 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Clonacion de mamiferos adultos. El camino iniciado Fidel O Castro Division de Genetica de Celulas de Mamiferos. Centro de Ingenieria Genetica y Biotecnologia. Apartado postal 6162. La Habana, Cuba.
Code Number:BA97063
Size of Files:
Text: 31.9K
Graphics: Photographs (jpg) - 77.3K
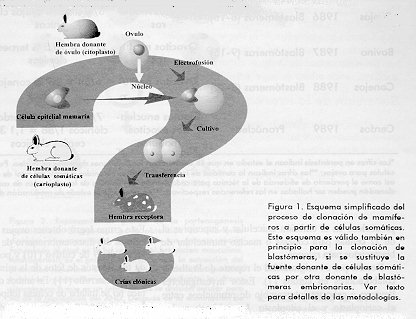
El ancestral mito de la irreversibilidad de la diferenciacion celular en animales ha sido al parecer roto con la aparicion del primer mamifero obtenido por clonacion a partir de tejido somatico de un individuo adulto (1). Dolly, la oveja asi nombrada por los cientificos escoceses del Instituto Roslin, que la generaron en julio del pasado ano, ha devenido motivo de polemica de cientificos, politicos, religiosos y de la sociedad en general. Clonacion La clonacion es la confeccion de copias identicas de algun elemento biologico. Asi tenemos clonacion de genes en plasmidos u otros vectores, que permiten la multiplicacion de copias exactas de dicho gen dentro del vector. En el campo de la inmunologia, la perpetuacion de una especificidad antigenica reflejada en anticuerpos unicos contra esta y descendientes de una estirpe unica, es tambien clonacion. Desde el punto de vista de organismos enteros, la clonacion se refiere a la obtencion de seres geneticamente identicos. En plantas, por ejemplo, la clonacion somatica permite propagar en miles de copias a una especie determinada. En mamiferos, los gemelos monocigoticos son clones que surgen de forma espontanea al producirse una duplicacion aberrada (generalmente por biparticion) de un embrion preimplantatorio. Experimentalmente se han generado mamiferos clonicos mediante el aislamiento de blastomeras totipotentes a partir de embriones de entre 8 y 16 celulas, una blastomera es fundida con un ovocito maduro, previamente enucleado. De este modo se logra, aunque con una eficiencia muy baja, que el nucleo de la blastomera gobierne los eventos subsiguientes de desarrollo embrionario. Este control nuclear es a su vez condicionado, al menos en las primeras etapas, por factores citoplasmaticos aun no identificados.
La clonacion somatica (Figura 1) cuyo unico exponente hasta el presente es Dolly, es en principio similar a aquella de blastomeras, pero fundiendo con el ovocito maduro y enucleado una celula de origen no embrionario (somatica) que debe en principio adquirir la capacidad de retornar a un estado no diferenciado para poder asumir el control del desarrollo embrionario subsiguiente.
La clonacion en animales data de la decada del 50 cuando Briggs y King lograron a partir de celulas de blastula de Rana pipiens fundidas con ovocitos de la misma especie, obtener animales identicos (2). Sin embargo la clonacion de mamiferos sobrevino casi 30 anos mas tarde y fue lograda por Illmensee y Hope (3) en Bar Harbor Maine, en Estados Unidos, como material genetico de partida estos cientificos emplearon celulas del macizo intracelular obtenidas de blastocistos de raton las cuales fueron fundidas mediante virus Sendai inactivado, con cigotos unicelulares recien fecundados y enucleados. En la segunda mitad de los 80, se lograron clonaciones exitosas en la mayoria de las especies de importancia economica (2-7; Tabla 1) Partiendo en casi todos los casos de blastomeras totipotentes aisladas de embriones de entre 8 y 32 celulas y fundiendolas con pulsos electricos a ovocitos maduros y enucleados. Tabla 1. Antecedentes de la clonacion en animales a partir de blastomeras.
