 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
IDENTIFICACION DE NUEVAS FUNCIONES BIOLOGICAS DEL EXTRACTO DIALIZABLE LEUCOCITARIO DE IMPACTO EN LA TERAPEUTICA DE ENFERMEDADES INFECCIOSAS E INFLAMATORIAS Celia Fernandez Ortega,^1 Manuel de J Arana,^1 Miriam Ojeda,^1 Marta Dubet,^2 Ignacio Ruibal,^2 Olga L Vilarubia,^2 Juan C Menendez de San Pedro^2 y Lourdes Navea^2 ^1 Centro de Investigaciones Biologicas, apartado postal 6996, La Habana, Cuba. ^2 Laboratorio Nacional de Referencia del SIDA, La Habana, Cuba.
Code Number:BA97046
Size of Files:
Text: 17.8K
Graphics: Line drawings (gif) -18.3K
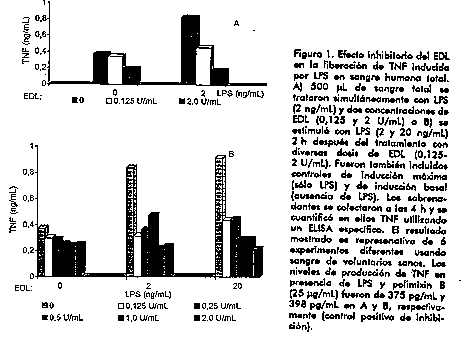
Introduccion En los ultimos anos, la inmunoestimulacion no especifica ha progresado desde el empleo de mezclas microbianas crudas y extractos hasta estrategias mas sofisticadas que utilizan una larga coleccion de compuestos farmacologicamente activos. El factor de transferencia (FT) es una preparacion con demostrados efectos sobre el sistema inmune. Es un material dializable de bajo peso molecular no inmunogenico, obtenido de leucocitos humanos de sangre periferica, que posee la capacidad de transferir inmunidad mediada por celulas (1). A pesar del empleo indistinto de los terminos FT y EDL, el FT define realmente a los componentes del EDL que median respuestas T de naturaleza antigeno especifica (2). Esta fraccion contiene multiples factores que parecen corresponder a la sumatoria de la experiencia inmune de un individuo particular. El EDL ha sido ampliamente utilizado en la practica clinica. Sin embargo, los mecanismos de accion farmacologica descritos respaldan solo parcialmente algunas de las respuestas terapeuticas observadas, especialmente, cuando su uso esta dirigido a entidades en las que desempenan un papel patologico mecanismos que no son corregibles con la potenciacion de la respuesta inmune afectada solamente. Este trabajo recoge la descripcion de dos nuevas actividades biologicas del extracto dializable leucocitario (EDL): la inhibicion de la produccion de factor de necrosis tumoral (TNF) por leucocitos humanos activados y la inhibicion de la replicacion del VIH. Estos efectos del EDL tienen particular relevancia por su repercusion terapeutica y su valor analitico. Resultados y Discusion Inhibicion de la produccion del TNF La produccion descontrolada o la superproduccion de citocinas constituye un mecanismo patogenico fundamental en respuestas inflamatorias sistemicas desencadenadas por el LPS de bacterias Gram negativas. Eventos tempranos asociados con respuestas perjudiciales a la endotoxina son atribuidos a una secrecion descontrolada de TNF por monocitos y macrofagos activados, asi como por otras celulas. Los monocitos humanos son extremadamente sensibles al LPS y niveles tan bajos como 10 pg/mL pueden inducir rapidamente la sintesis de citocinas inflamatorias como TNFalpha e interleuquina 1beta (3). Demostramos en nuestro estudio que el EDL no afecta la citotoxicidad mediada por TNF, pero si reduce significativamente la secrecion estimulada por LPS de la proteina biologicamente activa. El efecto inhibitorio del EDL se observo tanto en cultivos de celulas monociticas aisladas, como en experimentos con sangre total (Figura 1).
