 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Obtencion y purificacion de neuraminidasa de Vibrio cholerae O1 Mairet Perez,^1 Hilda M Garcia,^2 Jose L Perez,^2 Barbara Cedre^2 y *Luis Garcia^2
^1 Division Vacunas. Centro de Ingenieria Genetica y Biotecnologia, AP
6162, Ciudad de La Habana, Cuba. E-mail:
Ricardo.Silva@cigb.edu.cu Recibido en diciembre de 1996. Aprobado en julio de 1997.
Code Number:BA97057
Size of Files:
Text: 28.7K
Graphics: Line drawings (gif) - 10.1K
Photographs (jpg) - 23.7K
ABSTRACT Neuraminidase, a secreted enzyme from Vibrio cholerae O1, is considered a virulence factor, because it increases the number of cholera toxin's receptors in the gut. Besides, this protein is widely used in investigations involving sialocompounds, in order to know the role of sialic acid residues in different biologic functions. In this work we compared neuraminidase expression levels in 5 V. cholerae O1 strains from different biotypes and serotypes. The 569B strain (Biotype Classic, Serotype Inaba) had the highest level and with this strain we designed a purification procedure that included: ammonium sulphate precipitation combined with pH of minimum solubility of the protein, gel-filtration chromatography (Sephacryl S-200HR), and finally ion-exchange chromatography (Q-Sepharose FF). This scheme gave us a 30 % of recovery with 98 % purity as determined by gel-filtration in HPLC. Purified neuraminidase was used for the pre-treatment of human sera previous to a hemagglutination- inhibition test with influenza type B virus. The enzyme was effective in the removal of non-specific inhibitors of the sera and it yielded similar results that a standard neuraminidase from a commercial source. Key words: neuraminidase, V. cholerae, purification RESUMEN La neuraminidasa, una enzima secretada por Vibrio cholerae O1, es considerada un factor de virulencia por aumentar el numero de receptores a la toxina de este microorganismo en los enterocitos. Ademas, esta enzima es de amplio uso en las investigaciones con sialocompuestos, para el esclarecimiento de la participacion de los restos de acido sialico en diferentes funciones biologicas. En el presente trabajo comparamos los niveles de expresion de esta enzima por 5 cepas de diferentes biotipos y serotipos de V. cholerae O1. Se selecciono la cepa 569B (Biotipo Clasico, Serotipo Inaba) como la mas productora, y a partir de ella se establecio un protocolo de purificacion que incluyo: precipitacion con sulfato de amonio al pH de menor solubilidad de la proteina, y cromatografias de filtracion en gel (Sefacril S-200HR) e intercambio ionico (Q-Sefarosa FF). Estos pasos suministraron la enzima con un rendimiento del 30 % y un nivel de pureza del 98 % determinado por gel-filtracion en HPLC. La neuraminidasa obtenida fue utilizada para el tratamiento de sueros humanos previo a la prueba de inhibicion de la hemaglutinacion con virus influenza tipo B. La enzima fue efectiva en la eliminacion de inhibidores inespecificos de los sueros y dio resultados similares a los de una neuraminidasa comercial utilizada como control. Palabras claves: neuraminidasa, V. cholerae, purificacion Introduccion La bacteria Vibrio cholerae O1 es el agente etiologico del colera, una infeccion intestinal aguda que puede llevar a una rapida deshidratacion, acidosis, y a la muerte en el 50 % de los casos graves. En la patogenesis de la enfermedad el evento de mayor importancia es la secrecion al lumen intestinal de una enterotoxina cuyo receptor celular es el gangliosido GM1. La subunidad A de esta proteina posee la actividad ADP- ribosil transferasa que desencadena las reacciones bioquimicas causantes de los sintomas caracteristicos de la enfermedad: activacion de adenilato- ciclasa, inhibicion del consumo de Na^+, aumento de la secrecion de Cl^-, y grandes perdidas de fluido en general (1). El V. cholerae O1 secreta tambien otras proteinas consideradas importantes factores de virulencia. Una de estas proteinas es la enzima neuraminidasa, una glicosidasa que actua a nivel intestinal y convierte los oligosialogangliosidos a GM1; de esta forma aumenta el numero de receptores a la toxina y la cantidad efectiva de toxina acoplada a los mismos (2, 3). Ademas de su funcion en la patogenia del colera, la enzima neuraminidasa es muy usada en la practica con fines investigativos. La reaccion que cataliza es la