada insoluble en agua, se ha notado la ausencia de precipitado en soluciones estandares a temperaturas cercanas a los 0 C (7). La mayoria de los protocolos sugieren trabajar a temperatura ambiente, pero la practica ha demostrado que es imprescindible un estricto control sobre la temperatura de la reaccion.
En este sentido son de gran importancia los experimentos de Jordan y colaboradores (8). Este grupo desarrollo una metodologia para evaluar la formacion del precipitado ADN-fosfato de calcio. Esto permite optimizar los parametros y comprobar las soluciones antes de realizar las transfecciones. La esencia del metodo consiste en incubar el ADN con el calcio y el fosfato durante los periodos de tiempo y a temperaturas prefijadas y posteriormente medir la absorbancia de la solucion a 320 nm (en esta longitud de onda absorbe el ADN unido a la hidroxiapatita). A continuacion, se precipita la hidroxiapatita y el ADN por centrifuga rapida. El sobrenadante es leido a 260 nm y en principio solo el ADN no acoplado debe quedar en este.
Esta sencilla metodologia se convierte de hecho en una poderosa herramienta para optimizar las transfecciones mediadas por fosfato de calcio, ya que una vez caracterizadas las nuevas soluciones, incluidos los nuevos lotes de plasmidos, se pueden realizar series de transfecciones bajo las mismas condiciones sin necesidad de reoptimizacion.
La formacion del precipitado es un proceso dinamico que concluye con la transferencia de este al medio de cultivo con las celulas, donde es diluido varias veces. El tiempo durante el cual se permite la formacion del precipitado es un parametro clave. En los protocolos originales (1), se sugiere un tiempo de 30 min para la formacion de un precipitado optimo. Chen y Okayama (3, 4) publicaron la primera optimizacion profunda de los metodos originales y sugieren 20 min. Sin embargo los estudios recientes llevados a cabo en la Universidad de Melbourne (7, 10) demostraron que, por el contrario, la asociacion optima del ADN con el fosfato y el calcio ocurria en una ventana de tiempo extremadamente estrecha entre 0,5 y 1 min. El fundamento racional para disminuir tan drasticamente los tiempos de reaccion se basa en el hecho de que con el tiempo, los precipitados tienden a crecer. El precipitado formado en un minuto tiene la apariencia de un polvillo compuesto por multiples particulas pequenas, muy fino y cubre homogeneamente las celulas, a los cinco minutos se observan menos particulas pero mas grandes y a los 40 min, el precipitado es grosero y mayor incluso que la propia celula (8, Castro, F.O. observaciones no publicadas). Estos autores obtuvieron hasta 100 % de eficiencia de transfeccion empleando esos tiempos de formacion del precipitado en celulas de hepatoma humano HuH7.
Semejante eficiencia absoluta es indudablemente un fenomeno raro, que bien pudiera estar relacionado mas con la estirpe celular que con el metodo en si, ya que solo otro reporte habia sido descrito con anterioridad con semejante eficiencia, para otra linea de hepatomas humanos; HepG2 (11), utilizando los protocolos tradicionales (1). En general se asume que eficiencias de entre 60-70 % son los valores a alcanzar con protocolos optimos, basados en los alcanzados para celulas de rinon de mono verde africano (COS-7; 70 %, 12).
Jordan y colaboradores (8) han confirmado los hallazgos de O Mahoney y Adams, (7) y reportan hasta un 60 % de eficiencia de expresion del gen lacZ en celulas de ovario de hamster chino carentes del gen de la dehidrofolato reductasa (CHO DHFR^-) y hasta 38 % para celulas HEK-293 de rinon embrionario humano, empleando un minuto como tiempo de formacion del precipitado.
En resumen, el ajuste de la concentracion del ADN, la temperatura de la reaccion y el tiempo de duracion de esta, son parametros que han de ser fijados independientemente para cada linea celular. Factores que afectan la eficiencia de la transfeccion en las celulas Asumiendo la formacion de un precipitado optimo, aun quedan otros parametros de vital importancia para una eficiente transfeccion, aquellos relacionados con el comportamiento de las celulas y las condiciones de incubacion de estas con el precipitado.
La causa mas comun de fallos en las transfecciones se cree esta en la estabilidad del pH de la solucion de sales. El rango de pH optimo para la transfeccion es de entre 7,02-7,1 (1). Sin embargo otros autores (7) sugieren pH entre 6,96 y 7,02. El ajuste del pH en rangos tan estrechos cae practicamente dentro de los limites de error de la mayoria de los equipos de determinacion de pH disponibles comercialmente. Por ello como criterio ultimo de eficiencia debe establecerse el de la formacion de un fino precipitado sobre las celulas en un tiempo de aproximadamente 20 min (7, 8). Nuestra experiencia de trabajo con celulas epiteliales de glandula mamaria (HC-11, 13), confirman los hallazgos descritos anteriormente y por ello es precisamente la formacion del precipitado la que ha ser controlada cautelosamente sin prefijar ventanas de tiempo para ello.
Un segundo problema comunmente encontrado es que el medio celular se torna en extremo acido inmediatamente que se anade el coctel de la transfeccion, esto pudiera conllevar a la formacion de un precipitado muy grosero y a la muerte celular. Es este el caso para las transfecciones realizadas en medio RPMI-1640, la mayoria de los grupos ha encontrado que es imprescindible cambiar las celulas a medios menos acidos como el DMEM-alpha antes de la transfeccion, lo cual garantiza la sobrevida de las mismas (14).
El tiempo que las celulas son incubadas con el precipitado varia de acuerdo con la linea celular, asi por ejemplo, celulas HeLa, NIH 3T3, CHO son transfectadas rutinariamente dejando el precipitado actuar por 16 h; otras celulas como COS-7 no pueden sobrevivir tan largos tiempos de exposicion (15). Las celulas epiteliales mamarias HC-11 pueden ser incubadas hasta 20 h con el precipitado sin detrimento para su viabilidad (Portelles, Y., remitido para su publicacion).
Chen y Okayama (4) y Sambrook y colaboradores (16) han publicado variantes de los protocolos originales para celulas que son crecidas en monocapa, pero tripsinizadas durante la transfeccion. Estas variantes incluyen la formacion del precipitado durante una incubacion prolongada (15-24 h) posterior a la siembra de las celulas. La ventaja de este metodo radica en que permite transfectar celulas polarizadas (celulas epiteliales secretoras), las cuales de modo general no son faciles de transfectar debido a que la superficie apical es la unica que tiene acceso al ADN y esta superficie en dichas celulas esta altamente especializada para secretar, por lo cual se supone que sea la causa de la baja eficiencia de transfeccion que normalmente se obtiene en ellas (16).
La eficiencia de transfeccion puede ser aumentada si se emplea el procedimiento de shock osmotico de las celulas con DMSO o glicerol (17). Sin embargo eso es valido solo para algunas lineas celulares como CHO, DUKX, BII, las cuales aumentan dramaticamente su capacidad de transfectarse cuando son sometidas a esos procedimientos, mientras que otras lineas como COS-7, HeLa no responden a los shocks con incremento de la frecuencia de transfeccion (18). En celulas HC-11 no se logra un aumento de la eficiencia mediante el uso de shocks con crioprotectores (Portelles, Y, remitido para su publicacion).
