 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
GENETIC TRANSFORMATION OF SUGARCANE BY Agrobacterium tumefaciens USING ANTIOXIDANT COMPOUNDS Gil A Enriquez-Obregon, Roberto I Vazquez-Padron, Dmitri L Prieto-Samsonov, Marlene Perez and Guillermo Selman-Housein
Division Biotecnologia de las Plantas. Centro de Ingenieria Genetica y
Biotecnologia, apartado postal 6162, Ciudad de La Habana, CP 10600,
Cuba. Received in August 1996. Accepted for publication in December 1996.
Code Number:BA97039
Size of Files:
Text: 30.4K
Graphics: Line drawings (gif) - 7.5K
Photographs (jpg) - 108.9K
ABSTRACT The effects of 3 antioxidant compounds -ascorbic acid, cysteine and silver nitrate- on the growth of the Agrobacterium tumefaciens strain At 2260 and its interaction with plant cells, as well as on the viability and callus formation of meristematic tissue, were evaluated using explants from in vitro and field-cultured sugarcane plants commercial variety Ja 60-5. We achieved the transfer of a binary vector bearing the uid1 and bar genes to sugarcane meristematic explants, and obtained the generation of BASTA^R-resistant and GUS-positive calli. The use of an antioxidant mix caused an 80 % cell death decrease in respect to controls, and the callus quality was not affected in any of the culture phases. Key words: transgenic plants, plant-microorganism interaction, hypersensitivity, necrogenesis, BASTA^R, GUS RESUMEN Se evaluaron los efectos de tres compuestos antioxidantes - acido ascorbico, cisteina y nitrato de plata - sobre el crecimiento de Agrobacterium tumefaciens cepa At 2260 y la interaccion de este con los explantes vegetales, asi como sobre la viabilidad y la formacion de callos en tejidos meristematicos de cana de azucar variedad comercial Ja 60-5, provenientes de plantas in vitro y de campo. Se logro la transferencia de un vector binario con los genes uid1 y bar a explantes meristematicos de cana de azucar y se obtuvieron callos resistentes a BASTA^R y GUS-positivos. El empleo de la mezcla de antioxidantes redujo en un 80 % la muerte celular con relacion al control, y no altero la calidad del callo en ninguna de las fases de cultivo. Palabras claves: plantas transgenicas, interaccion planta- microorganismo, hipersensibilidad, necrogenesis, BASTA^R, GUS
Introduction Sugarcane (Saccharum officinarum L.) is one of the most extended crops in tropical and subtropical zones. The sugar industry and the production of certain chemicals such as furfural, dextranes and alcohol depend on this plant. By-products derived directly from sugarcane and its industrial processing represent a valuable alternative source for animal feeding and the cellulose industry. The application of biotechnology to sugarcane studies has a significant impact on agricultural yields and industrial production. During recent years, plant tissue culture, molecular biology and plant transformation have been developed (1). Recently, the first transgenic sugarcane plants resistant to stemborer (Diatraea saccharalis) were reported; revealing the possibilities of genetic engineering of this crop and offering new breeding possibilities (2). The DNA transfer to sugarcane cells has been a drawback in the genetic manipulation of these plant species. The plant transformation methodologies based on the naturally occurring Agrobacterium tumefaciens gene transfer system allowed many important crops to be genetically engineered. However, at first, they were mostly dicotyledonous plants due to the apparent stringency of the A. tumefaciens host range. Because of this, many important monocotyledoneous plants such as rice, wheat and sugarcane remained inaccessible to genetic engineering for a long time. For those cases, alternative direct transformation methods have been developed. In sugarcane, stable transformation by intact cell electroporation (3) and particle bombardment technology (4) were reported.
The development of efficient and reliable methods for the Agrobacterium-mediated gene transfer to monocotyledonous plants has represented the most important breakthrough of the last years in plant transformation technology. This approach has enabled the generation of transgenic plants in crops previously inaccessible for Agrobacterium-mediated transformation such as rice (5), maize (6) and banana (7).