liberacion de los residuos terminales de acido N- u O- acilneuraminico presentes en oligosacaridos, polisacaridos, mucopolisacaridos, glicoproteinas y glicolipidos. Como dichos residuos de acido sialico son constituyentes de muchos receptores de toxinas, hormonas y neurotransmisores, asi como de moleculas de adhesion celular, los tratamientos con neuraminidasa resultan muy valiosos en los estudios basicos sobre glicoconjugados y en el esclarecimiento de la posible participacion de estos residuos en la funcion biologica particular (4, 5). En el presente trabajo estudiamos la produccion de neuraminidasa por 5 cepas pertenecientes a diferentes biotipos y serotipos de V. cholerae O1 con el objetivo de seleccionar una cepa buena productora, de la cual obtener la enzima con la mayor pureza y rendimiento posibles. La neuraminidasa de V. cholerae ha sido purificada con anterioridad usando la cromatografia de adsorcion en hidroxiapatita (6). Tambien ha sido producida en la bacteria Escherichia coli y purificada mediante precipitacion salina y filtracion en gel (7). En nuestro caso, para la obtencion de un preparado enzimatico de alta pureza combinamos una precipitacion salina diferencial con las cromatografias de filtracion en gel e intercambio ionico.
La funcionalidad de la enzima purificada se verifico al tratar con ella los sueros que serian evaluados en inhibicion de la hemaglutinacion viral. Materiales y Metodos Cepas bacterianas y condiciones de crecimiento Se utilizaron 5 cepas pertenecientes a la especie V. cholerae O1 de diferentes biotipos y serotipos:
- 395 (Biotipo Clasico, Serotipo Ogawa), Center for Vaccine Development, EE. UU., J. Kaper. - T19479 (Biotipo El Tor, Serotipo Inaba), Depto. Microbiologia e Inmunologia, Universidad de Goteborg, Suecia, A.M. Svennerholm. - JBK70 (Biotipo El Tor, Serotipo Inaba) y N16961 (Biotipo El Tor, Serotipo Inaba), Centro de Investigaciones de Enfermedades Infecciosas, Cuernavaca, Mexico, J. Sanchez.
El cultivo se centrifugo a 6 400 g durante 15 min a 4 C y el sobrenadante con la proteina de interes se recupero. Actividad enzimatica Las muestras se diluyeron 1:2 (v:v) en solucion tampon acetato 0,1 M con cloruro de calcio 0,02 M pH 5,5 (6). A la mezcla de reaccion se adiciono el sustrato (GD1a 9 mM en la misma solucion tampon) en una relacion 1:5 (v:v). Las mezclas se incubaron durante 30 min a 37 C. Las condiciones para la actividad enzimatica (concentracion de sustrato y tiempo de reaccion) habian sido previamente estandarizadas por nosotros (no se muestran los resultados). La reaccion se detuvo calentando 2 min a 100 C. La cantidad de acido N-acetil neuraminico (NANA) liberado fue determinado cualitativamente por cromatografia en capa fina empleando Silica gel 60 (MERCK) como soporte; se revelo con resorcinol-HCl (8), o se determino cuantitativamente por el metodo del acido tiobarbiturico (9). La actividad enzimatica fue expresada en unidades internacionales, donde:
El sobrenadante libre de celulas fue concentrado 10 veces usando un equipo Hollow-fiber DC-2 (AMICON) con membrana de poro de exclusion 10 kDa. A continuacion se realizo una precipitacion con sulfato de amonio al 40 % de saturacion durante 2 h a 4 C. La proteina precipitada se removio por centrifugacion a 6 400 g a 4 C durante 15 min, y al sobrenadante se le anadio sulfato de amonio hasta el 60 % de saturacion, y se dejo precipitar durante 30 min a 4 C; en este segundo paso de precipitacion se ajusto el pH a 3,5 con acido ortofosforico 17 % (p:v). El precipitado obtenido se resuspendio a una concentracion de proteinas final de 25 mg/mL en solucion tampon fosfato 1 mM pH 6,8 y se aplicaron 5 mL en una matriz Sefacril S-200HR (Pharmacia, LKB) equilibrada en igual solucion tampon y empacada en una columna XK 16/100 (Pharmacia, LKB). Se colectaron fracciones de 3 mL, a las cuales se les determino cuantitativamente la actividad enzimatica; las fracciones con valores por encima de 0,2 U/mL se reunieron y esta muestra se aplico en una cromatografia de intercambio ionico empleando la matriz Q-Sefarosa FF (Pharmacia, LKB) equilibrada en solucion tampon fosfato 1 mM pH 6,8. Luego de lavar con 10 volumenes de columna, las proteinas se eluyeron y asi aumento la fuerza ionica hasta 1M de cloruro de sodio. Estas muestras fueron dializadas contra tampon acetato pH 5,5 durante 12 h a 4 C, y en esas condiciones conservadas a 4 C.