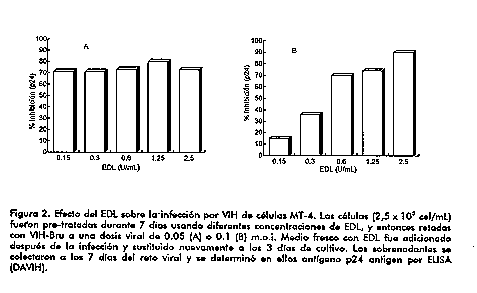
El extracto de leucocitos fue capaz de disminuir los niveles de TNF secretados y cuantificados al transcurrir un periodo de 5,5 h despues de iniciada la induccion con LPS en dosis elevadas. La inhibicion provocada por el EDL en ausencia de proteinas sericas no fue revertida por la presencia de concentraciones mas elevadas de LPS. Por consiguiente, la secrecion disminuida de TNF mediada por EDL no parece ser dependiente de moleculas competidoras con el LPS por sitios de reconocimiento especifico sobre la superficie celular. Es ampliamente aceptado que la activacion de leucocitos mediada por LPS ocurre principalmente como resultado de la interaccion de un complejo formado por la endotoxina y la proteina LBP (o el LPS acomplejado a otras proteinas) con la molecula de superficie CD14, que se expresa en monocitos, macrofagos (4) y leucocitos polimorfonucleares (3, 5). Mientras que esta via dependiente de CD14 es predominante a bajas concentraciones de LPS, en presencia de concentraciones elevadas de LPS y en ausencia de proteinas sericas accesorias, parece predominar una via independiente de CD14 (6). En nuestros experimentos, realizados en ausencia de suero, es probable que la activacion ocurra primariamente a traves del segundo mecanismo, aunque no deben descartarse eventos menores de estimulacion a traves de la interaccion directa entre CD14 y el LPS o, inclusive, la presencia de cantidades residuales de proteinas sericas accesorias absorbidas. De esta manera, nuestras observaciones relacionadas con una inhibicion significativa en condiciones de ausencia de suero, no parece involucrar mecanismos mediados por LBP, septinas o la molecula CD14 soluble. Es notable que el EDL adicionado al cultivo de monocitos 1 h despues de iniciada la estimulacion con LPS, inhibe la liberacion de TNF a niveles similares a aquellos obtenidos bajo condiciones de incubacion simultanea. Esta resulta una importante clave para la aplicacion terapeutica del EDL en procesos inflamatorios agudos y cronicos asociados a la liberacion de citocinas proinflamatorias. Nuestros resultados indican que el tratamiento con EDL afecta mas bien mecanismos postraduccionales dependientes de LPS, pero no debe descartarse la posible influencia del extracto sobre la regulacion de la transcripcion genica. Otros investigadores han reportado la inhibicion de la expresion de esta citoquina mediante el bloqueo de vias que involucren segundos mensajeros (7), las cuales no solo conducen a una rapida acumulacion de ARNm de TNFalpha, sino tambien a una acelerada produccion de TNFalpha (8). En nuestro trabajo evaluamos el efecto del EDL en ensayos con sangre total. El tratamiento simultaneo in vitro de los leucocitos humanos con EDL y LPS reduce potentemente la produccion de TNF, evaluada 4 h despues de iniciada la induccion con la endotoxina (Figura 1A). En efecto, la presencia de altas dosis de EDL bloquea la secrecion de TNF inducida por la endotoxina y solamente se observa un ligero incremento sobre los niveles basales de la citocina en leucocitos tratados con pequenas dosis del extracto. Esta potente respuesta inhibitoria, obtenida en un sistema que mimetiza condiciones fisiologicas, enfatiza el potencial terapeutico de este producto en enfermedades inflamatorias asociadas al LPS. La reduccion de la produccion de TNF en presencia de pequenas dosis de LPS, parece indicar que el tratamiento de las celulas con EDL afecta la secrecion de citocinas estimulada por LPS dependiente de CD14, o que un evento comun desencadenado por la interaccion del LPS con la molecula CD14 u otro receptor de superficie celular, son los blancos de la accion inhibitoria del EDL. El ensayo en sangre total, incluidas las celulas polimorfonucleares que producen TNF en respuesta al LPS (9), demuestra que el EDL afecta la respuesta de diferentes tipos