In this paper we report the evidences of Agrobacterium-mediated transfer of foreign DNA to sugarcane meristematic explants of the cv Ja 60- 5. The conditions for precocultivation, cocultivation, callogenesis and selection of BASTA^R-resistant calli are described. The effects of several antioxidants on the sugarcane-Agrobacterium interaction were evaluated. The presence of the heterologous DNA in the transformed sugarcane tissue was verified by polymerase chain reaction (PCR) and histochemical GUS assay. Materials and Methods Plant material An aseptic culture from sugarcane cv Ja 60-5 was established according to Ho et al. (8) in an MS basal medium (9). The genetic transformation assays were performed with meristematic sections of sugarcane plant spindlers. The plant material was taken directly from the field and was sterilized in 1.5 % NaClO solution during 20 min. Table 1 shows the compositions of all the culture media used in this study. Table 1. Composition of the culture media for Agrobacterium- mediated genetic transformation of sugarcane cv Ja 60-5.
---------------------------------------------------------------------------
Culture Composition
medium
---------------------------------------------------------------------------
P+5 MS Salts, 1 mg/L nicotinic acid, 0.8 mg/L vitamin B1, 0.5 mg/L
vitamin B6, 100 mg/L myo-inositol, 20 g/L sucrose, 500 mg/L casein
hydrolysate and 5 mg/L 2,4 D.
P+5 AO P+5 supplemented with 15 mg/L ascorbic acid, 40 mg/L cysteine
and 2 mg/L silver nitrate.
P+^ AZ P+5^ supplemented with 60 g/L sucrose and 30 g/L glucose.
P+^ SEL MS Salts, 1 mg/L nicotinic acid, 0.8 mg/L vitamin B1, 0.5 mg/L
vitamin B6, 100 mg/L myo-inositol, 20 g/L sucrose, 5 mg/L 2.4 D,
500 mg/L claforan and 4 mg/L BASTA^R.
P- MS Salts, 1 mg/L nicotinic acid, 0.8 mg/L vitamin B1, 0.5 mg/L
vitamin B6, 100 mg/L myo-inositol and 20 g/L sucrose.
P - AO P- supplemented with 15 mg/L ascorbic acid, 40 mg/L cysteine
and 2 mg/L silver nitrate.
--------------------------------------------------------------------------
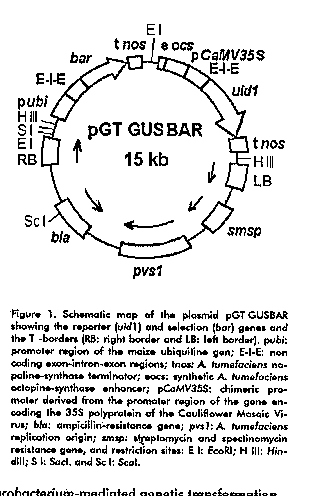
Bacterial strains and plasmids The A. tumefaciens C58C1Rifr: PGV2260 (At 2260) strain quoted by de la Riva et al. (10) was used for the establishment of the transformation methodology. The plasmid pGT GUSBAR containing the marker genes uid1 and bar under appropriate regulation signals for their expression in monocotyledoneous plants was constructed as follows: A 2.8 kb DNA fragment carrying the first non-coding exon-intron-exon sequence from the rice actin 1 gene, linked to the bar gene under the rice ubiquitin promoter regulation was obtained by EcoRI partial digestion of the plasmid pAHC-25 (11). This fragment was inserted in the EcoRI site of pBPFA5 (Coego A., personal communication). The resulting plasmid (pBPFA-GUSBAR), contained an expression cassette for the uid1 reporter gene (GUS activity), and the bar gene conferring resistance to the herbicide BASTA^R (Hoechst AG, Germany) fused to the first exon-intron-exon sequence from rice Actin 1 gene downstream of the CaMV 35S promoter. The described 6.1 kb expression cassette was separated by HindIII digestion from pBPFA-GUSBAR and transferred to the binary vector pGTDNA, a pDE1001 derivative in which the neomycin- resistance cassette was deleted. The final construct, named pGT GUSBAR (Figure 1), was inserted in A. tumefaciens by direct transformation (12).
The effects of antioxidant (AO) compounds on the growth of A. tumefaciens At 2260 were assessed by measuring the optical densities of the bacterial cultures at 260 nm after 24 h of cultivation in a YEB medium (10) supplemented with these compounds (Table 2). Table 2. Effects of antioxidant treatments on the A. tumefaciens growth, the meristematic tissue viability and the quality of the obtained calli.