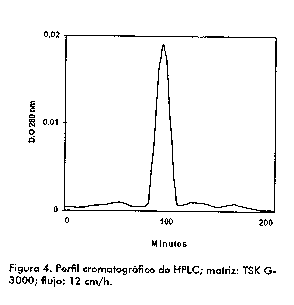
La pureza e integridad de las muestras se verifico por cromatografia liquida de alta resolucion (HPLC) de filtracion en gel, con una columna TSK G-3000 (Toyo SODA, Japon).
La determinacion de proteinas se realizo por el metodo de Lowry (10). Inhibicion de la hemaglutinacion Se realizo el ensayo de inhibicion de la aglutinacion de eritrocitos con el virus de influenza humano tipo B; para ello se emplearon sueros de individuos antes y un mes despues de haber recibido una vacuna antigripal (vacuna viva atenuada adaptada al frio B/14/5/1, del Instituto de Medicina Experimental de San Petersburgo, Rusia). Para la eliminacion de los inhibidores inespecificos de la hemaglutinacion existentes en los sueros se empleo el siguiente tratamiento enzimatico (11):
Los sueros se diluyeron 1:6 (v:v) en tampon acetato incluyendo 2,5 mUI de neuraminidasa de V. cholerae por cada muestra, y se incubaron 18 h a 37 C. En nuestro caso usamos una enzima de referencia (neuraminidase from V. cholerae, Boehringer Mannheim 20 UI/mg) asi como la enzima purificada en este trabajo.
A continuacion se anadieron 8 unidades hemaglutinantes del virus de influenza humano tipo B y se realizo el ensayo de aglutinacion con eritrocitos humanos (11).
Los titulos de los sueros fueron considerados como la mayor dilucion del suero donde se observo el boton correspondiente a los hematies no aglutinados, y los titulos superiores a 1:80 fueron considerados positivos. Resultados y Discusion Obtencion de neuraminidasa
Tabla 1. Relacion entre la actividad enzimatica y el rendimiento celular.
---------------------------------------------------------------------------
Cepas N16961 JBK70 T19479 569B 395
---------------------------------------------------------------------------
Act. enz. cuantitativa (U/mL) 0,06 0,077 0,055 0,112 0,094
Act. enz. cualitativa +(*) ++ + +++++ +++
Rend. celular (g/L) Peso humedo 8,32 10,32 9,94 7,28 8,1
Act. enz. / rend. cel. (U/mg) 0,0072 0,0075 0,0055 0,015 0,011
---------------------------------------------------------------------------
(*): Un mayor numero de signos positivos se corresponde con una mayor
actividad enzimatica cualitativa.
