 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Necessary redundancy: regulation and role of type I interferon system during embryogenesis Jose de la Fuente, *Eileen Riego, Fidel O Castro and Ricardo Lleonart Mammalian Cell Genetics Division. Center for Genetic Engineering and Biotechnology. PO Box 6162, Havana 6, Cuba. E-mail: clon@cigb.edu.cu Received in March, 1997. Accepted for publication July, 1997.
Code Number:BA97051
Size of Files:
Text: 26K
Graphics: Line drawings (gif) - 23.7K
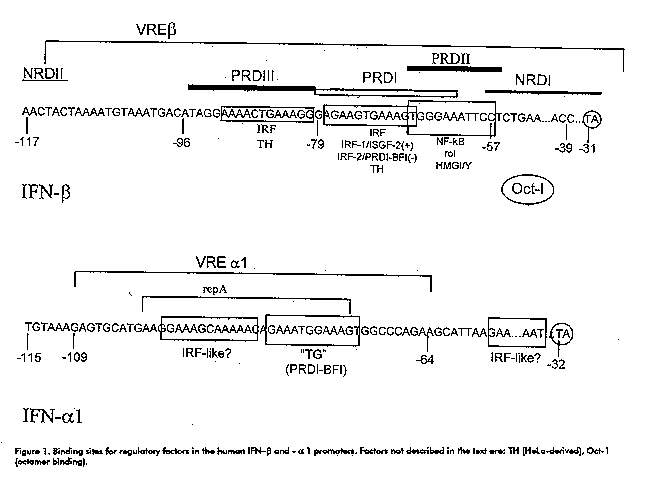
ABSTRACT Type I interferon (IFN) system is involved in the defense of the organism against viral infections and plays an important role in the control of cell growth and differentiation. How this system is regulated during embryogenesis and its role during this critical process are questions addressed in this manuscript. The redundancy, apparently existing in type I IFN system could be the result of a very specialized machinery needed to control the complex process of cell growth and differentiation during development. Key words: interferon, IRF, transgenic, mice, embryo RESUMEN El sistema interferon (IFN) tipo I participa en la defensa del organismo contra las infecciones virales y juega un papel importante en el control del crecimiento y la diferenciacion celular. Como esta regulado este sistema durante la embriogenesis y cual es su papel durante este critico proceso son preguntas que se tratan en este manuscrito. La aparente redundancia que existe en el sistema IFN tipo I pudiera ser el resultado de una maquinaria altamente especializada necesaria para controlar los complejos procesos involucrados en el crecimiento y la diferenciacion celular durante el desarrollo. Palabras claves: interferon, IRF, transgenico, raton, embrion Introduction Interferons Much of the development of Genetic Engineering and modern Biotechnology in Cuba, and especially at the Center for Genetic Engineering and Biotechnology (CIGB), has been related to interferons (IFNs). Promised as magic bullets when they were first discovered in 1957 (1) and cloned in 1979-1980 (2, 3), type I IFNs (IFN-alpha and IFN-beta) were useful in the treatment of only certain diseases (4), but constituted a starting point for molecular biologists and biotechnologists and a challenging model for the study of gene regulation. Type I IFNs are inducible, pleiotropic cytokines that are characterized by their ability to induce an antiviral state against a variety of viruses, they have been shown to have antiproliferative and antitumoral properties and to play a role in cell growth and differentiation (5, 6). In humans and rodents, type I IFNs are encoded by a superfamily of genes consisting of the IFN-alpha gene family and the IFN-beta gene which exists in a single copy.