Transfeccion mediada por polimeros cationicos Transfeccion por DEAE-dextrana El diethyl aminoethyl dextrana (DEAE-dextrana) fue empleado originalmente para facilitar la entrada de ARN de poliovirus a celulas (19) y posteriormente para introducir ADN de virus SV40 y poliovirus (20, 21). El metodo, sujeto a modificaciones menores, se emplea aun en la actualidad para la transfeccion de genomas virales y plasmidos a celulas eucariotas.
De forma general esta tecnica difiere de la de fosfato de calcio en tres aspectos: 1) se emplea solo para la transfeccion transiente de celulas, 2) trabaja de forma eficiente para algunas lineas celulares tales como BSC-1, CV-1 y COS-7 (16), pero no para otras, debido a su toxicidad y 3) se emplean menores cantidades de ADN.
Han transcurrido mas de 30 anos desde los primeros reportes del uso de esta metodologia, sin embargo poco ha variado la misma, si bien existen protocolos alternativos en los cuales las celulas son preincubadas con el DEAE-dextrana varias horas antes de ser anadido el ADN (22), o se varia el tiempo de exposicion al DEAE-dextrana en dependencia de su concentracion (16).
El mecanismo por el cual el DEAE-dextrana permite la entrada del ADN a la celula y su transporte hasta el nucleo es poco conocido. Convencionalmente se asume que grandes complejos que contienen tanto ADN como DEAE-dextrana se adhieren a la superficie de la celula y de alguna forma entran mediante endocitosis (23). A pesar del poco conocimiento existente sobre este mecanismo, esta probado que las transfecciones por este metodo son mas reproducibles que aquellas con fosfato de calcio y que son ademas un instrumento eficaz en el estudio de la expresion genica de varios tipos de celulas.
La gran desventaja de esta metodologia comparada con las de fosfato de calcio y como se vera mas adelante tambien con el resto de los metodos, radica en que con DEAE-dextrana solo se logran transfecciones transientes del ADN en la celula huesped (12). No esta claro por que esa diferencia en el destino final del ADN introducido a las celulas por metodos distintos.
Sussman y Milman (24) demostraron que hasta el 80 % de fibroblastos Ltk^- de raton (deficientes en la enzima timidina kinasa) expresaban dicha enzima despues de la transfeccion con el gen tk del virus Herpes simple, empleando DEAE-dextrana como vector. Por otra parte, se determino (25) que una vez dentro de la celula, el ADN anadido por DEAE-dextrana se ensamblaba rapidamente en los nucleosomas y que practicamente todo el ADN exogeno quedaba atrapado en esas estructuras a diferencia del mismo ADN introducido por el metodo de fosfato de calcio, donde se encontraba que al menos parte del ADN estaba en forma de agregados o precipitados.
Lopata y colaboradores (26) mejoraron los protocolos existentes de transfeccion por DEAE-dextrana mediante la adicion de DMSO o glicerol despues de la transfeccion. Estos autores tambien demostraron que la citotoxicidad del DEAE-dextrana debe ser tomada en cuenta, por lo cual deben optimizarse bien las concentraciones a las que sera usado asi como los tiempos de exposicion al mismo para cada linea celular.
Fujita y colaboradores (27) lograron optimizar los protocolos existentes al combinar el shock de DMSO o glicerol y una incubacion con cloroquina o butarato de sodio. Los mecanismos de accion no estan del todo establecidos aun, pero al parecer la cloroquina se une al ADN e inhibe la degradacion intracelular por las hidrolasas lisosomales (28). Por su parte el butarato de sodio pudiera influir en la formacion de una heterocromatina mas activa para la expresion de los genes introducidos (29).
Otros policationes se han empleado para la transfeccion de celulas eucariotas, entre ellos el mas popular es el polibreno. Este polication se ha empleado para facilitar la transfeccion en celulas con demostrada resistencia a la transformacion por metodos convencionales, fundamentalmente por fosfato de calcio (30, 31). Por ejemplo, en celulas CHO, se logra hasta 15 veces mayor eficiencia en transfecciones estables utilizando polibreno que con fosfato de calcio (16). Liposomas, policationes lipidicos comerciales y polietilenimina El desarrollo de las tecnicas de terapia genica en animales se ha basado tradicionalmente en el uso de vectores virales para portar el ADN de interes (32). Las tecnicas no virales estan aun a varios ordenes de magnitud de alcanzar la eficacia de aquellas virales. A pesar de esta limitacion, la terapia genica mediada por plasmidos es estudiada y empleada debido a que no introduce elementos de riesgo virales para la salud del hombre. Es por ello que se ha dedicado mucho esfuerzo en los ultimos anos a la busqueda de vectores no virales. En realidad el advenimiento de la terapia genica basada en vectores no virales ha significado una notable mejoria para la optimizacion de los metodos clasicos de transfeccion, tales como fosfato de calcio y DEAE-dextrana y mas aun, ha despertado el interes por la busqueda de nuevos vectores sinteticos u organicos.
Entre estos destacan el diseno de estructuras cationicas de carga neta altamente positiva para que interactuen con el esqueleto fosforado de la molecula de ADN. De los polimeros cationicos empleados en la actualidad, los que mejores resultados ofrecen en terminos de transfecciones eficientes y estables son: los polimeros de la cascada de las poliamidoaminas y las lipopoliaminas (33, 34). Estas familias de polimeros, aunque muy distintas en su estructura quimica, conservan una similitud adicional a su alta eficiencia de transferencia de ADN a las celulas: ambas contienen residuos protonables aun a pH fisiologico. Esta propiedad es clave pues permite el control de pH en el endosoma y en teoria puede proteger al ADN de la accion de degradacion del sistema lisosomal.
- Lipofectin^TM. Lipofectin^TM es una preparacion de liposomas cationicos compuesta por un nuevo lipido cargado positivamente; DOTMA (N[1-(2-3- dioleiloxi)propil]-N,N,N-trimetilamonio y DOPE (L-dioleoil phosphatidilethanolamina, 38).
Este comportamiento fusogenico de DOTMA provoca una efectiva entrega intracelular de los polinucleotidos, de modo que se obvian las enzimas degradativas presentes en el compartimento lisosomal (41, 42).
Todos estos atributos contribuyen a una transfeccion en extremo sencilla, repetible y eficiente en una amplia variedad de celulas y para un amplio rango de moleculas tales como ADN, ARN, ARN de doble cadena y oligonucleotidos (43).
Las ventajas mas importantes de Tfx^TM-50 sobre otros sistemas lipidicos existentes incluido el de Lipofectin^TM, son su sencillez, ya que se requiere solo un tubo de cultivo para todas las reacciones y no emplea solucion tamponada especial. Las transfecciones se pueden realizar de modo optimo en un tiempo de solo dos horas, lo que le confiere otra ventaja sobre los metodos usuales, que toman entre 4 y 20 h.