---------------------------------------------------------------------------
Especie Ano Origen celular* Recipiente Resultado** Referencia
---------------------------------------------------------------------------
Rana 1952 Blastula Ovocitos*** Ranas clonicas 2
Raton 1981 ICM Cigotos 3/16 = 18,8 %
enucleados ratones clonicos 3
Ovejas 1986 Blastomeras Ovocitos maduros 3/4 = 75 % 4
(8-16) ovejas clonicas
Bovino 1987 Blastomeras Ovocitos maduros 2/19 = 10,5 % 5
(9-15) terneros clonicos
Conejos 1988 Blastomeras Ovocitos maduros 6/64 = 3,7 % 6
(8-32) conejos clonicos
Cerdos 1989 Pronucleos Cigotos enucleados 7/56 = 12,5 %
ovocitos maduros cerdos clonicos
1/88 = 1,1 3 %
cerdos clonicos 7
---------------------------------------------------------------------------
* Las cifras en parentesis indican el estadio en que se encontraban los
embriones donantes de nucleo. Por ejemplo, entre 8 y 16 celulas para
ovejas; ** las cifras indican la cantidad de individuos clonicos obtenidos a partir del total de embriones transferidos, asi como el porciento de eficiencia de la tecnica para cada especie. Los detalles de las eficiencias de cada paso del proceso de clonacion pueden ser hallados en las referencias respectivas; *** no se estudio el estado de maduracion del ovocito.
Los primeros avances de importancia fueron publicados por Willadsen en 1986 trabajando con embriones y ovocitos de ovejas. La evaluacion de la capacidad de reprogramacion nuclear de blastomeras en diferentes estados de desarrollo fue el objetivo de estudio principal en los trabajos pioneros de finales de la decada de los 80. De este modo se pudo demostrar que si bien puede variar el potencial de desarrollo de las blastomeras aisladas en dependencia de la especie, los estadios entre 8 y 16 celulas son los optimos para realizar la clonacion a partir de blastomeras, siendo los limites inferiores el estado de pronucleos en el embrion unicelular y superiores el de 32 celulas para conejas o macizo intracelular de blastocistos para ratones. Resalta por su originalidad el reporte de Prather y colaboradores en cerdos (7). Estos investigadores llevaron a cabo un intercambio de pronucleos entre el carioplasto y el citoplasto. Los cigotos recien fecundados fueron enucleados y en su lugar recibieron pronucleos de los embriones donantes de nucleo. La eficiencia del proceso fue relativamente alta (12,5%) sin embargo cuando emplearon ovocitos maduros enucleados para recibir nucleos pronucleares la eficiencia disminuyo considerablemente. Esto puede haber estado relacionado con la presencia de factores citoplasmaticos activos en el cigoto inmediatamente despues de la fertilizacion y que son requeridos por los pronucleos para su ulterior division. El exito alcanzado con la clonacion convencional o de blastomeras no conllevo a avances fundamentales en la clonacion a partir de celulas somaticas. Aunque los estudios realizados en ese sentido no aparecieron publicados debido a sus resultados negativos, no hay duda de que si se estuvieron llevando a cabo en ciertos laboratorios del mundo. Una vez mas los antecedentes de este nuevo sistema de clonacion surgieron en anfibios, gracias a los trabajos de Gurdon y colaboradores (8), quienes obtuvieron renacuajos (pero no ranas adultas) a partir de queratinocitos de Xenopus laevi trasplantados a ovulos cuyo material genetico habia sido eliminado. Los primeros avances de importancia hacia la clonacion somatica en mamiferos fueron precisamente del grupo escoces del Instituto Roslin en Edimburgo (1, 10, 11; Tabla 2). Este grupo logro obtener ovejas clonicas a partir de celulas del disco embrionario de embriones de oveja en el dia 9 de cultivo (10) y posteriormente a partir de fibroblastos de fetos de la misma especie en el dia 26 del desarrollo (11). En ambos casos, es cierto que si bien no se trataba de celulas totipotentes, si eran celulas pluripotentes. Los discos embrionarios del dia 9 de cultivo son capaces de dar lugar a multiples estructuras celulares y de tejido en el embrion en formacion. Los fibroblastos fetales daran lugar a distintas estirpes de celulas tales como osteoblastos y mioblastos. Tabla 2. Antecedentes de la clonacion en animales a partir de otras estirpes celulares.