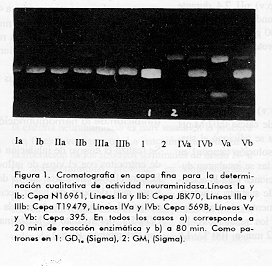
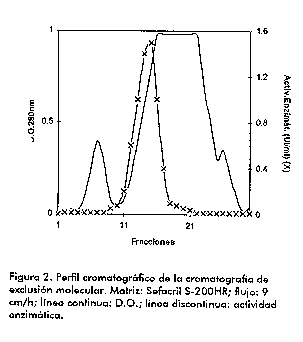
Para cada cepa se evaluo la actividad enzimatica cualitativamente para dos tiempos de incubacion muestra-sustrato (20 y 80 min de reaccion) con el objetivo de ver si se lograba un total agotamiento del GD1a. Como se muestra en la Figura 1, esto se logro con la cepa 569B coincidiendo con el resultado de la determinacion cuantitativa anterior. Con la utilizacion de la tecnica de cromatografia en capa fina se determino la especificidad de sustrato que presenta la enzima estudiada, y se comprobo que solo libera el resto de acido sialico mas externo del GD1a (que es el que cuantificamos por el metodo del acido tiobarbiturico). A pesar de tener el GM1 otro resto de acido sialico que podria ser liberado, este no es utilizado como sustrato por la enzima pues esta inaccesible por impedimentos estericos en la estructura del gangliosido (13, 14). Como se muestra en la Figura 1, incluso a los 80 min de accion de la neuraminidasa de la cepa 569B, en que el GD1a desaparecio completamente, el GM1 se mantuvo sin ser desialilado por la enzima. Purificacion de la enzima Durante el proceso de purificacion de neuraminidasa determinamos la actividad cuantitativamente por el metodo del acido tiobarbiturico (9). En el proceso seguido para la purificacion partimos de un sobrenadante de cultivo concentrado diez veces para facilitar el trabajo con volumenes menores. El poro de exclusion de la fibra utilizada fue de 10 kDa, lo que permite la eliminacion de contaminantes con pesos moleculares menores de esa talla, asi como productos de degradacion. Seguidamente, usando un efecto combinado de la fuerza ionica y el pH de minima solubilidad de la enzima (determinado por nosotros en experimentos previos no mostrados), logramos enriquecer la proteina de interes a partir del crudo inicial concentrado. En el precipitado obtenido con un 40 % de saturacion de sulfato de amonio hubo una minima perdida de actividad enzimatica, la que quedo mayoritariamente en el sobrenadante. En el otro paso de la precipitacion fraccionada no detectamos ninguna actividad neuraminidasa en el sobrenadante y si en el precipitado, si bien la recuperacion no fue total. Esto pudiera deberse a una menor estabilidad de la enzima a pH 3,5, lo que provocaria desnaturalizacion de una parte de la misma y perdida de su actividad. No obstante, preferimos este metodo pues resulto mas efectivo en la precipitacion total de la proteina que cuando no ajustamos el pH y se precipitaba solo por el efecto de la sal. Al utilizar solo una precipitacion fraccionada con sulfato de amonio como en trabajos anteriores (15) es necesario en el segundo paso adicionar sulfato de amonio hasta un 80 % de saturacion para que precipite completamente la proteina de interes; con ello aumentan tambien los contaminantes precipitados a esa mayor fuerza ionica. En nuestro caso, al complementar el efecto de la sal con el del pH de minima solubilidad, se favorecio la precipitacion preferencial de la neuraminidasa con un por ciento de saturacion de sulfato de amonio inferior (60 %) al anteriormente utilizado. El precipitado del paso anterior se resuspendio en un volumen minimo de tampon fosfato 1 mM pH 6,8, aproximadamente 15 mL (concentracion de proteinas 25 mg/mL), asi se restituyo a la enzima en un valor de pH en el cual resulta mas estable, y en estas condiciones se realizaron los pasos cromatograficos siguientes. Las muestras reconstituidas tras la precipitacion quedan con una alta concentracion salina, la que es eliminada mediante el paso de filtracion en gel que se realiza a continuacion. Aunque la cromatografia de exclusion molecular tiene limitacion en el volumen de aplicacion, pudimos realizar un procesamiento rapido de toda la muestra al resuspender el precipitado en un volumen pequeno. Probamos las matrices Sefacril S-100HR y Sefacril S-200HR, y se obtuvieron cromatogramas similares al mostrado en la Figura 2 en ambos casos. En el Sefacril S-200HR, la actividad enzimatica se detecto solo en la subida e inicio del segundo pico; de este modo se aislo la enzima de los contaminantes que eluian primero (Figura 2). En el caso del Sefacril S- 100HR, la actividad se detectaba tanto en el primero como en el segundo pico, por lo que decidimos trabajar con la matriz Sefacril S-200HR en nuestro esquema final de purificacion. La neuraminidasa de V. cholerae obtenida recombinante en E. coli, ha sido purificada por otros autores utilizando una matriz Sefacril-300 (7) que en nuestro caso no consideramos necesario utilizar, pues como ya ha sido mencionado logramos una buena resolucion con la matriz Sefacril S-200HR. Por demas, el peso molecular de la proteina a purificar (aproximadamente 85 kDa), se encuentra en el rango de separacion de las dos matrices probadas en este trabajo.