El precio de estos polimeros si bien no es en extremo alto (oscila entre los 150-220 dolares para aproxidamente 50-100 transfecciones), no es asequible para trabajo rutinario en muchos laboratorios.
Estructuralmente la PEI es el polimero organico de mayor carga positiva que existe debido a que, cada dos atomos de carbono hay un nitrogeno de tipo amino secundario, el cual puede ser protonado. Esto provoca la union del polimero a los grupos fosfato del ADN, quedando aun grupos positivos capaces de unirse a la membrana celular. La estructura quimica de la PEI protonada es la siguiente:
La PEI mantiene la capacidad de conservar esa estructura incluso a pH fisiologico, se piensa que esta simple propiedad molecular de tampon a cualquier pH le confiere al PEI su eficiencia en las transfecciones.
Boussif y colaboradores (32) demostraron que en 1 uL de una solucion stock de 10 mM de PEI existen 10 nmol de nitrogeno aminados, mientras que 1 ug de ADN posee 3 nmol de fosfato. El exceso molar de aminos protonables con respecto a fosforo inorganico en el ADN, fue empleado como medida de optimizacion de la transfeccion. Se emplearon proporciones desde 3 hasta 135 veces de exceso de PEI con respecto al ADN y resulto optima la proporcion de alrededor de 10 veces. Nuestros resultados empleando celulas epiteliales mamarias murinas HC-11 y CID-9, asi como COS-7 y CHO coinciden con lo reportado (32). La proporcion optima de PEI:ADN fue 9:1. Cuando se elevo esta al doble (18:1), la eficiencia se disminuyo drasticamente (Portelles, Y., remitido para su publicacion, Castro, FO, observaciones no publicadas).
El uso de la polietilenimina es aun incipiente. Boussif y colaboradores (32) transfectaron diferentes lineas celulares y en todos los casos se obtuvieron resultados positivos. Las lineas en cuestion fueron: 3T3, HepG2, COS-7, HeLa y MRC-5. Ademas se logro, empleando PEI como vector, la expresion in vivo en cerebro de ratones neonatos, de un gen reportero y de pequenos fragmentos de oligonucleotidos. Este ultimo resultado es muy importante ya que ofrece la posibilidad de usar PEI como vector universal para transferencia de genes in vivo e in vitro. Electroporacion de celulas de mamiferos La electroporacion es una tecnica de introduccion de ADN a celulas en cultivo y se lleva a cabo mediante la aplicacion de pulsos electricos a las celulas, lo que provoca la apertura de poros en la membrana de estas con la consiguiente penetracion del ADN presente en el medio de electroporacion. El pulso electrico puede ser de caida exponencial o cuadrado, sin que al parecer existan ventajas obvias de una u otra forma de pulso con respecto a la eficiencia de la transfeccion (47). La mayoria de los estudios se ha basado en pulsos exponenciales o cuadrados unicos (47). Recientemente se publico el uso de tandems de pulsos cuadrados para cultivos de alta densidad de celulas CHO en suspension, con una elevada eficiencia. Hasta el 60 % de las celulas resultaron transfectadas luego de recibir 10 pulsos cuadrados de 5 ms de duracion y con una intensidad de 0,6 kV/cm. Los autores disenaron un sistema de flujo para la transfeccion de grandes volumenes celulares, que alcanzaron una eficiencia de hasta el 25 % de transfeccion.
La electroporacion se ha empleado para introducir ADN en una amplia variedad de celulas animales (47, 49-53), de plantas (54) y tambien en bacterias y levaduras (55). La electroporacion puede ser usada indistintamente para expresion transiente o estable. La eficiencia de la transfeccion por electroporacion esta influida por una serie de factores, entre los cuales destacan:
2. Duracion del pulso electrico. Normalmente se deja pasar un pulso unico a traves de la suspension celular. En el caso de los pulsos cuadrados, se ha hallado que la descarga de varias replicas de estos aumenta la eficiencia de transfeccion para algunas lineas celulares. Entre 0,2-10 ms esta el tiempo optimo de electroporacion (58-60). 3. Temperatura. Algunos autores reportan que los niveles maximos de transfeccion se logran cuando las celulas son mantenidas a temperatura ambiente durante la electroporacion (61); otros indican que es mejor mantener las celulas a 0 C (59). Esas discrepancias pueden resultar de las diferencias en la respuesta de distintas lineas celulares al paso de la corriente, o en la cantidad de calor generado durante la electroporacion con altos valores de voltaje (> 1 000 V/cm) y/o tiempos superiores a los 100 ms. 4. Concentracion y tipo de ADN. Si bien se puede transfectar mediante electroporacion tanto ADN circular como lineal, los mayores niveles de expresion tanto transientes como estables, se obtienen cuando se usa ADN lineal (47, 50, 52). La concentracion optima de ADN para las transfecciones se encuentra en el rango de 1 a 40 ug/mL. 5. Composicion ionica del medio. La constante de tiempo de pulso (ç) es afectada dramaticamente por la composicion del medio de electroporacion, asi las celulas resuspendidas en soluciones salinas tamponadas tales como: PBS, Buffer-Hepes salino, o medios de cultivo celulares permiten una menor duracion del pulso electrico. El empleo de sustancias no ionicas (manitol, sacarosa, etc.) en soluciones tamponadas aumenta la constante de tiempo de pulso (57).
A diferencia de la transfeccion por fosfato de calcio o de la fusion de protoplastos (ver mas adelante), la electroporacion por lo general conlleva a la formacion de lineas celulares establemente transformadas con la integracion de una sola copia del ADN foraneo (62-64).
La electroporacion se ha usado tambien para transferir ADN foraneo a espermatozoides bovinos para su uso en transgenesis mediada por inseminacion artificial (65). A partir de 1986, la electroporacion se ha empleado como el metodo de eleccion para la transfeccion de celulas embrionarias totipotentes (embryonic stem cells) tanto en experimentos de gene knock-out, como de reemplazamiento alelico y recombinacion de homologos (66).
En terminos generales la mayor desventaja de la electroporacion es su costo, ya que la mayoria de los equipos comercialmente disponibles son caros, en el orden de los miles de dolares, ademas de los accesorios como cubetas y adaptadores que tambien lo son (67). Microinyeccion directa de ADN en el nucleo de celulas Este metodo fue desarrollado por Capecchi (68) y si bien tiene la ventaja de no exponer los polinucleotidos a compartimentos celulares tales como el endosoma de bajo pH, no puede ser empleado de forma rutinaria para introducir ADN a un gran numero de celulas como para permitir despues analisis bioquimicos. Por ello, la microinyeccion es usada basicamente como un metodo para establecer lineas que lleven copias integradas del ADN de interes.
Esta tecnica requiere de infraestructura, equipamiento y personal altamente entrenado para realizarla, por lo que su costo es relativamente alto. Las tecnologias desarrolladas por Capecchi (68) para celulas fueron adaptadas exitosamente a embriones de una gran cantidad de especies, para abrir paso a la modificacion genetica de animales o transgenesis (69).