---------------------------------------------------------------------------
Especie Ano Origen celular Recipiente Resultado* Referencia
---------------------------------------------------------------------------
Xenopus 1975 Keratinocitos Ovocitos** Renacuajos 8
Raton 1993 Celulas totipotentes Embriones 2/34 = 6 %
embrionarias tetraploides*** ratones clonicos 9
Ovejas 1994 Disco embrionario Ovocitos maduros 3/20 = 15 %
dia 9 de cultivo ovejas clonicas 10
Ovejas 1996 Fibroblastos Ovocitos maduros 8/32 = 25 %
embrionarios dia 26 ovejas clonicas 11
Ovejas 1997 Glandula mamaria Ovocitos maduros 1/29 = 3,4 %
ovejas clonicas 1
---------------------------------------------------------------------------
* Las cifras indican la cantidad de individuos clonicos obtenidos a partir
del total de embriones transferidos, asi como el porciento de eficiencia de
la tecnica para cada especie. Los detalles de las eficiencias de cada paso
del proceso de clonacion pueden ser hallados en las referencias
respectivas; ** se desconocia el estado de maduracion del citoplasma del ovocito; *** las celulas embrionarias totipotentes fueron fundidas con embriones de 4 celulas tetraploidizados mediante fusion electrica. Ver texto para detalles.
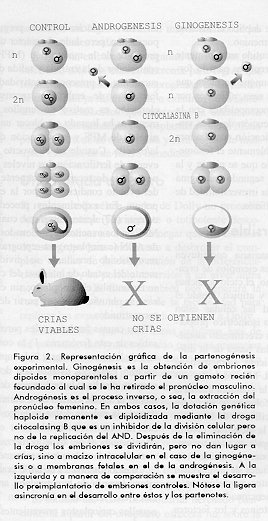
De especial interes fue el reporte de Nagy y colaboradores en 1993 (9) en el cual se obtuvieron ratones a partir de celulas embrionarias totipotentes (CET). Estas celulas que son aisladas a partir de blastocistos, son como la palabra lo dice totipotentes, cuando son agregadas con morulas o microinyectadas a blastocistos. En otras palabras, ellas necesitan la cooperacion de celulas embrionarias para colonizarlas y contribuir a la formacion de ciertas estirpes celulares. Es este el principio empleado para obtener animales transgenicos con delecion de genes o con recombinacion homologa. Sin embargo en el articulo en discusion, los autores fundieron CET con embriones cuatricelulares tetraploides. El objetivo de esta manipulacion es el de preparar un recipiente "inerte" (los embriones tetraploides no son viables y por tanto no contribuyen a la formacion de estirpes celulares) sobre el cual se fundieron las CET y estas por si solas dieron lugar a ratones viables. No cabian dudas ya respecto a que no eran estrictamente necesarias poblaciones de celulas totipotentes (blastomericas) para lograr la clonacion, comenzaba a abrirse la esperanza pues, de clonar a un individuo adulto. El resto de la historia es ya conocida, Dolly se convirtio en el primer mamifero nacido a partir de una celula adulta. Dolly, un fenomeno trascendental Desde el punto de vista experimental es indudable que el fenomeno del nacimiento de Dolly significa un avance trascendental en la biologia moderna. En esencia se han roto dos paradigmas hasta ahora inamovibles de la biologia de los mamiferos, a saber; la necesidad de los dos parentales para la obtencion de una nueva progenie y la irreversibilidad de la diferenciacion celular. Ginogenesis? Con respecto al primer punto, estudios que datan de la decada de los 70 demostraron firmemente que los embriones requerian de la participacion de ambos gametos en su concepcion durante la fertilizacion para poder generar un feto y consecuentemente un nuevo individuo. Los experimentos clasicos de produccion de embriones monoparentales a traves de la androgenesis y ginogenesis en ratones (Figura 2) demostraron que los embriones androgeneticos cuando eran trasplantados a madres receptoras producian solo tejidos derivados del trofoblasto embrionario (membranas fetales), mientras que aquellos embriones producidos por ginogenesis daban lugar solo a tejido embrioblastico (12-15). Ambos fenomenos eran incompatibles con la formacion de un nuevo individuo. Era obvio que se necesitaba la conjugacion de las dos dotaciones geneticas para completar la embriogenesis. Al parecer, la metilacion diferencial a la que es sometida el genoma paterno influye decisivamente sobre el futuro de la actividad transcripcional y mitotica del embrion recien formado (15). Por otra parte la ausencia del cromosoma X en los androgenones producidos a partir de espermatozoides Y es toxica y letal para la celula (14, 15).