-----------------------------------------------------------------------
Treatments Antioxidant Bacterial % explant Callus
compounds^1 OD620^2 viability^3 quality^4
-----------------------------------------------------------------------
AAS1 15 mg/L ascorbic acid 0.847 50 Type II
AAS2 30 mg/L ascorbic acid 0.803 50 Type III
CIS1 40 mg/L cysteine 0.570 60 Type II
CIS2 90 mg/L cysteine 0.598 80 Type III
Ag1 2 mg/L silver nitrate 0.490 70 Type II
Ag2 5 mg/L silver nitrate 0.010 60 Type III
P+5^5 none 0.600 10 Type II
AO 15 mg/L ascorbic acid;
40 mg/L cysteine;
2 mg/L silver nitrate 0.512 90 Type II
-----------------------------------------------------------------------
^1 Antioxidant compounds supplementing the liquid P+5 medium. ^2 Average of 3 independent determinations of OD620 in YEB A. tumefaciens cultures grown at 28 C for 20 h. ^3 Viability percentages represented as the averages from 10 independent stereoscopic determinations of the fraction of explant area remaining unstained after Evans Blue stain vs. total explant area. ^4 Callus qualities were assessed visually according to Ho et al. (8), and classified as follows: Type II. Friable yellow calli, with high capacity of embryogenesis. Type III. Soft, opaque, non-embryogenic white calli. ^5 P+5 medium without antioxidants used as negative control. --------------------------------------------------------------------------- Evaluation of the effects of antioxidants on explant viability and callogenesis The sugarcane meristematic explants (ca. 0.5 cm) were incubated in a P+5 medium (8), supplemented with the AO compounds for 60 h in the dark. The 8 treatments are described in Table 2. After exposure to the antioxidants, cell viability in the meristematic sections was evaluated using the Evans Blue method (13). Percentages of cell viability were determined as rates of unstained vs stained areas by direct stereoscopic observation of each treated ex-plant. Averages were calculated from 10 independent repetitions per treatment. The callogenic capacities of the antioxidant- treated explants were assessed in a solid P+5 medium and the generated calli were classified according to Ho et al. (8). Genetic transformation
A. tumefaciens At 2260 containing the binary plasmid pGT GUSBAR, was grown in a liquid medium supplemented with ampicillin 100 mg/L, spectinomycin, streptomycin 100 mg/L, rifampicin 50 mg/L and carbenicillin 50 mg/L. When optical density (OD) at 620 raised to 0.6, the cells were collected by centrifugation and resuspended in the same volume P+5 medium supplemented with antioxidants (P+5-AO). The same operations were carried out in other sample series adding 20 mg/L of acetosiringone to the final concentration. The effect of AO mixtures on the transfer of foreign genes from A. tumefaciens to sugarcane cells under 8 different treatments was evaluated (Table 3). The coculture was performed with explants from in vitro cultured plants and from field plants. After the explants were incubated in 10 mL of the P+5 or P+5-AO medium for 6-12 h in the dark, 1 mL of the A. tumefaciens culture was added and they were incubated for 10 min with strong agitation. The explants were then placed on filter paper and for cocultivation on a solid P+5 or P+5-AO medium for 3 days. Table 3. Effects of antioxidant compounds on the Agrobacterium- sugarcane gene transfer.