La microinyeccion de embriones de especies de mamiferos, anfibios y peces ha devenido una forma util de probar construcciones geneticas directamente en los embriones y en algunas estirpes celulares derivadas de ellos (70). En nuestro laboratorio se ha desarrollado con especial interes esta tecnologia para la produccion de animales transgenicos. De gran utilidad ha sido el empleo de embriones de tilapia y carpas para el ensayo transiente de distintos promotores unidos a genes reporteros. Estos sistemas permitieron evaluar de forma rapida las distintas construcciones obviando el uso de celulas de peces, que son mas complejas de mantener y de transfectar que las celulas de mamiferos (71).
Por otra parte embriones de conejos microinyectados con el plasmido pCH110 que contiene el gen bacteriano lacZ fueron empleados como celulas transientemente transfectadas para determinar el tiempo optimo de microinyeccion en experimentos de generacion de conejos transgenicos con expresion de genes en la glandula mamaria (72). De la Fuente y colaboradores (manuscrito enviado para su publicacion) han utilizado el sistema de embriones murinos para el ensayo transiente de promotores de genes de interferones humanos. Fusion de protoplastos Desarrollado por Schaffner en 1980 (73) y perfeccionado por Rassoulzadegan en 1982 (74), el metodo consiste en obtener protoplastos de bacterias que llevan alto numero de copias del plasmido de interes y fundirlos directamente con las celulas de mamiferos en cultivo. Para ello, el plasmido de interes se multiplica en la bacteria, en presencia de cloranfenicol para amplificar el ADN; posteriormente se rompe la pared celular bacteriana con lisozima. Los protoplastos resultantes son centrifugados sobre una monocapa de celulas superiores y tratados con un agente de fusion (comunmente se emplea el polietilen glicol; PEG). Como resultado del proceso de fusion de las membranas celulares, el contenido de las bacterias es depositado en el citoplasma de la celula superior y el ADN plasmidico es transferido al nucleo de esta. El PEG es retirado y las celulas tratadas con kanamicina para inhibir el crecimiento de posibles bacterias sobrevivientes.
La fusion de protoplastos, a pesar de no ser de los metodos mas eficientes para las lineas celulares normalmente usadas en los laboratorios, se ha utilizado tanto en transfecciones transientes como estables, fundamentalmente para aquellas lineas celulares en las cuales el proceso de endocitosis ocurre de forma ineficiente. Asi Gillies y colaboradores (75) introdujeron de forma estable, genes de inmunoglobulinas en celulas B murinas y genes de globina en celulas eritroleucemicas de raton fueron introducidos por Charnay y colaboradores (76).
Para estos tipos de celulas, la fusion de protoplastos puede llegar a ser de altisima eficiencia. Schaffner (73) reporto hasta 6 % de celulas transientemente expresando un gen, empleando para ello una proporcion de 10 000 bacterias por celula de mamifero. Mientras que Rassoulzadegan y colaboradores (74), obtuvieron 100 % de transfectantes transientes y 0,02 % de transfectantes estables empleando otra bacteria hospedera y la misma proporcion que Schaffner (73).
La gran ventaja del metodo radica en su eficiencia para ciertos tipos celulares, mientras que su desventaja es la gran cantidad de manipulaciones que conlleva, el tiempo de trabajo y que normalmente no son posibles las cotransfecciones, por lo que el gen de interes tiene que estar siempre contenido en un plasmido junto con el marcador de seleccion (16).
Como resultado de la fusion, por lo general se obtienen multiples copias del plasmido integradas en forma de tandems repetitivos en el cromosoma huesped (77). Bombardeo de celulas y tejidos con microproyectiles En 1984, Sandford y colaboradores desarrollaron un equipo destinado a la introduccion de ADN en celulas y tejidos de distintas especies de plantas. En 1987 se publica el primer reporte del uso de microproyectiles en transferencia de genes (78). Los primeros experimentos se realizaron acelerando en una bolsa de aire, particulas de tungsteno cubiertas con ADN. Sin embargo la bolsa de aire era mortal para las celulas y esto conllevo al diseno de nuevos equipos que permitian otras formas de descarga de las particulas (79). La expresion transiente del ARN del virus del mosaico del tabaco y del gen de cloranfenicol acetil transferasa (CAT) en celulas de cebolla, demostraron que el ADN permanecia activo despues del bombardeo (79- 80). La biobalistica, como se le ha denominado a esta tecnologia, ha logrado espectaculares avances en la transgenesis de plantas, no asi en mamiferos (81, 82) y su uso es aun limitado.
Dos equipos comercialmente disponibles (Biolistic^R^ y Accell^R ) a traves de compra o alquiler en Bio-Rad (Hercules, CA, USA) han sido modificados para su uso en bombardeo de celulas de mamiferos (83).
El primer reporte de transformacion de celulas de mamiferos empleando microproyectiles fue publicado en 1989 (84). Este grupo introdujo el gen de la resistencia a neomicina en fibroblastos NIH/3T3. Otros reportes han aparecido en los ultimos anos que incluyen la transformacion de ovocitos de peces (85), celulas humanas, fibroblastos de pollo, miotubulos esqueleticos, celulas epiteliales y miocardiocitos (86). En algunos casos, la eficiencia de transformacion ha excedido a las que se obtienen por los metodos tradicionales (82, 87). El rango de eficiencia vario desde 6 x 10^- 4 hasta 1,7 x 10^-3 (87). Recientemente tambien se adapto la metodologia para celulas en suspension, y se ha logrado transfectar celulas linfoides humanas (88).
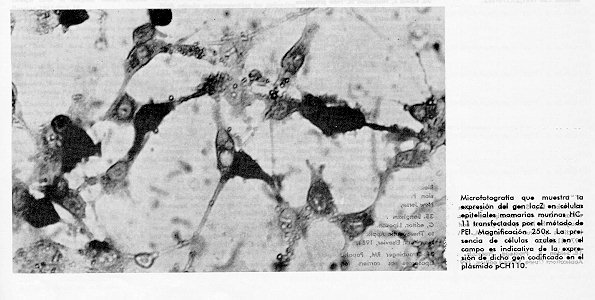
No obstante estos avances, la biobalistica dista mucho de ser una tecnica de uso rutinario, por su carestia, dificultades tecnicas y por la existencia de otros metodos mucho mas simples y costeables. Esta metodologia pudiera ser de gran utilidad en el estudio de la regulacion de la expresion de los genes de leche, si se lograra la introduccion de ADN en explantes de glandula mamaria de las especies de interes economico. Transfeccion: la eleccion de un metodo Como se aprecia, el uso de uno u otro metodo de transfeccion para una linea celular dada no es una cuestion trivial. En la Tabla 2, aparecen resumidos los resultados de nuestro laboratorio en la optimizacion de la transfeccion de celulas epiteliales mamarias murinas HC-11 (13), gentilmente cedidas por el Dr. Bernd Groner, (University of Freiburg, Germany). Estas celulas fueron transfectadas con el gen lacZ de E. coli por 5 metodos distintos, incluido una modificacion del protocolo tradicional de calcio fosfato, referida anteriormente como metodo acelerado de Jordan y colaboradores (8). Tabla 2. Resumen de los resultados obtenidos con los diferentes metodos de transfeccion empleados en celulas HC-11 y valor de uso de los mismos para dichas celulas.