Este suceso echa por tierra lo discutido anteriormente sobre la necesidad de ambos parentales para la formacion de un nuevo individuo, aunque no niega que la ginogenesis y la androgenesis sean, al menos por el momento, eventos celulares incompatibles con la reproduccion a termino y la obtencion de crias viables. No es facil hallar una explicacion conciliadora entre lo clasico y Dolly y, probablemente el fenomeno sea mucho mas complejo de lo que se piensa y la respuesta hay que buscarla en el segundo paradigma roto por el nacimiento de Dolly, la irreversibilidad de la diferenciacion celular en mamiferos. Reversion de lo irreversible Las celulas de la glandula mamaria constituyen probablemente uno de los mejores ejemplos de organizacion secretora en el organismo, el ciclo de dichas celulas es en extremo complejo y atipico. Desde el nacimiento y hasta la pubertad las celulas estan sujetas primero a un crecimiento isometrico proporcional al corporal y despues a un crecimiento alometrico durante el cual se sientan las bases de la diferenciacion secretora. El crecimiento alometrico se lleva a cabo durante la pubertad. Mas adelante, durante la gestacion, las celulas de la glandula mamaria terminan su diferenciacion morfologica y secretora bajo el control de los genes especificos y de la matriz extracelular. Una vez concluida la lactacion, involucionan mediante un proceso apoptotico para comenzar de nuevo el ciclo, con la proxima gestacion (16). A medida que las celulas epiteliales de la glandula mamaria transitan por los eventos morfologicos y diferenciativos anteriormente mencionados, cambia tambien la naturaleza de las proteinas y los factores liberados por ellas. La celula madre de Dolly provino de una hembra en el ultimo tercio de la gestacion, en donde la actividad proliferativa debe de haber estado finalizando y las celulas se encontrarian en un estado diferenciado, secretor y cuasi quiescente desde el punto de vista mitotico. La pregunta es: ¨de que estirpe celular exactamente? Es sabido que en un cultivo primario obtenido a partir de la glandula mamaria se encuentran presentes poblaciones de celulas epiteliales, mioepiteliales, fibroblastos entre otros. Si bien es irrelevante a los efectos de la clonacion cual de las poblaciones existentes en un cultivo primario dio lugar a Dolly, ya que en cualquier caso implica la reprogramacion de la diferenciacion, no lo es la posibilidad de que en este coctel primario de celulas mamarias existan poblaciones de tipo pluripotencial que sean desconocidas aun para la comunidad cientifica y que a su vez esten involucradas en la regeneracion del tejido mamario despues del primer destete y durante las subsiguientes gestaciones. Que factores estan involucrados en la reprogramacion nuclear de una celula somatica fusionada a un ovocito enucleado? Esta pregunta no tiene respuesta aun, pero dado el caracter universal del factor promotor de la maduracion (MPF) como elemento controlador de la entrada y salida del ciclo celular en eucariotas, su presencia y actividad en los eventos de formacion de embriones reconstituidos mediante clonacion es motivo de amplios estudios. Actualmente se ha logrado establecer un vinculo entre los niveles de MPF y la activacion del complejo celula/ovocito. Cuando en el ovocito maduro ocurre el evento de fertilizacion, los niveles de MPF son altos, pero estos decrecen drasticamente con la activacion del ovocito condicionada por la entrada del nemaspermo. En experimentos precedentes, Wilmut y colegas (17) demostraron que cuando los niveles de MPF en los ovocitos eran elevados, la celula donante de ADN (carioplasto) era reprogramada y el embrion reconstituido resultante se dividia independientemente del estado de interfase del ciclo celular en que se encontraba el carioplasto; sin embargo en ningun caso se obtuvieron crias a partir de los