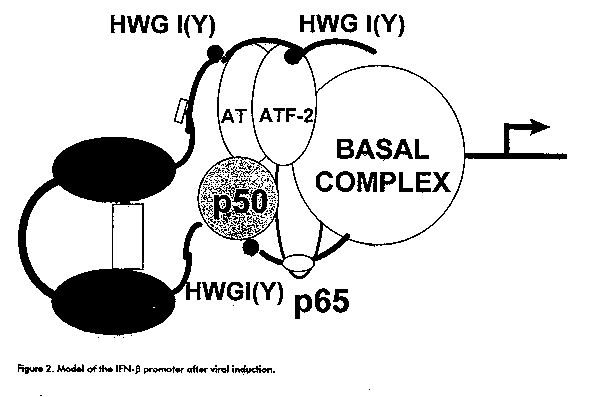
The IFN-alpha group has recently been subdivided into subfamilies: IFN-alphaI, IFN-alpha or IFN-omega and IFN-tau to which ruminant trophoblastic IFNs belong (7). Type I IFNs are intronless genes, tightly regulated by regulatory factors that interact with the 5' flanking region, although for the IFN-beta gene, sequences located in the 3' untranslated region may contribute to the regulation of the levels of the cognate protein by modulating mRNA turnover. Gene regulation The human IFN-beta promoter is regulated by at least 4 sequence elements contained within a 125 bp segment preceding the cap site (8, 9, Figure 1). The positive regulatory domain (PRD) II is a binding site for the activating factor NF-kappaB and for the high mobility group (HMG) Y/I protein which interacts with the minor groove of the DNA. Both factors are necessary for the viral induction of the IFN-beta promoter. PRDI and PRDIII bind the activating IFN regulatory factor 1 (IRF-1). PRDIV binds a protein of the cAMP response element binding protein (ATF/CREB) family of transcription factors (ATF-2), which is also required for full activation of the IFN-beta promoter. PRDI also binds negative regulatory factors IRF- 2, the zinc-finger protein PRD-binding factor 1 (PRD-BF1) and others. PRDII and two negative regulatory domains (NRDI and NRD II) are also believed to bind negative regulatory factors. IFN-beta gene transcription activation by a virus is achieved by derepression as well as by the activation of preexisting NF-kappaB and by the de novo synthesis and activity enhancement of IRF-1 (Figure 2). However, although IRF-1 is efficiently induced by virus, IFNs-gamma and -alpha and other cytokines, the expression of the IFN-beta gene is highly inducible only by virus. Mice devoid of functional IRF-1 show unimpaired type I IFN induction and response of type I IFN inducible genes in vivo by a virus or dsRNA (10, 11). These facts indicate that IRF-1 is not enough for IFN-beta induction and is dispensable for type I IFN induction in vivo.
Figure 2. Model of the IFN-beta promoter after viral induction.
IFNs are usually not detectable in normally growing tissue cultures or animals until appropriate stimuli trigger the induction of the IFN system. However, constitutive IFN expression has been reported in organs from normal individuals (18-20), peripheral blood leukocytes (PBL), where IFN- alpha could act in an autocrine fashion (21), preimplantation mouse embryos and embryonal carcinoma (EC) P19 cells (22), 13 day post coitum (dpc) mouse embryos (23), mouse blastocysts (24, 25) and in ruminant trophoblasts (7). Recently, a mouse IFN-alpha 11 gene has been identified with a Newcastle Disease Virus (NDV)-uninducible expression pattern (26). Expression of type I interferon system during embryogenesis Employing the reverse transcriptase-polymerase chain reaction (RT-PCR) method, the presence of mRNAs has been shown for TGF-alpha, -beta1, PDGF-A, IL-1-7, TNF-alpha, IRF-1 and IFN-gamma and IFN-alpha in preimplantation mouse embryos (22, 27), for IRF-1 and IRF-2 and IFN-alpha in undifferentiated (D^-) and differentiated (D^+) EC P19 cells, equivalent to the epiblast cells of the early postimplantation blastocyst (4.5 dpc) (22). Furthermore, preimplantation embryos possess the basic apparatus required to respond to polypeptide growth factors that could serve to regulate their growth in an autocrine fashion (28).
It has been shown that the type I IFN system is developmentally regulated in EC cells as well as in normal embryonic cells (17, 23). The IFN genes are refractory to viral induction in their undifferentiated stages and Harada et al. (17) have shown that this effect could be due, at least in part, to the developmental regulation of the transcription factors IRF-1 and IRF-2. In IRF-1^+/+ and IRF-1^0/0 murine embryonic stem (ES) D3 cells, Ruffner et al. (29) found that the viral induction of the IFN-alpha genes is not dependent on differentiation, while IFN-beta transcripts were barely detectable in (D^-) ES cells and increased about 10 fold after 8 days of differentiation. In EC P19 cells we (22) and others (30) have found that IFN-beta transcripts appear only after 24 h of differentiation with retinoic acid. The complete silencing of the IFN-beta gene in (D^-) cells might be due to a more efficient association of repressors to the 5' regulatory sequences and/or to the presence in these cells of other repressor molecules interacting with the upstream sequences in the IFN-beta promoter (31). Alternatively, the absence of some positive regulatory factors could also be implicated in silencing the IFN-beta gene in these cells. The analysis of the IFN-alpha genes constitutively expressed in (D^-) EC P19 cells resulted in the cloning of MuIFN-alphaAlpha and a new MuIFN-alpha gene (MuIFN-alpha 12) (22, Figure 4). The IFN-alpha A virus responsive element (VRE) in the promoter region showed the presence of nucleotides that differ from those present in mouse virus-inducible (-alpha 4) and uninducible (-alpha 11) IFN genes and in the Hu IFN-alpha 1 (Figure 5). Further characterization of IFN-alphaA will permit the study of the regulation of its constitutive expression in early mouse embryos. Figure 4. Sequence of the fragments cloned for Mu IFN- alphaA and - alpha-12. Amino acid numbers correspond to Mu IFN-alpha A (31).