---------------------------------------------------------------------------
Metodo Resultados Uso recomendado Observaciones
---------------------------------------------------------------------------
Fosfato de calcio ~0,1% eficiencia Transfeccion Ensayar con celulas
tradicional estable en suspension
Fosfato de calcio 0 % Ninguno Estudiar variantes
"acelerado"
DEAE-dextrana ~0,1% eficiencia Ninguno -
PEI ~3-5% eficiencia Transfeccion Valorar en
transiente transfecciones
estables
Electroporacion 0 % Ninguno Estudiar variantes
de optimizacion
---------------------------------------------------------------------------
Las celulas HC-11 resultaron ser totalmente refractarias a la transfeccion por el metodo acelerado de Jordan y colaboradores, electroporacion y a la transfeccion por DEAE-dextrana. El uso de la metodologia de fosfato de calcio tradicional arrojo una baja eficiencia de transfeccion en celulas HC-11, mientras que PEI permitio obtener buenos resultados en las transfecciones realizadas. De los resultados alcanzados, podemos concluir que en nuestras condiciones y para celulas HC-11 en el mismo pase e igual fase de crecimiento, el metodo tradicional de fosfato de calcio puede ser empleado con su eficiencia actual para transfecciones estables, ya que rinde al menos 1 000 celulas transfectadas por placa de 60 mm. PEI puede ser usada para transfecciones transientes y se han alcanzado ya transfectantes estables tanto de celulas HC-11 como de CHO (Fernandez y colaboradores, manuscrito en preparacion). Los genes reporteros en las transfecciones de celulas de mamiferos Como se ha venido discutiendo hasta ahora, los metodos de introduccion de ADN foraneo en celulas de mamiferos son variados y la eleccion de uno u otro depende de multiples factores, entre los cuales se destaca el objetivo de la transfeccion. De este modo en ocasiones se necesita generar una linea celular con un caracter nuevo incorporado (transfeccion estable), y a veces se persigue otro objetivo experimental que no requiere la integracion del ADN transfectado al genoma celular. A este tipo de transfeccion se le conoce con el nombre de transfeccion transitiva o transitoria y es ampliamente empleada en los laboratorios de biologia celular y molecular.
De especial interes es el ensayo de construcciones geneticas en transfecciones transientes. Esto permite: 1) conocer el correcto funcionamiento de una construccion, antes de emprender un proyecto mayor con ella (transgenesis, transfeccion estable, etc.); 2) ensayar varias construcciones y seleccionar la mejor, expresada en terminos de calidad y cantidad del ARN mensajero deseado, niveles de expresion del transgen, actividad biologica de la proteina secretada, etc. y 3) economizar recursos por los datos que aportan los puntos 1 y 2 anteriormente descritos.
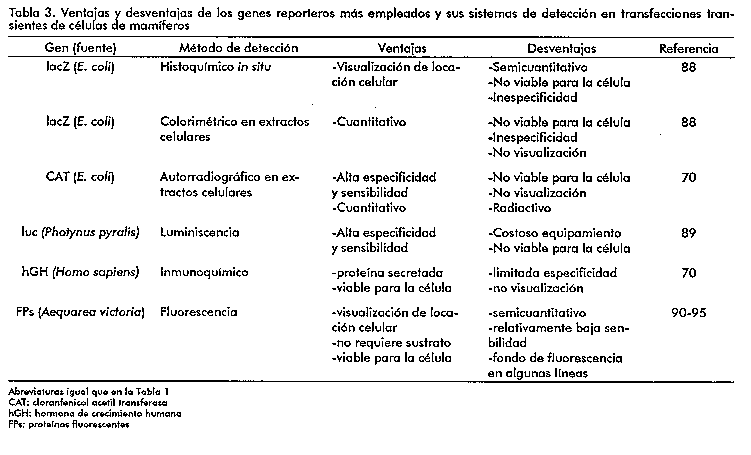
En las transfecciones transientes, se emplean de forma rutinaria los genes reporteros. Los reporteros mas utilizados han sido el gen lacZ de E. coli que codifica para la beta-galactosidasa bacteriana (89), el gen de la cloranfenicol acetil transferasa de bacterias, la hormona de crecimiento humano (71), el gen de la luciferasa de las luciernagas (90) y mas recientemente los genes que codifican para las proteinas fluorescentes verdes (GFP) y azul (BFP, (91) siglas en ingles en ambos casos) de la medusa (Aequorea victoria).
En la Tabla 3 se resumen las ventajas y desventajas en el uso de los genes reporteros anteriormente mencionados. Como se aprecia, cada gen reportero, con su metodo especifico de deteccion, presenta ventajas y desventajas en su aplicacion, por lo que no se debe recomendar el uso de uno u otro a priori, sin antes analizar minuciosamente los objetivos y posibilidades reales de cada laboratorio.
Los genes lacZ, CAT, luc y hGH han sido empleados como reporteros desde mediados de la decada de los 80, por lo que no seran analizados en detalle en esta revision. De especial interes resulta, sin embargo, el auge que ha ido teniendo en los ultimos anos el empleo como reporteros de genes de proteinas fluorescentes de la medusa (91-93) lo cual se ha denominado "revolucion verde".
La proteina verde fluorescente monomerica (GFP) consiste en 238 aminoacidos y posee un peso molecular de 27 KDa (94) y no requiere de ninguna otra proteina del hospedero, sustrato o cofactores para fluorescer. Esta caracteristica la hace superior como reportero a otras enzimas de E. coli como la beta-galactosidasa que precisa del transporte a traves de la membrana del sustrato fluorogenico (95).
En su forma nativa, GFP emite luz verde brillante a una longitud de onda de 508 nm cuando es excitada con luz ultravioleta de hasta 395 nm, o fluoresce levemente cuando es excitada con luz azul de hasta 470 nm de longitud de onda. La fluorescencia con luz ultravioleta es intensa pero desaparece rapidamente, mientras que con luz azul es tenue pero sostenida (95). Los microscopios opticos de rutina en los laboratorios contienen filtros destinados a excitar la fluoresceina o sus derivados a aproximadamente 480 nm; con ello se logran emisiones en verde alrededor de los 530 nm. Por otra parte, los equipos de citometria de flujo de un solo rayo laser ionico de argon excitan a 488 nm, por lo que de modo general tanto microscopios convencionales como citometros de flujo resultan suboptimos para el estudio de la GFP como reportero.
Todos estos elementos llevaron a la busqueda de mutantes de GFP con picos de excitacion de alrededor de los 490 nm y con picos de emision entre los 505-511 nm. Se han generado ya mutantes red-shifted y blue- shifted, con hasta 24 veces mas intensidad de emision que la variante natural (95, 96) que en combinacion con la GFP natural ofrecen una amplia gama de posibilidades combinatorias no solo ya para ensayos en celulas, sino tambien para terapia genica.