celulares. Se ha demostrado que la produccion de TNF inducida por LPS tiene un papel importante en el sindrome septico. Dadas las observaciones realizadas en este estudio relacionadas con la capacidad del EDL de suprimir la secrecion de TNF inducida por LPS y de reducir substancialmente la produccion de TNF por celulas previamente activadas (Figura 1B), se sustenta la evaluacion terapeutica de EDL en sindromes de respuesta inflamatoria generalizada. En pacientes con sepsis, se han reportado concentraciones reducidas de proteina C reactiva luego del tratamiento con EDL (10). Esta respuesta in vivo puede ser dependiente del control de la secrecion de TNF. Los resultados de este estudio tambien tienen particular relevancia para el tratamiento de otras enfermedades en las cuales la secrecion de TNF, y de otras citocinas y mediadores inducidos por TNF, desempenan un papel patogenico, como son el asma bronquial, la artritis reumatoide, el SIDA y la esquistosomiasis. Vale destacar que estas entidades tambien cursan con defectos en la respuesta inmune que pueden ser corregibles bajo los efectos inmunomoduladores del EDL. Inhibicion de la replicacion del VIH Desde el descubrimiento del VIH como agente causal del SIDA, han sido descritos diferentes compuestos con actividad anti-retroviral in vitro y alguno de ellos son comunmente utilizados en el tratamiento de la infeccion por VIH. Estas terapias anti-retrovirales ofrecen diferentes beneficios clinicos, como la disminucion de la velocidad de aparicion de infecciones oportunistas y prolonga la sobrevida, pero estos agentes son incapaces de detener la progresion de la infeccion viral. La mejoria inmunologica lograda es incompleta, y por otra parte, algunos de los actuales agentes terapeuticos son toxicos para las celulas del sistema inmune (11). Recientemente, la inmunoterapia ha prestado especial atencion a la busqueda de estrategias dirigidas contra el VIH. La terapia de combinacion que involucra el uso de agentes anti-retrovirales y compuestos inmunoterapeuticos parece ser prometedora. En este estudio, se demostro la capacidad del EDL de reducir la produccion viral en dependencia de la dosis en un modelo in vitro de infeccion por VIH. Este efecto se observo al tratar el cultivo celular con EDL durante 7 dias antes del enfrentamiento con el virus (Figura 2). No se modifico la produccion del VIH transcurrido un periodo de tratamiento breve (3 h) con el extracto.
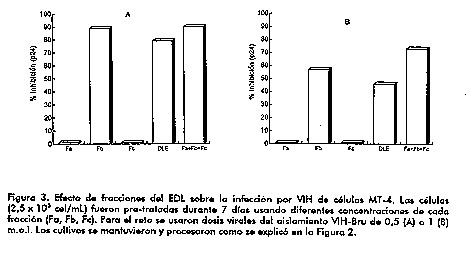
Probablemente, se requieren efectos acumulativos o prolongados para reducir la sensibilidad de las celulas a la infeccion por VIH o para lograr una afectacion parcial de la expresion viral. Deben explorarse posibles mecanismos relacionados con la actividad moduladora del EDL sobre la expresion de diferentes citocinas. Otros estudios refieren la participacion de redes de citocinas en el control de la expresion del VIH (12-14). En este estudio, se utilizo como modelo experimental la linea celular MT-4, celulas linfoblastoides CD4^+ infectadas con HTLV-1 altamente sensibles al VIH (15). Se ha reportado la produccion endogena de TNFalpha por celulas linfoblastoides y una reduccion de la sensibilidad de estas celulas a la infeccion por VIH al ser tratadas con anticuerpos especificos anti-TNFalpha (16, 17). La capacidad que tiene el EDL de reducir la produccion de TNF en monocitos activados con LPS, probablemente ejerce su efecto a nivel postranscripcional. A pesar de las diferencias entre ambos modelos experimentales, el EDL podria mediar efectos inhibitorios similares sobre la liberacion de TNF por la linea celular MT-4, que afectan indirectamente la expresion del VIH. Por otra parte, deben evaluarse otros efectos moduladores de este extracto sobre distintas citocinas reguladoras de la expresion del VIH, ademas de sobre eventos a nivel membranario que median la infeccion viral.