---------------------------------------------------------------------------
aminoacid 15
Leu Asn Ser Leu Ala Gln Met Arg Arg Leu Ser Pro Leu Ser Cys
MuIFN-alpha12 --TTG AAC TCT CTG GCA CAA ATG AGG AGA CTC TCC CCT CTC TCC TGC
MuIfn-alphaA --*** *C* CTC *** *T* *** *** *** *** *** *** *** *** *** ***
Thr Leu Val
Leu Lys Asp Arg Lys Asn Phe Arg Phe Pro Gln Glu Lys Val Asp Ala Gln Gln
CTG AAG GAC AGA AAG AAC TTT AGA TTC CCC CAG GAG AAG GTG GAT GCC CAG CAG
*** *** *** *** *** G** *** *** *** *** *** *** *** *** *** *** *** ***
Asp
Ile Lys Lys Ala Gln Val Ile Pro Val Leu Ser Glu Leu Thr Gln Gln Ile Leu
ATC AAG AAG GCT CAA GTC ATC CCT GTC CTG AGT GAG CTG ACC CAG CAG ATC CTG
*** C** **T *** *** *C* *** *** *** **A CAA *** *** *** *** *** G** ***
Gln Asn Ala Gln Val
Thr Leu Phe Thr Ser Lys Asp Ser Ser Ala Gly Asp Ser Ser Ala Ala Trp Tyr
ACC CTC TTC ACA TCA AAG GAC TCA TCT GCT caa gac tca tct gct gca tgg tac
*A* A** *** *** *** *** *** *** *** *** --- --- G*T *GG *A* *** *CC ---
Asn Ile Ala Trp Asp Ser
Thr Leu Leu Asp Ser Phe Cys Asn Asp Leu His Gln Gln Leu Asn Asp Gln Gly
acc CTC CTA GAC TCA TTC TGC AAT GAC CTC CAT CAG CAG CTC AAT GAC CTG CAA
--- *** *** *** *** *** *** *** *** *** *** *** *** *** *** *** **C A**
Leu Lys
Ala Cys Leu Met Gln Gln Val Gly Val Gln Glu Pro Pro Leu Thr Gln Glu Asp
GGC TGT CTG ATG CAG CAG GTG GGG GTG CAG GAG CCT CCC CTG ACC CAG GAA GAC
*C* *** G** *** *** G** *** *** *** *** **A *** *** *** *** *** *** ***
Val Glu
Ser Leu Leu Thr Val Arg Lys Tyr Phe His Arg Ile Thr Val Tyr Lys Arg Glu
TCC CTG CTA ACT GTG AGG AAA TAC TTC CAC AGG ATC ACT GTG TAC CTG AGA GAG
*A* *** **G G** *** *** *C* *** *** *** *** *** G** *** *** *C* *** ***
Tyr Ala Thr Leu
149
Lys Lys His Ser Pro Cys Ala Trp Glu Val Val Arg Ser Glu Val Trp
AAG AAA CAC AGC CCC TGT GCC TGG GAG GTG GTC AGA TCA GAA GTC TGG
*** *** *** *** *** *** *** *** *** *** *** *** G** *** *** ***
Ala
---------------------------------------------------------------------------
Figure 5. Comparison of the nucleotide sequence of the VRE alpha A with other VRE in virus-inducible and not-inducible IFN genes.