---------------------------------------------------------------------------
Treatments^1 Precoculture Coculture % GUS positive % BASTA^R- Analysis
medium medium explants^2 resistant by PCR^4
calli^3
---------------------------------------------------------------------------
IN 1 P-AO P+5 AO 100 15.5 (55) +
IN 2 P- P+5 AO 70 8.8 (55) +
IN 3 P-AO P+5 80 6.6 (55) +
IN 4 P- P+5 20 0 (55) na
PC 1 P- AO P+5 AO 100 26.5 (25) +
PC 2 P- P+5 AO 40 0 (25) na
PC 3 P-AO P+5 80 13.25 (25) +
PC 4 P- P+5 0 0 (25) na
---------------------------------------------------------------------------
^1 IN: explants from in vitro cultured sugarcane plants; PC:
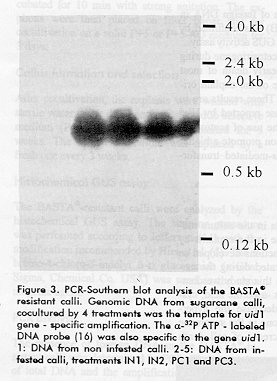
explants field-grown sugarcane plants.^^2 GUS activity was tested 2 days after co-cultivation, using a standard histochemical procedure for 10 samples randomly selected from each treatment. ^3 Total number of Agrobacterium infected explants appears between parentheses.^ ^4 PCR analysis performed with genomic DNA extracted from one callus per treatment; na: samples not available. -------------------------------------------------------------------------- Callus formation and selection After cocultivation, the explants were washed with sterile water, wipe dried and transferred to selective medium (P+5 SEL) where they remained for 5 weeks. The selective medium was replaced by a fresh one every 3 weeks. Histochemical GUS assay The BASTA^R-resistant calli were analyzed by the histochemical GUS assay. The staining procedure was performed according to Jefferson (14) with the modification recommended by Hiei et al. (5). The 5-bromo-4-chloro-3- indolyl _-d glucoronide (x-Gluc, Sigma, Chemical Co. USA) was used as the chromogenic agent. PCR analysis The presence of foreign DNA in the BASTA^R-resistant calli was determined by PCR. The extraction of total DNA and the amplification reactions with the synthetic primers gus A +350 (5' GCC ATT TGA AGC CGA TGT CAC GCC 3') and gus A +1400 (5' GTA TCG GTG TGA GCG TCG CAG AAC 3') were done as reported previously (15).The PCR products were analyzed by Southern blot using a 1.8 Kb DNA fragment containing the gene uid1 as a probe (16). Results Effect of different antioxidant compounds on necrogenesis of sugarcane meristems
To obtain sugarcane meristematic explants with low rates of necrogenesis in the cut surface zones and highly competent to transformation by A. tumefaciens, the effects of 3 AO compounds (ascorbic acid, cysteine and silver nitrate) were assessed during the precocultivation stage (Table 2). The fraction (percentage) of meristematic section areas remaining unstained by the Evans Blue reagent was taken as a cell viability criterion. Each compound was individually tested at 2 different concentrations in the liquid P+5 media. In all cases, a significant decrease in necrogenesis (less than 50 % of the explant areas) was observed after 60 h of incubation (Table 2). Although the hypersensitivity in each assayed treatment decreased, the best results were obtained when an AO mix was used. In this case, the meristematic explant necrosis was inhibited up to 90 % during the AO treatment. This result shows that the synergistic effects on the hypersensitive response propagation through the meristematic tissue among the AO agents used may exist. However, more detailed studies are needed to characterize the phenomena observed. After the pre-incubation step, the explants were placed in the dark for 5 weeks. The morphophysiological features of the formed calli were different in each case. For treatments AAS1, Cis 1, Ag 1 and AO, yellow and friable calli with high regeneration capacity (type II) similar to those obtained from the control (P+5) explants, were formed (Table 2). In all cases when higher levels of AO compounds were used, the calli resulted opaque and soft, with low regeneration capacity (type III). The negative action of silver nitrate on callus formation in potato cv Bintje and Desiree when the quality of the callus and its regeneration capacity were affected at a concentration of 10 mg/L, was described (17). In all the experiments concerning the effects of AO compounds on sugarcane callogenesis, there were no differences between the calli obtained from in vitro and field-grown sugarcane explants. All the treatments affected bacterial growth when compared with the controls, with the exception of Ag 2 which showed a negative effect. Selective agent To our knowledge, there were no previous reports on chemical selection of genetically transformed cells from the sugarcane cv Ja 60-5. The use of selective agents in transformation procedures is essential to avoid chimeric plants. For this reason we decided to determine the effectiveness of the herbicide BASTA^R -a compound reported to be cytotoxic in monocotyledonous plants- for the selection of transformed sugarcane calli from cv Ja 60-5. The meristems from in vitro cultured plants were maintained in the callus formation medium P+5, supplemented with BASTA^R at concentrations of 0, 1, 2, 3, 4 and 5 mg/L. After 2 weeks, the fraction of proliferated calli was determined. We observed that the meristematic tissues were sensitive to concentrations of 4 mg/L and higher (data not shown). Effect of antioxidants on plant tissue Agrobacterium interaction The effect of AO compounds on the interaction of A. tumefaciens with sugarcane meristematic tissues was studied using the A. tumefaciens At 2260 strain transformed with the binary plasmid pGT GUSBAR. This plasmid contains the genes uid1 and bar under the posttranscriptional regulation signals for monocotyledonous plants, which include introns. This fact made impossible the expression of marker genes in the bacterial host as was previously reported (18). We observed no blue staining when pGT GUSBAR-transformed A. tumefaciens samples were submitted to a histochemical assay similar to the one used to evaluate the GUS activity in sugarcane explants (data not shown). The AO treatment was selected to study the effect of AO compounds on gene transfer. The fractions of GUS positive and BASTA^R-resistant calli were taken as criteria for transformation efficiency using different treatments, assayed with explants from in vitro culture (IN) and from the field (PC) (Table 3).