Estos resultados, obtenidos en sistemas in vitro utilizando una linea celular, indudablemente contribuyen a dilucidar los mecanismos involucrados en las interacciones celula-virus y permiten establecer un enfoque racional del uso terapeutico del EDL en la infeccion por VIH. Se ha demostrado una mejoria de la respuesta inmune en pacientes tratados con EDL (18). Esos resultados clinicos, conjuntamente con los del presente estudio, indican la regulacion por el EDL de la replicacion viral en un sistema in vitro y reafirman la importancia de continuar los estudios de caracterizacion del EDL y de instaurar su empleo de modo definitivo en la busqueda de una terapeutica efectiva del SIDA. References 1. Lawrence HS. Adv Immunol 1969;11: 195-266. 2. Fudenberg HH, Fudenberg HH. (1989) 557-626. En: Res. & Applic. Transfer Factor and DLE. Baolai, H. Ed. Xue Yuan Press, Beijing, China. 3. Zhong WW, Burke PA, Hand AT, Walsh MJ, Hughes LA, Forse RA. Regulation of cytokine mRNA expression in lipopolysaccharide-stimulated human macrophages. Arch Surg 1993;128:158-164. 4. Lynn WA, Liu Y, Golenbock DT. Neither CD14 nor serum is absolutely necessary for activation of mononuclear phagocytes by bacterial lipopolysaccharide. Infect Immun 1993;61:4452-4461. 5. Lynn WA, Raetz CRH, Qureshi N, Golenbock DT. LPS induced stimulation of CD11b/CD18 expression on neutrophils: evidence of specific receptor-based response and inhibition by lipid A-based antagonists. J Immunol 1991; 147:3072-3079. 6. Strieter RM, Kunkel SL, Bone G. Role of tumor necrosis factor-alpha in desease states and inflammation. Crit Care Med 1993;21:447-463. 7. Kovacs EJ, Radzioch D, Young HA, Varesio L. Differential inhibition of IL-1 and TNFalpha mRNA expression by agents which block second messenger pathways in murine macrophages. J Immunol 1988;141:3101-3105. 8. Gong JH, Renz H, Sprenger H, Nain M, Gemsa D. Enhancement of tumor necrosis factor-alpha gene expression by low doses of prostaglandin E2 and cyclic GMP. Immunobiol 1990;182:44-55. 9. Haziot A, Tsuberi BZ, Goyert SM. Neutrophil CD14: Biochemical properties and role in the secretion of tumor necrosis factor-alpha in response to lipopolysaccharide. J Immunol 1993;150: 5556-5565. 10. Lokaj J, Pekarek J, Kuklinek P. Dialysable leukocyte extract in the therapy of sepsis. Leukocyte dialysates and transfer factor. Slovak Academy of Science. Bratislava 1987;516-525. 11. Yarchoan R, Mitsuya H, Broder S. Challenges in the therapy of HIV infection. Immunol Today 1993;14:303-308. 12. Klesius P, McMeeking A, Borkowsky W, Bonk S, Holzman RS, Lawrence HS. Immunotherapy of cryptosporidium parvum with bovine Transfer Factor. In: Baolai H, Ruzhang W, Zhaofen Z, eds. Research and applications of Transfer Factor and DLE. Beijing: Xue Yuan Press, 1989;227-238. 13. Butera ST, Roberts BD, Folks TM. Regulation of HIV-1 expression by cytokine networks in a CD4+ model of chronic infection. J Immunol 1993;150: 625-34. 14. Poli G, Fauci AS. The effect of cytokines and pharmacologic agents on chronic HIV infection. AIDS Res Human Retro 1992;8:191-96. 15. Harada S, Koyanagi Y, Yamamoto N. Infection of HTLV-III/ LAV in HTLV-1- carrying cells MT-2 and MT-4 and application in a plaque assay. Science 1985;229:563-66. 16. Ito M, Baba M, Sato A, Hirabayashi K, Tanabe F, Shigeta S, DeClercq E. Tumor necrosis factor enhances replication of human immunodeficiency virus in vitro. Biochem Biophysic Res Commun 1989;158:307-12. 17. Kobayashi N, Hamamoto Y, Yamamoto N. Production of tumor necrosis factors by T cell lines infected with HTLV-1 may cause their high susceptibility to human immunodeficiency virus infection. Med Microbiol Immunol Berl 1990;179:115-22. 18. Rivero J, Limonta M, Gonzalez A, Ramirez V, Aguilera A, Lopez Saura P. Tratamiento con factor de transferencia en portadores del virus de la inmunodeficiencia humana. Biotecnologia Aplicada 1993;10: 2-3. Copyright 1997 Elfos Scientiae The following images related to this document are available:Line drawing images[ba97046b.gif] [ba97046c.gif] [ba97046a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}