---------------------------------------------------------------------------
VRE
-109 ---------------------------------------- -64
HuIFN-alfa1 TAAAGAGTGCATGAAGGAAAGCAAAAACAGAAATGGAAAGTGGCCCAGAAGCATTAA
MuIFN-alfa4 CAGAGAGTGAAGTAAAGAAAGTGAAAAGAGAATTGGAAAGCAAGGGGAGGCTATTCC
MuIFN-alfa11 CAGAGAGTGAAGTAAAGAAAGTGAAAAGAGAATTGGGAAGCAAGGGGAGGCTATTTA
MuIFN-alfaA CAGAGAATGAGTTAAAGAAAGTGAAAAGACAAGTGGAAAGAGAGAGGAGGACATTTC
* *** ** ** ***** **** * * * *** *** ***
MuIFN-alfa4 GAAAGGAGAAACTTCTATTTGTCCCTATTTAA
MuIFN-alfa11 CAAAGGAGAAACTTGTGTTTGTCCCTATTTAA
MuIFN-alfaA AGAAATGAAAACTAGTGTTTG-CCCTATTTAA
** ***** * **** **********
---------------------------------------------------------------------------
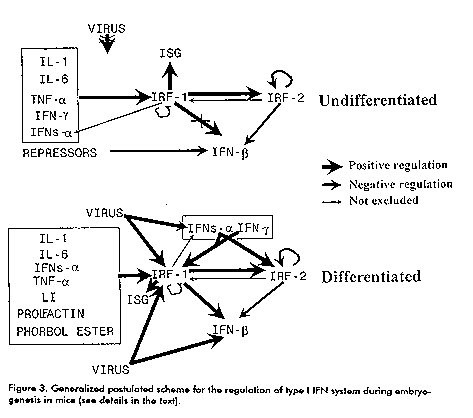
Nevertheless, from the present knowledge, we can propose a model for type I IFN system regulation during embryogenesis (Figure 3). Different regulatory pathways may lead to the constitutive expression of particular IFN-alpha and cytokine genes in (D^-) cells, that result in the induction of IRFs, other regulatory factors and IFN-stimulated genes (ISG). The IFN-beta gene remains repressed in these cells. After differentiation, the virus- inducible machinery becomes active and other IFN genes become inducible. Role of type I interferon system during embryogenesis The presence of uninduced mRNAs for IRF-1 and IRF-2, IFN-alpha and other cytokines in early mouse embryos suggest a role for type I IFN system during embryogenesis (22). Although we do not know the biological role of these constitutively expressed MuIFN-alpha A and -alpha12 genes, we could speculate on their role in the process of embryogenesis because of the known effects of IFNs in the development of the cytoskeleton and extracellular matrix and its capacity to decrease cell growth (6). These constitutive IFNs might also be involved in the protection of embryos against viral infections and the immune reaction of the host, thus playing a role in immune tolerance. It has been reported that mice devoid of the IFN-alpha/beta receptor have no evident problems during embryonic development (32). Although this result may contradict the possible role for IFN-alpha in the process of embryogenesis, it is not the first time that in ^-/^- mutant mice we see no obvious phenotype, suggesting that alternative pathways may exist. In some cases, the role of specific genes could be partially understood by overexpressing this gene, keeping in mind that it does not reflect physiological situations. The overexpression of MuIFN-alpha 1 in transgenic mice testes resulted in the degeneration of spermatogenic cells and sterility (33). For IRFs, they have been implicated in the regulation of cell growth and proliferation. As we have found (22), the expression of IRF-1 after the first cell cleavage could contribute to the control of cell growth in the developing embryo. As stated earlier, restrained cell growth depends on a balance between IRF-1 and IRF-2, that is also present during early stages of development (22). Again, for IRF genes, only a slightly deviating phenotype has been reported in IRF-deficient mice (10-11). We have generated transgenic mice with chimeric genes containing the human IRF-1 or IRF-2 cDNAs under the control of strong constitutive chicken beta- actin (pbActneoIRF-1 and pbActneoIRF-2) or human cytomegalovirus (pCMVIRF- 1) promoters (34). These constructs were shown to direct the synthesis of IRF-1 or IRF-2 mRNAs in human and/or mouse cell lines. Preliminary results indicated that the efficiency for the generation of IRF-1 transgenic individuals was lower (p < 0.05) than that for IRF-2 transgenic individuals and for a control group including 37 transgenic mice generated with different transgenes (35, Table 1). Because of the antiproliferative properties of IRF-1 in its role in the control of cell growth, we speculate that the high level ubiquitous constitutive expression of IRF-1 could be deleterious for the developing mouse embryo. Current experiments are directed to further characterize these transgenic mice. Table 1. Efficiency of the generation of IRF-1, IRF-2 and non-related transgenic individuals (control).