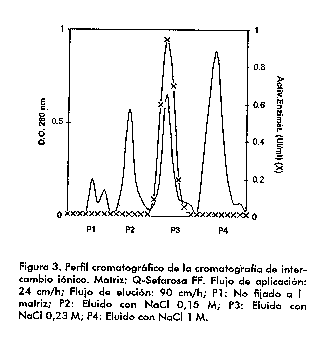
Posteriormente, la muestra (fracciones de la 11 a la 17, Figura 2) fue aplicada en un intercambiador anionico al cual debe fijarse la neuraminidasa al encontrarse por encima de su pH isoelectrico (pI < 5) en las condiciones de la corrida (tampon fosfato 1 mM pH 6,8). En este paso se pudo procesar un volumen grande de muestra por la alta capacidad de la matriz empleada (Q-Sefarosa FF). Para la elucion, se fue aumentando gradualmente la concentracion de cloruro de sodio, y a las fracciones colectadas se les determino actividad enzimatica. Finalmente, decidimos utilizar 0,15 M NaCl para la eliminacion de los contaminantes que se pegaban mas debilmente a la matriz; 0,23 M NaCl para eluir la neuraminidasa en una sola fraccion, y 1 M NaCl para el lavado final de la columna. El cromatograma tipico obtenido con estas condiciones de elucion aparece en la Figura 3.
Tabla 2. Resumen del proceso de purificacion.
---------------------------------------------------------------------------
Pasos de la Unidades Proteina Proteina Actividad Factor de Rendimiento
purificacion totales (mg/mL) total especifica purifi- (%)
(U) (mg) (U/mg) cacion
---------------------------------------------------------------------------
Sobrenadante 258 14 35 000 0,0074 1 100
Concentrado 240 27 6 750 0,035 4,7 93
Sulf. de amonio 170 25 725 0,23 31 66
Sefacril S-200HR 150 0,4 150 1 135 58
Q-Sefarosa FF 78 0,2 24 3,25 439 30
---------------------------------------------------------------------------
En la Figura 4 se muestra la evidencia (filtracion en gel HPLC) del alto grado de pureza (98 %) alcanzado tras los dos procesos cromatograficos. La recuperacion global de la actividad de la enzima fue del 30 % tomando como referencia la actividad inicial existente en el sobrenadante de cultivo. Resultados similares en cuanto a rendimiento han sido obtenidos en trabajos de purificacion anterior tanto para la neuraminidasa natural purificada por cromatografia de adsorcion en hidroxiapatita (6) como para la recombinante producida en E. coli y purificada por Sefacril-300 (7).
La actividad enzimatica de nuestra preparacion purificada se mantuvo invariable durante al menos 6 meses de conservacion a 4 C (no se muestran los resultados), lo que sugiere que la misma esta libre de proteasas contaminantes secretadas al medio por V. cholerae (16), las que en cantidades trazas pudieran degradar en largos periodos de tiempo a la enzima obtenida.
Por ultimo, nuestro protocolo ha resultado ser de excelente reproducibilidad en nuestras condiciones experimentales, y en principio resulta escalable para la produccion de cantidades mayores de la enzima. Inhibicion de la hemaglutinacion En los sueros humanos normales existen inhibidores no especificos de la reaccion de aglutinacion del virus de la influenza con los eritrocitos. Estos inhibidores que se suponen sean sialocompuestos pueden ser eliminados mediante tratamientos quimicos o enzimaticos, se obtienen menos frecuencia de falsos seropositivos y por tanto, valores mas reales en estos ensayos (11). Con el metodo enzimatico, en el que se aplica el tratamiento con la enzima neuraminidasa, se obtiene una mayor especificidad que con la oxidacion quimica con periodato, por lo que generalmente es preferible para este proposito. En nuestro trabajo los sueros fueron tratados enzimaticamente como se menciona en la seccion de Materiales y Metodos. Los resultados obtenidos son mostrados en la Tabla 3. En ambos casos se aprecia que despues de la vacunacion aumenta el numero de seropositivos. La comparacion de los resultados obtenidos empleando la neuraminidasa purificada en este trabajo y la disponible comercialmente (Boehringer-Mannheim) reflejo un comportamiento muy similar de las dos enzimas. Tabla 3. Resultados de la prueba de inhibicion de la hemaglutinacion.