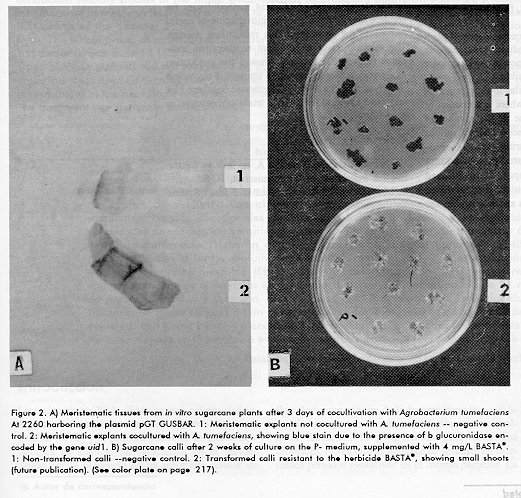
In all cases when the culture media were supplemented with antioxidants (IN1, PC1) during the pre and cocultivation, 100 % of infected explants became GUS positive (Figure 2A). In contrast, the infection efficiency was remarkably lower when antioxidants were not included. No GUS activity was found in infected controls.
The addition of 20 mg/L of acetosiringone during bacterial growth provoked the early death of most meristematic tissues irrespective of the explant origin and the treatments applied. These results are in apparent contradiction with those reported by Hiei et al. (5), who supported that the use of acetosiringone and high sugar concentration promote a higher efficiency in the A. tumefaciens-mediated transformation of rice. Discussion The inclusion of AO compounds in the culture media decreases the cell death rate in the explants by inhibiting the hypersensitive reactions developed as a response to the damage generated during the manipulation of the tissue (19). In this study we demonstrated that 3 AO compounds -ascorbic acid, cysteine and silver nitrate- can decrease the hypersensitivity reaction on the cut zone in the sugarcane meristematic explants. The decrease of cell death rates after cutting improved the competence of plant tissue to the Agrobacterium-mediated gene transfer. In the opposite cases, a fast hypersensitive response made the transformation impossible. The results obtained are similar to those reported in grape (Vitis vinifera) (19) where a reduction of callus necrosis and an increase in the number of transformation events were observed when various AO compounds were added during explants Agrobacterium interaction. These studies showed that the target of the AO compounds mentioned should be the oxygen reactive species produced by the extracellular peroxidases and involved in the propagation of the local hypersensitivity response in the plant. The host-spectrum of wild A. tumefaciens strains in natural conditions is limited mainly to dicotyledonous plants. Recently, the Agrobacterium-mediated transformation of several monocotyledonous plant species was successfully achieved under in vitro cell viability favoring conditions (5-7). We established those conditions for the transfer of foreign DNA to sugarcane meristematic explants. These explants were previously treated with AO compounds inhibiting cellular necrosis. The genetic transformation was performed and the GUS activity was histochemically detected in infected tissues as well as in regenerated calli. These calli also showed a remarkable resistance to BASTA^R. The transgenic material was positive when tested by PCR.
The explants from field plants showed better callogenesis in the P+5 SEL medium compared to those obtained from plants cultured in vitro. This fact is possibly related with the capacity of dedifferentiation and the physiological conditions of the meristems.