---------------------------------------------------------------------------
Index^a Transgenic individuals
--------------------------------
IRF-1 IRF-2 Control
---------------------------------------------------------------------------
Transgenic individuals/microinjected embryos 0.0026* 0.0217 0.045
Transgenic individuals/transferred embryos 0.0059* 0.0598 0.098
Transgenic individuals/born pups 0.0206* 0.3030 0.230
---------------------------------------------------------------------------
^a: The number of microinjected and transferred embryos and of pups born
were similar in the three groups; *: different (p < 0.05) by a Student T
test for non-paired samples with Levene's test for equality of variances
supported by SPSS5.01 for Windows (SPSS Inc, USA).Conclusion Necessary redundancy
The fact that IFN-alpha consists of a clustered multigene family and that individual genes are differentially expressed in a cell type-specific manner through probably different induction pathways, lead to the possibility that there is a differential expression of the IFN genes which is related to a given function in a given specific tissue, for a precise time, and at a particular moment of the cell physiology and developmental stage (Figure 6). This study could possibly contribute to the understanding of the highly conserved structure and function of the IFN-alpha gene family.
We want to thank Drs. T.Taniguchi and H. Harada for providing us with the pbActneoIRF plasmids and Dr. S. Pestka for the MuIFN-alpha A clone. Part of this work was supported by a Matsumae International Foundation Research Fellowship to J. F. References 1. Issacs A, Llindenmann J. Virus Interference. I. The Interferons. Proc Royal Soc Lond 1957;B147:258-267. 2. Taniguchi T, Sakai M, Fujii-Kuriyama Y, Muramatsu M, Kobayashi S, Sudo T. Construction and identification of a bacterial plasmid containing the human fibroblast interferon gene sequence. Proc Japan Acad 1979;Ser B55:464-469. 3. Nagata S, Mantei N, Weissmann C. The structure of eight or more distinct chromosomal genes for human interferon-alpha. Nature 1980;287:401-408. 4. Lopez-Saura P. What is interferon good for? Ten years of experience in Cuba. Biotecnologia Aplicada 1992;9:207-227. 5. Weissmann C, Weber H. The interferon genes. Prog Nucleic Acid Res Mol Biol 1986;33:251-300. 6. Vilcek J. Interferons. In: Peptide growth factors and their receptors. Handbook of experimental pharmacology. In: MA Sporn, AB Roberts, (eds);New York: Springer-Verlag, 1990;3-38. 7. Imakawa K, Hansen TR, Malathy P-V, Anthony RV, Polites HG, Marotti KR, Roberts RM. Molecular cloning and characterization of complementary deoxyribonucleic acids corresponding to bovine trophoblast protein-1: A comparison with ovine trophoblast protein-1 and bovine interferon -alphaII. Mol Endocrinol 1989;3:127-139. 8. Taniguchi T. In: The biology of the interferon system (Kawade and Kobayashi eds.). Tokyo 1990;3-10. 9. Maniatis T. In: Transcriptional regulation, Part 2 (McKnight and Yamamoto eds.). CSH 1992;1193-1220. 10. Matsuyama T, Kimura T, Kitagawa M, Pfeffer K, Kawakami T, Watanabe N et al. Targeted disruption of IRF-1 or IRF-2 results in abnormal type I IFN gene induction and aberrant lymphocyte development. Cell 1993;75:83-97. 11. Reis LFL, Ruffner H, Stark G, Aguet M, Weissman C. Mice devoid of interferon regulatory factor 1 (IRF-1) show normal expression of type I interferon genes. The EMBO J 1994;13:4798-4806. 12. Harada H, Takashi E-I, Itoh S, Harada K, Hori T-A, Taniguchi T. Structure and regulation of the human interferon regulatory factor 1 (IRF- 1) and IRF-2 Genes: Implications for a Network in the Interferon System. Mol Cell Biol 1994;14:1500-1509. 13. Willman CL, Sever CE, Pallavicini MG, Harada H, Tanaka N, Slovak ML et al. Deletion of IRF-1, mapping to chromosome 5q31.1, in human leukemia and preleukemia myelodysplasia. Science 1993;259:968-971. 14. Harada H, Kitagawa M, Tanaka N, Yamamoto H, Harada K, Ishihara M, Taniguchi T. Anti-oncogenic and oncogenic potential of interferon regulatory factors -1 and -2. Science 1993;259:971-974. 