embriones asi reconstituidos. Dos causas fundamentales parecen ser las responsables de este fenomeno: 1) la condensacion prematura de los cromosomas y 2) la ruptura de la envoltura nuclear. Estos dos eventos son detrimentales para la correcta ploidia y la replicacion del ADN respectivamente, si son llevados a cabo durante la interfase y no durante la mitosis. Estos hallazgos indican que algunos factores transactivadores u otros componentes citoplasmaticos del ovocito fuerzan al nucleo interfasico a llevar a cabo eventos meramente mitoticos. Cuando los niveles de MPF eran bajos en los ovocitos, despues de la activacion artificial con pulsos electricos, entonces estos eran receptivos solo a aquellos carioplastos provenientes de etapas G1 o S del ciclo celular, los embriones reconstituidos se dividian (y como se demostro, nacio Dolly a partir de uno de ellos). Cuando, en lugar de ovocitos se emplearon zigotos pronucleares enucleados, no se obtenian resultados adecuados. La conclusion parece clara, al menos MPF esta involucrado en la reprogramacion nuclear y es reclutado por los pronucleos del ovocito recien fertilizado. La importancia de este hallazgo fue trascendental para el establecimiento de las condiciones necesarias para el nacimiento de Dolly. Quiza otros grupos hayan subestimado el papel del estado de sincronia entre el citoplasto (ovocito receptor) y el carioplasto o celula donante del ADN. Es obvio que las celulas donantes de ADN deben de encontrarse en estadios diploides, para: 1) estar en sincronia con el ovocito metafasico y por ende diploide y 2) para mantener la ploidia correcta en el nuevo embrion resultante de la fusion. Por tanto las fases del ciclo celular S, G2 y M no son adecuadas para los experimentos de clonacion somatica, mientras que G1 es la etapa optima. Bajo ciertas condiciones la celula sale del ciclo celular durante la fase G1 y se queda detenida en un estado quiescente, de no crecimiento y no division, denominado G0. Muchas celulas en el organismo adulto, tales como las celulas nerviosas, del lente del ojo y de musculo estriado no se dividen nunca, otras como los fibroblastos que participan en la regeneracion de heridas crecen y se dividen al ser solicitado esto por el organismo, de otro modo permanecen en estado quiescente justo antes de la sintesis del ADN. Es probable que las celulas en este estado presenten una cierta relajacion de la cromatina que la hace mas asequible a factores de citoplasmaticos de reprogramacion, asi como a factores de transcripcion (18). En cultivo, las celulas son llevadas a G0 mediante la deprivacion de factores nutricionales especialmente el suero, esto conlleva a la suspension de la transcripcion de los genes codificantes para las ciclinas dependientes de quinasas, asi como para las ciclinas G1 (grupos D y E) y a la caida brusca de los niveles de las proteinas correspondientes. Como consecuencia, la celula queda desprovista de los factores auxiliadores para traspasar los puntos de restriccion del ciclo celular y quedan detenidas en G0.
Existen varias formas de saber si una celula se encuentra en la interfase o en mitosis dentro del ciclo celular, siendo la separacion activada por fluorescencia (FACS) la mas usada, ya que permite cuantificar la cantidad de ADN presente en una celula, a saber: x; para la fase G1, x-2x para la fase S, 2x para G2 y M. Sin embargo resulta dificil conocer exactamente si una celula detenida en G1 se halla fuera del ciclo o en G0. Entre los marcadores mas utilizados para ello, se encuentra el antigeno celular nuclear de proliferacion (PCNA). Este antigeno es un cofactor para la polimerasa delta y es absolutamente necesario para la replicacion del ADN; por lo tanto las celulas que se encuentren progresando a traves del ciclo celular necesitan de la sintesis de PCNA, mientras que aquellas detenidas en G0 no lo tendran.