Recientemente se han introducido otras mejoras en los sistemas de GFP, tales como cambio en el uso de codones hacia aquellos mas utilizados por las celulas de mamiferos. Esto permitio obtener lineas celulares transfectadas con una sola copia del transgen y la activacion simultanea de dos elementos transcripcionales distintos de GFP en una misma linea celular (95, 96) .
La principal desventaja del uso de GFP y sus mutantes radica en su limite de sensibilidad y el fondo de fluorescencia que normalmente presentan algunas lineas de celulas de mamiferos. Mientras que la proteina beta- galactosidasa tetramerica cataliza en 1 h > 10^5 moleculas de sustrato, la expresion de una copia de GFP por celula es apenas detectable en un microscopio de fluorescencia convencional (93).
Recientemente se ha publicado el uso de una variante de GFP que porta codones de uso preferencial en celulas de mamiferos como gen reportero en ratones transgenicos; la deteccion del transgen es en extremo sencilla, mediante la iluminacion breve de un dedo o un fragmento de cola de los ratones a ensayar con luz ultravioleta (97).
El uso de GFP como reportero en experimentos de generacion de pez-cebra y ratones transgenicos esta siendo explotado intensamente en nuestro laboratorio en la actualidad (Morales, R, comunicaciones personales; Castro, FO, no publicado).
Otro grupo de marcadores o reporteros para transferencia genica en celulas de mamiferos son aquellos que codifican a la resistencia a una droga. Mediante la seleccion de colonias resistentes a dicha droga, se logran aislar clones independientes que han integrado este marcador de seleccion. Si bien estos marcadores permiten seleccionar aquellas celulas vivas que expresan el transgen, es preciso que la celula sobreviva en un ambiente toxico por un periodo de tiempo generalmente largo. Por ello estos marcadores son empleados solo para transfecciones estables. Los mas comunmente usados son: resistencia a neomicina (GPT) y resistencia a higromicina (HigroB). Para algunas lineas celulares carentes de alguna enzima especifica (CHO DHFR^- por ejemplo), se emplea medio de cultivo sin nucleosidos para seleccionar las lineas que posteriormente son amplificadas con la droga metrotexate.
Conclusiones La optimizacion de los sistemas actuales de transfeccion de celulas de mamiferos, unida a la busqueda de nuevos vectores y al mejoramiento de los genes reporteros, permitira un sustancial avance en los metodos de estudio de la expresion de genes in vitro. Notables avances se preven tambien en el area de la introduccion de material genetico exogeno a explantes, organoides y grupos de tejidos ex vivo, asi como directamente a animales in vivo mediante tecnicas de terapia genica. Es indudable que la transfeccion de ADN en celulas de mamiferos a pesar de sus grandes avances en las dos ultimas decadas, aun espera por perfeccionamiento y depara sorpresas para los proximos anos.
1. Graham FL, van der Eb AJ. A new technique for the assay of infectivity of human adenovirus 5 DNA. Virology 1973;52:456-461. 2. Wigler M, Pellicer A, Silverstein S, Axel R. Biochemical transfer of single-copy eucaryotic genes using total cellular DNA as donor. Cell 1978;14:725-732. 3. Chen C, Okayama H. High-efficiency transformation of mammalian cells plasmid DNA. Mol. Cell Biol 1987;7:2745-2752. 4. Chen C, Okayama H. Calcium phosphate-mediated gene transfer: A highly efficient transfection system for stably transformingcells with plasmid DNA. BioTechniques 1988;6:632. 5. Alam J, Cook JL. Reporter genes: Application to the study of the mammalian cells transcription. Anal Biochem 1990;188:245-254. 6. Farr A, Roman A. A pitfall of using a second plasmid to determine transfection efficiency. Nucleic Acids Res 1992;20:920. 7. O'Mahoney JV, Adams TE. Optimization of experimental variables influencing reporter gene expression in hepatoma cells following calcium phosphate transfection. DNA and Cell Biol 1994;13:1227-1232. 8. Jordan M, Schallhorn A, Wurm FM. Transfecting mammalian cells: Optimization of critical parameters afecting calcium-phosphate precipitate formation. Nucl Acids Res 1996;24:596-601. 9. Kingston J, editor. Current Protocols in Molecular Biology. Oxford: Currents Protocols Ltd, 1987. 10. O'Mahoney JV, Brandon MR, Adams TE. Identification of a liver-specific promoter for the ovine growth hormone receptor. Mol Cell Endocrinol 1994;101:129-139. 11. Aden DT, Knowles BB. Controlled synthesis of HBsAg in a differentiated human carcinoma-derived cell line. Nature 1979; 282:615-616. 12. Gluzman Y. SV40-transformed simian cells support the replication of early SV40 mutants. Cell 1981;23:175-182.
13. Ball RK, Friis RR, Schoenenberger CA, Doppler W, Groner B. Prolactin regulation of beta-casein gene expression and of a cytosolic 120-kd protein in a cloned mouse mammary epithelial cell line. EMBO J 1988;7:2089-2095. 14. Burdon TG, Maitland KA, Clark AJ, Wallace R, Watson CJ. Regulation of the sheep beta-lactoglobulin gene by lactogenic hormones is mediated by a transcription factor that binds an interferon- activation site-related element. Mol Endocrinology 1994;8:1528-1536. 15. Selden R. DNA Transfection. In: Kingston J, editor. Current Protocols in Molecular Biology. Oxford: Currents Protocols Ltd, 1987. 16. Sambrook J, Fritsch D, Maniatis T. Molecular Cloning. A Laboratory Manual. Second edition. Cold Spring Harbor Press, 1989. 17. Parker BA, Stark GR. Regulation of simian virus 40 transcription: Sensitive analysis of the RNA species present early in infections by virus or viral DNA. J Virol 1979;31:360. 18. Selden R. Promega Protocols and Applications Guide, Second edition, 1991. 19. Vaheri A, Pagano JS. Infectious poliovirus RNA: A sensitive method of assay. Virology 1965;27:434. 20. McCutchan JH, Pagano JS. Enhancement of the infectivity of simian virus 40 deoxyribonucleic acid with diethyl aminoethyl-dextran. J Natl Cancer Inst 1968; 41:351-359. 21. Warden D, and Thorne HV. Infectivity of polyoma virus DNA for mouse embryo cells in presence of diethylaminoethyl-dextran. J Gen Virol 1968;3:371-375. 22. Eukaryotic Gene Regulation. In: Promega Protocols and Applications Guide, Second edition, 1991:282-310. 