The results obtained are an important step in the whole extension of genetic engineering to sugarcane by the development of efficient methodologies for stable transformation and the introduction of useful traits into the genome of this recalcitrant species. The application of an efficient regeneration technique (Figure 2B) recently allowed the production of Agrobacterium-transformed transgenic sugarcane plants, as confirmed by PCR, genomic Southern blot and field herbicide-resistance trials (manuscript in preparation). Acknowledgments We acknowledge Dr. Gustavo A. de la Riva for the English translation of the manuscript; B. Sc. Ariel F. Martinez for style corrections and Jesus Seoane for the photographs. References 1. Martin JR, Galvez G, de Armas R, Espinosa R, Vigoa R, Leon A. La cana de azucar en Cuba. Editorial Cientifico-Tecnica. La Habana 1982. 2. Arencibia A, Vazquez RI, Prieto D, Carmona E, Tellez P, Coego A et al. Transgenic sugarcane (Sacharum spp.) plants are tolerant to stem borer attack despite the low expression levels of cryIA(b) gene from Bacillus thuringiensis var kurstaki HD1. Biotecnologia Aplicada 1996;13(2):130-131. 3. Arencibia A, Molina P, de la Riva G, Selman-Housein G. Production of transgenic sugarcane (Saccharum officinarum L.) plants by intact cell electroporation. Plant Cell Reports 1995;14: 305-309. 4. Bower R, Birch RG. Transgenic sugarcane plants via microproyectile bombardment. The Plant Journal 1992;2: 409-416. 5. Hiei Y, Ohta S, Komari T, Kumashiro T. Efficient transformation of rice (Oriza sativa) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. The Plant Journal 1994; 6(2): 271- 282. 6. Ishida Y, Saito H, Ohta S, Hiei Y, Komari T, Kumashiro T. High efficiency transformation of maize (Zea mayz L.) mediated by Agrobacterium tumefaciens. Nature Biotechnology 1996;14:745-750. 7. May GD, Afza R, Mason HS, Wiecko A, Novak FJ, Arntzen CJ. Generations of transgenic banana (Musa acuminata) plants via Agrobacterium mediated transformation. Bio/Technology 1995; 13:486-492. 8. Ho J, Vasil IK. Somatic embryogenesis in sugarcane (Saccharum officinarum). The Morphology and Physiology of callus formation and the ontogeny somatic embryos. Protoplasma 1983;118: 169-180. 9. Murashige T, Skoog F. A revised medium for rapid growth and bioassay with tobacco tissue culture. Physiol Plant 1962;15:473-479. 10. de la Riva G, Montagu MV, Inze D, Dhaese P. High-efficiency transformations of Agrobacterium tumefaciens with plasmid DNA by electroporation. Biotecnologia Aplicada 1992;8(2):345-351. 11. Taylor MG, Vasil V, Vasil IK. Enhanced GUS gene expression in cereal/grass cell suspensions and immature embryos using the maize ubiquity based plasmodia pAHC25. Plant Cell Reports 1993;12:491-495. 12. Holster M, de Wale D, Depicker A, Messens E, Van Montagu M, Shell J. Transfection and transformation of Agrobacterium tumefaciens. Mol Gen Genet 1978;163:181-187. 13. Evans DA, Sharp WR, Ammirato PV, Yamada Y. Handbook of Plant Cell Culture, Volume I published by Macmillan, New York 1983. 14. Jefferson RA. Assaying chimeric genes in plants: the gene fusion system. Plant Mol Biol Rep 1987;5:387-405. 15. Klimyut I, Bernard JC, Colwin MT, Jones DJ. Alkali treatment for rapid preparation of plant material for reliable PCR analysis. Plant Journal 1993;3(3):493-494. 16. Sambrook J, Fritsh EF, Maniatis. Molecular cloning. A laboratory Manual (Sec. Ed.). Cold Spring Harbor Laboratory Press, USA, 1989. 17. de Block M. Genotype-independent leaf disc transformation of potato (Solanum tuberosum L.) using Agrobacterium tumefaciens. Theor Appl Genet 1988;76:767-774. 18. Vancannyt G, Schmidt R, O Connor -Sanchez A, Willmitzer L and Rocha- Sosa M. Construction of an intron-containing marker gene: splicing of intron in transgenic plant and its use in Agrobacterium-mediated plant transformation. Mol Gen Genet 1990;220:245-250. 19. Perl A, Lotan O, Abu-Abied M, Holland D. Establishment of an Agrobacterium-mediated transformation system for grape (Vitis vinifera L.): The role of antioxidants during the grape- Agrobacterium interactions. Nature Biotechnology 1996;14:624-628. Copyright 1997 Elfos Scientiae The following images related to this document are available:Photo images[ba97039c.jpg] [ba97039b.jpg]Line drawing images[ba97039a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}