En los experimentos del grupo del Instituto Roslin en Edimburgo se emplearon como carioplasto celulas quiescentes o en G0. Dicho estado fue determinado por ensayo inmunoenzimatico empleando anticuerpos anti PCNA/ciclina revelados con un segundo anticuerpo rodaminado. La validez del ensayo reside en los controles de celulas en division, ya que PCNA es muy dificil de detectar mediante anticuerpos; de ahi que es imprescindible dejar claro que su ausencia no es un artefacto experimental. Como citoplasto fueron utilizados ovocitos previamente enucleados y activados por pulsos electricos en los cuales los niveles de MPF eran bajos. El grupo escoces ha propuesto para estos citoplastos el nombre de "recipiente universal" para la clonacion. Es muy probable que estos meticulosos estudios de sincronizacion de los ciclos entre el cario y citoplastos hayan sido los responsables de la exitosa clonacion de Dolly y constituye sin lugar a dudas el mayor merito de las investigaciones realizadas. Surgen sin embargo interrogantes tales como: ¨fue suficiente solo esa exacta sincronia para lograr que los factores citoplasmaticos reprogramaran un nucleo de una celula ya diferenciada, o existiran otros factores aun desconocidos que gobiernan la interaccion entre el nucleo donante y el ooplasma receptor? Esto por supuesto no se conoce aun y sera objeto de multiples estudios en un futuro inmediato. Una mirada al futuro Si realmente la celula madre de Dolly no procedio de alguna estirpe pluripotente o totipotente desconocida, entonces no se puede negar que los estudios de este grupo han comenzado a desbrozar el complejo camino de la comprension de la diferenciacion celular y su posible reversion a un nivel antes no imaginado. El futuro tambien nos deparara la confirmacion o la negacion de la factibilidad para replicar los experimentos que dieron lugar a Dolly. Es indudable que otras lineas celulares han de ser probadas, incluyendo no solo cultivos primarios sino lineas bien establecidas y caracterizadas como carentes de actividad pluri o totipotencial y por que no, aquellas con sabido potencial pluripotente. El uso de lineas celulares conservadas en congelacion sera motivo de estudios en futuros experimentos de clonacion. Tambien se deben repetir los experimentos en otras especies, incluyendo roedores en donde el ciclo celular es mucho mas rapido, lo que permitira evaluar el efecto del tiempo de activacion del genoma cigotico en la reprogramacion nuclear.
El estudio del ADN mitocondrial heredado basicamente a partir del citoplasto ofrecera luz sobre la viabilidad a largo plazo de animales clonados a partir de individuos adultos, ya que algunas enfermedades relacionadas con desordenes metabolicos tienen su origen en el envejecimiento de las mitocondrias.
Las biotecnologias aplicadas, como la transgenesis, se enriqueceran notablemente si se logra demostrar la repetibilidad de la clonacion somatica, pues permitiran en principio modificar en el laboratorio celulas de la estirpe deseada, evaluar el comportamiento de la proteina secretada por el transgen de interes y solo entonces y con esas celulas ya estudiadas, se llevaria a cabo la clonacion entendiendose por ello la generacion de un individuo clonico y transgenico. Esto es especialmente valido e importante para la transgenesis en animales de granja y sus aplicaciones fundamentales actuales, la expresion de proteinas recombinantes en la leche y el xenotrasplante de organos.