15. Tanaka N, Ishihara M, Kitagawa M, Harada H, Kimura T, Matsuyama T et al. Cellular commitment to oncogene-induced transformation or apoptosis is dependent on the transcription factor IRF-1. Cell 1994;77:829-839. 16. Mac Donald N, Kuhl D, Maguire D, Naf D, Gallant P, Goswamy A et al. Different pathways mediate virus inducibility of the human IFN- alpha1 and IFN-alpha genes. Cell 1990;60:767-779. 17. Harada H, Willison K, Sakakibara J, Miyamoto M, Fujita T, Taniguchi T. Absence of the type I interferon system in EC cells: Transcriptional activator (IRF-1) and (IRF-2) genes are developmentally regulated. Cell 1990;63:303-312. 18. Tovey MG, Streuli M, Gresser I, Gugenheim J, Blanchard B, Guymarho J et al. Interferon messenger RNA is produced constitutively in the organs for normal individuals. Proc Natl Acad Sci USA 1987;84:5038-5042. 19. Khan NUD, Pulford KAF, Farquharson MA, Howatson A, Stewart C, Jackson R et al. The distribution of immunoreactive interferon-alpha in normal human tissues. Immunology 1989;66:201. 20. Khan NUD, Gibson A, Foulis AK. The distribution of immunoreactive interferon-alpha in formalin-fixed paraffin embedded normal human fetal and infant tissues. Immunology 1990;71:230. 21. Greenway AL, Brandt ER, Hertzog PJ, Devenish RJ, Linnane AW. Post- transcriptional regulation of interferon-alpha 4 subtype production by lymphoblastoid cells. Hematol Oncology 1993;23:229-235. 22. Riego E, Perez A, Martinez R, Castro FO, Lleonart R, de la Fuente J. Differential constitutive expression of interferon genes in early mouse embryos. Mol Reprod Dev 1995;41:157-166. 23. Barlow DP, Randle BJ, Burke DC. Interferon synthesis in the early post- implantation mouse embryos. Differentiation 1984;27: 299-235. 24. Cross JC, Farin CE, Sharif SF, Roberts RM. Characterization of the antiviral activity constitutively produced by murine conceptuses: Absence of placental mRNAs for interferon alpha and beta. Mol Reprod Dev 1990;bn 26:122-128. 25. Nieder GL. Protein secretion by the mouse trophoblast during attachment and outgrowth in vitro. Biol Reprod 1990;43:251-259. 26. Civas A, Dion M, Vodjdani G, Doly J. Repression of the murine interferon alpha 11 gene: identification of negatively acting sequences. Nucl Acid Res 1990;19:4497-4502. 27. Rothsteins JL, Johnson D, Delogia JA, Skowronski J, Solter D, Knowles B. Gene expression during preimplantation mouse development. Genes Dev 1991;6:1190-1201. 28. Pal SK, Crowell R, Kiessling AA, Cooper GM. Expression of proto- oncogens in mouse eggs and preimplatation embryos. Mol Reprod Dev 1993;35:8-15. 29. Ruffner H, Reis LFL, NF D, Weissmann C. Induction of type I interferon genes and interferon-inducible genes in embryonal stem cells devoid of interferon regulatory factor 1. Proc Natl Acad Sci USA 1993;90:11503- 11507. 30. Belhumeur P, Lanoix J, Blais Y, Porget D, Steyaert A, Skup D. Action of spontaneously produced beta interferon in differentiation of embryonal carcinoma cells shrough an autoinduction mechanism. Mol Cell Biol 1993;13:2846-2857. 31. Haggarty A, Ponton A, Paterno GD, Diagneault L, Skup D. An embryonic DNA-binding protein specific for a region of the human IFN-beta1 promoter. Nucl Acids Res 1988;16: 10575-10592. 32. Van den Broek MF, Muller U, Huang S, Aguet M, Zinkernagel RM. Antiviral defense in mice lacking both alpha/beta and gamma interferon receptors. J Virology 1995;69: 4792-4796. 33. Hekman ACP, Trapman J, Mulder AH, van Gaalen JLM, Zwarthoff EC. Interferon expression in testes of transgenic mice leads to sterility. J Biol Chem 1988;263:12151-12155. 34. Riego E, Castro FO, de la Fuente J. Reduced transgenic efficiency in mice with transgenes directing a high level expression of human interferon regulatory factor-1. Interferon and Cytokine Res 1995;15 (Suppl. 1): S130. 35. Daugherty B, Martin-Zanca D, Helder B, Collier K, Seamans TC, Hotta K, Petska S. Isolation and bacterial expression of a murine alpha leukocyte interferon gene. J IFN Res 1984;4:635-643. Copyright 1997 Elfos Scientiae The following images related to this document are available:Line drawing images[ba97051d.gif] [ba97051b.gif] [ba97051a.gif] [ba97051c.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}