23. Selden RF, Burke-Howie K, Rowe ME, Goodman HM, Moore DD. Human growth hormone as a reporter gene in regulation studies employing transient gene expression. Mol Cell Biol 1986;6:3173-3179. 24. Sussman DJ, Milman G. Short-term, high-efficiency expression of transfected ADN. Mol Cell Biol 1984;4:1641-1647. 25. Reeves R, Gorman C, Howard B. Minichromosome assembly of nonintegrated plasmid DNA transfected into mammalian cells. Nucl Acids Res 1985;13:3599- 3607. 26. Lopata MA, Cleveland DW, Sollnerweb B. High level of a chloramphenicol acetyltransferase gene by DEAE-dextran-mediated DNA transfection coupled with a dimethyl sulfoxide or glycerol shock treament. Nucl Acids Res 1984;12:5707. 27. Fujita T, Shubiya H, Ohashi T, Yamanishi K, Taniguchi T. Regulation of human interleukin-2 gene: Funcional DNA sequences in the 5' flanking region for the gene expression in activated T lymphocytes. Cell 1986;46:401- 407. 28. Luthman H, Magnusson G. High efficiency DNA transfection of chloroquine treated cells. Nucl Acids Res 1983;11:1295. 29. Reeves G. DNA Expression. In: Kingston J, editor. Current Protocols in Molecular Biology. Oxford: Currents Protocols Ltd, 1987:9.4.1-9.4.3. 30. Kawai S, Nishizawa M. New procedure for DNA transfection with polycation and dimethyl sulfoxide. Mol Cell Biol 1984;4: 1172-1177. 31. Chaney WG, Howard DR, Pollard A, Sallustio S, Stanley P. High- frequency transfection of CHO cells using Polybrene. Somatic Cell Mol Genet 1986;12:237-247. 32. Boussif O, Lezoualc'h F, Zanta MA, Mergny MD, Scherman D, Demeneix B, Behr JP. A versatile vector for gene and oligonucleotide transfer into cells in culture and in vivo: Polyethylenimine. Proc Natl Acad Sci USA 1995;92:7297-7301. 33. Behr JP, Loeffler JP, Demeneix B, Perez-Mutul J. Efficient gene transfer into mammalian primary endocrine cells with lipopolyamine-coated DNA. Proc Natl Acad Sci USA 1989;86:6982-6986. 34. Felgner PL. In: Methods in Molecular Biology, Vol 7: Gene Transfer and Expression Protocols. Murray EJ, editor Clifton, New Jersey. The Human Press Inc, 1991. 35. Bangham AD. Introduction. In: Knight G, editor. Liposomes from Physical Structure to Therapeutic Application. North-Holland, New York: Elsevier, 1981. 36. Straubinger RM, Papahadjopoulos D. Liposomes as carriers for intracellular delivery of nucleic acids. Methods Enzymol 1983;101:512- 527. 37. Cudd A, Nicolau C. Entrapment of recombinant DNA in liposomes and its transfer and expression of eukaryotic cells. Liposome Technology vol. II. Boca Raton: CRC, 1984:207-221. 38. Felgner PL, Gadek TR, Holm M, Roman R, Chan HW, Wenze M et al. Lipofection: A highly efficient, lipid-mediated DNA-transfection procedure. Proc Natl Acad Sci USA 1987;84:7413-7417. 39. Israelachvili JN, Mitchell DJ, Ninham BW. Theory of self-assembly of lipid bilayers and vesicles. Biochim Biophys Acta 1977;470:185-201. 40. Felgner PL, Holm M. Cationic liposome-mediated transfection. Focus 1989;11:21-25. 41. Duzgunes N, Goldstein JA, Friend DS, Felgner PL. Fusion of liposomes containing a novel cationic lipid N[1-(2,3-Dioleyloxy) Propyl]-N,N,N- Trimethylammonium: Induction by multivalent anions and asymmetric fusions with acidic phospholipid vesicles. Biochem 1989;28:9179-9184. 42. Felgner PL, Ringold GM. Cationic liposome-mediated transfection. Nature 1989;337:387-388. 43. Malone R, Felgner PL, Verma I. Lipofectin-mediated RNA transfection. Proc Natl Acad Sci USA 1989;86:6077-6081. 44. Promega Corporation. Technical Bulletin 1995;216: 53. 45. Promega Notes 1995;56:58. 46. Promega Corporation. Reporter and transfection systems, 1995:193. 47. Neumann E, Schaefer-Ridder M, Wang Y, Hofschneider PH. Gene transfer into mouse lyoma cells by electroporation in high electric fields. EMBO J 1982;1:841-849. 48. Rols MP, Coulet D, Teissie J. Highly efficient transfection of mammalian cells by electric field pulses. Application to large volumes of cell culture by using a flow system. Eur J Biochem 1992;206:115-121. 49. Wong TK, Neumann E. Electric field mediated gene transfer. Biochem. Biophys Res Commun 1982;107:584-587. 50. Potter H, Weir L, Leder P. Enhancer-dependent expression of human k inmunoglobulin genes introduced into mouse pre-B lymphocytes by electroporation. Proc Natl Acad Sci USA 1984;81:7161-7168. 51. Sugden B, Marsh K, Yates J. A vector that replicates as a plasmid and can be efficiently selected in B-lymphoblast transformed by Epstein-Barr virus. Mol Cell Biol 1985;5:410-419. 52. Toneguzzo F, Hayday AC, Keating A. Electric field-mediated DNA transfer: Transient and stable gene expression in human and mouse lymphoid cells Mol Cell Biol 1986;6:703-713. 53. Tur-Kaspa R, Teicher L, Levine BJ, Skoultchi AI, Shafritz DA. Use of electroporation to introduce biologically active foreign genes into primary rat hepatocytes. Mol Cell Biol 1986;6:716-728. 54. Fromm M, Taylor LP, Walbot V. Expression of genes transferred into monocot and dicot plant cells by electroporation. Proc Natl Acad Sci USA 1985;82:5824-5831. 55. Andreason GL, Evans GA. Introduction and expression of DNA molecules in eukariotic cell by electroporation. BioTechniques 1988;6:50-53. 56. Patterson MK Jr. Measurement of growth and viability of cells in culture. Methods Enzimol 1979;58:141-149. 57. Rabussay D, Uher L, Bates G, Piastuch A. Electroporation of mammalian and plant cells. Bethesda Res Lab Focus 1987;9:1. 58. Chu G, Hayakawa H, Berg P. Electroporation for the efficient transfection of mammalian cells with DNA. Nucleic Acids Res 1987;15:1311- 1313. 59. Reiss M, Jastreboff MM, Bertino JR, Narayanan R. DNA-mediated gene transfer into epidermal cells using electroporation. Biochem Biophys Res Commun 1986;137: 244-248. 60. Boggs SS, Gregg RG, Borenstein N, Smithies O. Efficient transformation and frequent single-site, single-copy insertion of DNA can be obtained in mouse erythroleukemia cells transformed by electroporation. Exp Hematol 1986;14:988. 61. Hama-Inaba H, Takahashi M, Kasai M, Shiomi T, Ito A, Hanaoka F, Sato K. Optimum conditions for electric pulse-mediated gene transfer to mammalian cells in suspension. Cell Struct Funct 1987;12:173. 62. Wolf H, Rols M, Boldt P, Neumann E, Teissie J. Control by pulse parameters of electric field-mediated gene transfer in mammalian cells. Biophys J 1994;66:524-531. 