Por otra parte se podrian multiplicar de forma casi indefinida animales de alto valor genetico (transgenicos incluidos) o superproductores en el caso de la ganaderia; si se establece la universalidad de la tecnologia, se podrian quiza preservar celulas de especies en peligro de extincion para restaurar en un futuro las mismas. Como riesgo fundamental, se vislumbra al parecer la clonacion de la susceptibilidad a enfermedades al llevarse a cabo la clonacion de hatos de animales de alta productividad. No queda duda alguna que la clonacion de un individuo adulto a partir de una celula diferenciada ha trascendido las meras fronteras de la procreacion y del nacimiento de una ovejita de raza Finn Dorset de abundante produccion lanar, y llamada Dolly, pues con ella se han roto paradigmas de la biologia que situan a esta ciencia unos milimetros mas adelante en el camino hacia el entendimiento de su propia esencia. Agradecimientos Quiero expresar mi gratitud a Boris Ramos por su apoyo en todo momento, a los doctores Ian Wilmut (Roslin Institute, AFRC, Edimburgo, Reino Unido) y David Wittingham (Medical Research Council, Londres, Reino Unido) por sus comentarios sobre la ginogenesis en el caso de Dolly y Dr. Bill Jefery de la Pennsylvania State University por los comentarios sobre PCNA. References 1. Wilmut I, Schnieke AE, McWhir J, Kind AJ, Campbell KHS. Viable offspring derived from fetal and adult mammalian cells. Nature 1997;385:810-813. 2. Briggs R, King T. Transplantation of living nuclei from blastula cells into enucleated frog eggs. Proc Natl Acad Sci USA 1952;38:455. 3. Illmensee K, Hope PC. Nuclear transplantation in Mus musculus: Developmental potential of nuclei from preimplantation embryos. Cell 1981;23: 9-18. 4. Willadsen SM. Nuclear transplantation in sheep embryos. Nature 1986; 320:63-65. 5. Prather RS, Barnes FL, Simms MM, Robl JM, Eyestone WH, First NL. Nuclear transplantation in the bovine embryo assessment of donor nuclei and recipient oocyte. Biol Reprod 1987;37:859-866. 6. Stice SL, Robl JM. Nuclear reprogramming in nuclear transplantant rabbit embryos. Biol Reprod 1988;39: 657-664. 7. Prather RS, Simms MM, First NL. Nuclear transplantation in early pig embryos. Biol Reprod 1989;41:414-418. 8. Gurdon JB, Laskey RA, Reeves OR. The developmental capacity of nuclei transplanted from keratinized skin cells of adult frogs. J. Embryol. Exp. Morph 1975;34:93-112. 9. Campbell KHS, McWhir J, Ritchie B Wilmut I. Production of live lambs following nuclear transfer of cultured embryonic disc cells. Theriogenology 1994;43:181. 10. Campbell KHS, McWhir J, Ritchie WA, Wilmut I. Sheep cloned by nuclear transfer from a cultured cell line. Nature 1996;380:64-66.
11. Nagy A, Rossant J, Nagy R, Abramow-Newerly W, Roder JC. Derivation of completely cell culture-derived mice from early-passage embryonic stem cells. Proc. Natl. Acad. Sci. USA 1993;90:8424-8428. 12. McGrath J, Solter D. Completion of mouse embryogenesis requires both the maternal and paternal genomes. Cell 1984;37:179-183. 13. Surani, MAH, Barton SC. Development of gynogenetic eggs in the mouse: implications for parthenogentic embryos. Science 1983;222:1034-1036. 14. Surani MAH, Barton SC, Norris, ML. Development of reconstituted mouse eggs suggests imprinting of the genome during gametogenesis. Nature 1984; 308:548-550. 15. Tarkowski A. In vitro development of haploid mouse embryos produced by bisection of one cell fertilized eggs. J. Embryol Exp Morph 1977;38:187-202. 16. Akers M. Lactation. Encyclopedia of Agricultural Science 1994;2:635- 643. 17. Campbell KHS, Loi P, Cappai P, Wilmut I. Improved development to blastocysts of ovine nuclear transfer embryos reconstructed during the presumptive S-phase of enucleated activated oocytes. Biol Reprod 1994;50: 1385-1393. 18. Stewart C. An udder way of making lambs. Nature 1997;385:769-770. Nota anadida a posteriori Posteriormente a la entrega de este trabajo para su revision y edicion, se hizo publica por parte de cientificos del Instituto Roslin y la compania patrocinadora PPL Therapeutics de Edimburgo, Escocia la noticia del nacimiento de otra oveja clonada a partir de celulas somaticas. En este caso Polly (por ser de la raza Poll Dorset) fue obtenida a partir de celulas de piel fetal. Previamente las celulas fueron transfectadas con un gen codificante para una proteina humana no revelada, que ha de ser secretada en la leche de las ovejas clonadas y transgenicas. Este es un importante paso de avance hacia la simplificacion del proceso de transgenesis y, ademas, constituye la primera comprobacion de la veracidad de los resultados publicados por el propio grupo en febrero del presente ano. Copyright 1997 Elfos Scientiae The following images related to this document are available:Photo images[ba97063b.jpg] [ba97063a.jpg] |
| |||||||||

{kind=link}
{kind=link}