-----------------------------
Vacunacion Tratamientos
-----------------
A B
-----------------------------
Antes SP 3 3
SN 25 25
Despues SP 9 10
SN 19 18
-----------------------------
A: tratamiento enzimatico de los sueros (neuraminidasa comercial); B:
tratamiento enzimatico de los sueros (neuraminidasa purificada en este
trabajo); SP: seropositivos; SN: seronegativos.Conclusiones La neuraminidasa de V. cholerae O1 puede ser obtenida hasta con un 98 % de pureza mediante el procedimiento descrito en este trabajo, el cual es escalable y repetible. La neuraminidasa obtenida resulta completamente funcional para ser utilizada en trabajos con sialocompuestos, por ejemplo en la eliminacion de inhibidores inespecificos del ensayo de la hemaglutinacion viral. Agradecimientos
Agradecemos al doctor Rodrigo Felipe Varela del Laboratorio de Medios Diagnosticos de la Division de Virologia del Instituto Finlay, por su asesoramiento para la realizacion de la prueba de inhibicion de la hemaglutinacion. Tambien al licenciado Enrique Munoz de la Unidad Analitica del propio Instituto, por su ayuda en la cromatografia de alta resolucion, y a Orlando L Pardo por la revision del manuscrito. References 1. Spangler BD. Structure and function of the cholera toxin and other related enterotoxins. Microbiol Rev 1992;56:622-636. 2. Kaper JB, Morris JG, Levine MM. Cholera. Clin Microbiol Rev 1994;1(1). 3. Galen JE, Ketley JM, Fassano A, Richardson SH, Wasserman SS, Kaper JB. Role of Vibrio cholerae neuraminidase in the function of cholera toxin. Infection and Immunity 1992;60:406-415. 4. Hulinska D, Volf P, Grubhoffer L. Characterization of Borrelia burgdorferi glycoconjugates and surface carbohydrates. Int J Med Microbiol Virol Parasitol Infect Dis 1992;276:473-480. 5. Rens-Domano S, Reisine T. Structural and functional role of the carbohydrate component of somatostatin receptors. J Biol Chem 1991;266:20094-20102. 6. Ada GL, French EL, Lind PE. Purification and properties of neuraminidase from Vibrio cholerae. J Gen Microbiol 1961;24:409-421. 7. Taylor G, Vimr E, Garman E, Laver G. Purification, crystallization and preliminary cristallographic study of neuraminidase from Vibrio cholerae and Salmonella typhimurium LT2. J Mol Biol 1992;226:1287-1290. 8. Svennerholm L. Procedure for quantitative isolation of brain gangliosides. Neurochem 1957;24:604-611. 9. Aminoff D. Methods for the quantitative estimation of N-acetyl neuraminic acid and their application to hydrolysates of sialomucoids. Biochem J 1961;81: 384-391. 10. Lowry OH. Protein measurement with the Folin phenol reagent. J Biol Chem 1951;193:265-275. 11. Subi K. The hemagglutinating unit of Influenza virus in the hemagglutination-inhibition test. Rev Roum Med Virol 1984; 35:69-70. 12. Kabir S, Ahmad N, Ali S. Neuraminidase production by Vibrio cholerae O1 and other diarrheagenic bacteria. Infection and Immunity 1984;44:747- 749. 13. Portoukalian J, Bouchan B. Hydrolysis of all gangliosides by Vibrio cholerae neuraminidase. J of Chromatography 1986;380:386-392. 14. Ogura K, Ogura M, Anderson RL, Sweley CC. Peroxidase-amplified assay of sialidase activity toward gangliosides. Anal Biochemistry 1992;200:52- 57. 15. Vimr E, Galen J, Lawrisuk L, Kaper JB. Cloning and expression of the Vibrio cholerae neuraminidase gene nanH in Escherichia coli. J Bacteriology 1988; 170:1495-1504. 16. Schneider DR, Parker CD. Isolation and characterization protease deficient mutants of Vibrio cholerae. J Infect Dis 1978;138:143-149. Copyright 1997 Elfos Scientiae The following images related to this document are available:Photo images[ba97057a.jpg]Line drawing images[ba97057b.gif] [ba97057d.gif] [ba97057c.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}