63. Kubiniec RT, Liang H, Hui SW. Effects of pulse length and strength on transfection by electroporation. Biotechniques 1990;8:1-3. 64. Ray FA. Electroporation of plasmid DNA into normal human fibroblasts. In: Nickoloff, JA, editor. Animal Cell Electroporation and electrofusion protocols. Methods in Molecular Biology. Vol. 48. Totowa, New Jersey: Humana Press, 1995:133-140. 65. Gagne M, Pothier F, Sirard H. Electroporation of bovine spermatozoa to carry foreign DNA in oocytes. Molec Reprod Develop 1991;29:6-15. 66. Gossler A, Doetschman T, Korn R, Serfling E, Kenler R. Transgenesis by means of blastocysts-derived embryonic stem cell lines. Proc Natl Acad Sci USA 1986;83: 9065-9069. 67. Nickoloff, JA. Animal Cell Electroporation and electrofusion protocols. Methods in Molecular Biology. Vol. 48. Totowa, New Jersey: Humana Press, 1995:133-140. 68. Capecchi MR. High efficiency transformation by direct microinjection of DNA into cultured mammalian cells. Cell 1980;22: 249-260. 69. Gordon JW, Scangos GA, Plotkin DJ, Barbosa JA, Ruddle FH. Genetic transformation of mouse embryos by microinjection of purified DNA. Proc Natl Acad Sci USA 1980;77:7380-7384. 70. Castro FO, de la Fuente J. Animales transgenicos. Posibilidades biotecnologicas. Interferon y Biotecnologia 1988;5:210-223. 71. Lleonart R, Martinez R, Garcia del Barco D, Hernandez O, Castro FO, de la Fuente J. Reporter genes for in vivo transient gene expression studies in tilapia (Oreochromis aureus) and common carp (Cyprinus carpio) one celled embryos [abstract]. Theriogenology 1994;41:240 72. Ramos B, de Armas R, de la Fuente J, Castro FO. Activity of simian virus 40 early promoter in rabbit embryos [abstract]. Theriogenology 1994;41:281. 73. Schaffner W. Direct transfer of cloned genes from bacteria to mammalian cells. Proc Natl Acad Sci USA 1980;77:2163-2168. 74. Rassoulzadegan M, Binetruy B, Cuzin F. High frequency of gene transfer after fusion between bacteria and eukaryotic cells. Nature 1982;295:257- 259. 75. Gillies SD, Morrison SL, Oi VT, Tonegawa T. A tissue- specific transcription enhancer element is located in the major intron of a rearranged immunoglobulin heavy chain gene. Cell 1983;33:717-724. 76. Charnay P, Treisman R, Mellon P, Chao M, Axel R, Maniatis T. Differences in human alpha- and beta- globin gene expression in mouse erythroleukemia cells: The role of intragenic sequences. Cell 1984;38:251- 257. 77. Robert de Saint Vincent B, Delbruck S, Eckhart W, Meinkoth J, Vitto L, Wahl G. The cloning and reintroduction into animal cells of a functional CAD gene, a dominant amplifiable genetic marker. Cell 1981; 27:267-275. 78. Sandford JC, Klein TM, Wolf ED, Allen N. Delivery of substances into cells and tissues using a particle bombardment process. J Part Sci Technol 1987;5:27-37. 79. Sandford JC. The biollistic process. TIBTECH 1988;6:299-302. 80. Klein TM, Wolf ED, Wu R, Sanford JC. High velocity microprojectiles for delivering nucleic acids into living cells. Nature 1987;327:70-73. 81. Yang N-S, Christou, P. Particle bombardment technology for gene transfer. UWBC Biotechnical Resource Series. Oxford University Press, 1994. 82. Batty NP, Evans JM. Biological ballistics-no longer a shot in the dark. Transgenic Research 1992;1:107-113. 83. Yang N-S, Ziegelhofler PR. The particle bombardment system for mammalian transfer. In: Yang N-S, Christou P, editors. Particle bombardment technology for gene transfer. UWBC Biotechnical Resource Series. Oxford University Press, 1994:117-142. 84. Zelenin AV, Titomirov AV, Kolesnikov VA. Genetic transformation of mouse cultur ed cells with the help of high-velocity mechanical DNA injection. FEBS Lett 1989; 244:65-66. 85. Kolesnikov VA, Alimov AA, Barmintsev VA, Benyumov AO, Zelenina IA, Kraasnov AM, Dzhabur R, Zelenin AV. High velocity mechanical injection for foreign DNA into fish eggs. Genetika (Moskva) 1990;26:2122-2126. 86. Johnston SA. Biollistic transformation: microbes to mice. Nature 1990;346:776-777. 87. Yang N-S, Burkhelberg J, Roberts B, Martinell B, McCahe D. In vivo and in vitro gene transfer to mammalian somatic cells by particle bombardment. Proc Natl Acad Sci USA 1990;87:9568-9572. 88. Li C-Ch, Ruscetli FW, Rice NR, Chen E, Yang N-S, Mikovits J, Longo DL. Differential expression of Rel family members in human T-cell leukemia type-I infected cells. J Virol 1993;67:4205-4213. 89. Sanes JR, Rubinstein JLR, Nicolas JF. Use of a recombinant retrovirus to study post-implantation cell lineage in mouse embryos. EMBO J 1986;5:3133-3142. 90. de Went JR, Wood KV, de Luca M, Helinski DR, Subramani S. Firefly luciferase gene: Structure and expression in mammalian cells. Mol Cell Biol 1987;17:725-737. 91. Cheng L, Fu J, Tsukamoto A, Hawley R. Use of variants of the green fluorescent protein to monitor gene transfer and expression in mammalian cells. Nature Biotechnology 1996;14:606-609. 92. Cody CW, Prasher DC, Westler WM, Prendergast FG, Ward WW. Chemical structure of the hexapeptide chromophore of the Aequarea green fluorescent protein. Biochemistry 1993;32:1212-1218. 93. Delagrave S, Hawtin RE, Silva CM, Yang MM, Youvan DC. Red-shifted excitation mutants of the green fluorescent protein. Biotechnology 1995;13:151-154. 94. Heim R, Cubitt AB, Tsien RY. Improved green fluorescence. Nature 1995;373:663-664. 95. Anderson MT, Tjive IM, Lorincz MC, Parks DR, Herzenberg LA, Nolan GP, Herzenberg LA. Simultaneous fluorescence-activated cell sorter analysis of two distinct transcriptional elements within a single cell using engineered green fluorescent proteins. Proc Natl Acad Sci USA 1996;93:8508-8511. 96. Levy JP, Muldon RR, Zolotukhin S, Link Jr, CJ. Retroviral transfer and expression of a humanized red-shifted green fluorescent protein gene into human tumor cells. Nature Biotechnology 1996;14:610-614. 97. Ikama M, Kominami K, Yoshimura Y, Tanaka K, Nishimune Y, Okabe M. Green fluorescent protein as a marker in trans genic mice. Develop Growth Differ 1995; 37:455-459. Copyright 1997 Elfos Scientiae The following images related to this document are available:Photo images[ba97037b.jpg]Line drawing images[ba97037a.gif] |
| |||||||||

{kind=link}
{kind=link}