 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
LOS POXVIRUS COMO VECTORES VIVOS: UNA PROMESA EN LA VACUNACION Dania Vazquez Blomquist Division de Vacunas, Centro de Ingenieria Genetica y Biotecnologia, apartado postal 6162, La Habana 10600, Cuba. Fax: (53-7) 218070, 336008; E- mail: carlos.duarte@cigb.edu.cu
Recibido en octubre de 1997. Aprobado en noviembre de 1997.
Code Number: BA98001
ABSTRACT Vaccination against smallpox was the beginning of the use of poxviruses as live vectors. The first works in the molecular biology of vaccinia appeared in the 80's. Since that date many poxviruses have been used for vaccination of animals against different infectious diseases. In many cases, the vaccination with recombinant poxviruses confered protection against diseases. Nevertheless, the complications and side effects associated with vaccinia virus immunization led to the search of new variants of poxviruses such as, attenuated vaccinia and avipoxviruses. The last ones, have a very restricted host range, so they do not replicate in mammals but the antigens can be expressed at early steps of the life circle and induce an immune response. NYVAC (derivative of vaccinia) and ALVAC (derivative of canary pox) are two of the attenuated recombinants. They have demonstrated their security in preclinical assays, so phase I trials in humans have already begun. Until now, ALVAC expressing antigens from the human immunodeficiency virus, is the immunogen with the best results in the induction of a CD8^+ T cell response in immunized seronegative individuals. These recombinants have become in a promise for the fight against the acquired immunodeficiency syndrome. Key words: poxviruses, vaccinia, ALVAC, NYVAC, HIV RESUMEN El uso de los poxvirus en la vacunacion contra la viruela humana, abrio el camino para su empleo como vectores vivos en la vacunacion. El exito del virus vaccinia, propicio que en la decada de los 80 comenzaran los trabajos de biologia molecular con este virus. Asi, surgieron diferentes recombinantes que expresaban antigenos de distintos agentes infecciosos. En muchos casos, la inoculacion de estos recombinantes en animales de experimentacion, confirio proteccion contra la enfermedad. Sin embargo, los efectos secundarios asociados a la vacunacion con vaccinia, alentaron la busqueda de variantes atenuadas de este virus y la utilizacion de los avipoxvirus en la vacunacion. Estos ultimos tienen un rango de hospedero restringido a celulas aviares y la replicacion en mamiferos es abortiva, pero son capaces de expresar los antigenos en estadios tempranos del ciclo de vida, lo cual es suficiente para inducir la respuesta inmune contra los mismos. Entre los vectores atenuados se encuentran NYVAC (derivado de vaccinia) y ALVAC (proveniente de una cepa de viruela de canarios). El uso de estos recombinantes en ensayos preclinicos ha demostrado su seguridad y ha permitido la realizacion de ensayos clinicos en humanos. En especial, los recombinantes derivados de ALVAC, que expresan antigenos del virus de inmunodeficiencia humana, se han convertido en una promesa en la lucha contra el sindrome de inmunodeficiencia adquirida. Palabras claves: poxvirus, vaccinia, ALVAC, NYVAC, VIH Introduccion La viruela se considero una enfermedad severa por mas de 2 000 anos, con una mortalidad entre un 20 y un 30 %. Aparentemente originada en la India o en el Asia occidental, se extendio poco a poco, hacia todo el mundo, y de este modo causo epidemias. La caracteristica fundamental de esta enfermedad era la aparicion de eritemas, que podian estar acompanados por fiebre, malestar y otros sintomas (1). Todas las personas eran susceptibles a la adquisicion de la enfermedad, si bien la mortalidad era mayor en ninos y personas de avanzada edad. En los sobrevivientes se presentaban secuelas, como marcas generalmente en la cara, y en pocos casos, ceguera.
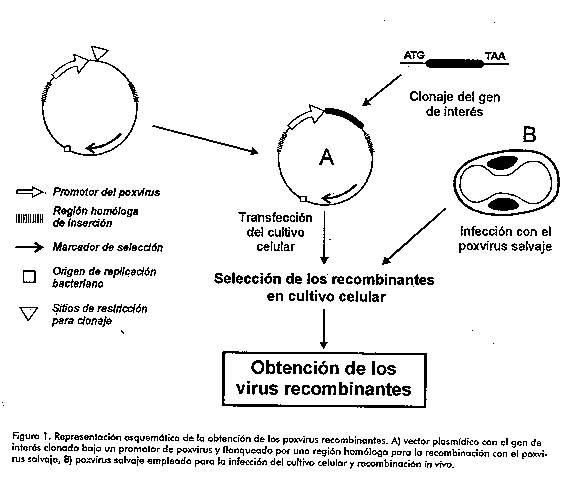
Desde el siglo VI dne los chinos trataron la viruela humana con la inoculacion. No obstante, el primer articulo publicado ("El tratamiento correcto de la viruela humana") le atribuye el merito a una monja budista, practicante durante el reinado de Jen Tsung (1022-1063 dne). Ella recomendaba la seleccion de costras con un mes de aparecidas y en lugares donde el clima es caliente, se recomendaban pustulas de 15 a 20 dias. Estas costras se secaban y se unian con un tipo especifico de planta. La mezcla obtenida, se introducia en la nariz de los pacientes con un tubo curveado de plata (2). Otro texto medico chino (El espejo de oro de la medicina), enumera cuatro formas distintas de inoculacion contra la viruela. Un siglo antes de Jenner, los chinos usaban pulgas blancas de las vacas para la prevencion de la viruela (3). Las pulgas blancas se crecian en un polvo y se hacian pildoras, lo cual quizas fue uno de los primeros intentos de inmunizacion oral. En 1774, en Yeminster, Inglaterra, un ordenador de vacas llamado Benjamin Jesty, inmune a la viruela despues de haber contraido la viruela de las vacas, inoculo a su esposa y dos hijos, a partir de vacas infectadas con la viruela para escapar de la epidemia humana. Gracias a este experimento se mantuvieron protegidos por 15 anos (4). Mas tarde, Jenner reconoceria que esta experiencia de Jesty le sirvio de inspiracion para sus trabajos. Alrededor de esta fecha Edward Jenner, medico ingles, tambien observo casos similares de personas que eran protegidas de la viruela humana por previa exposicion a la viruela vacuna. En una publicacion en 1798 donde expone sus resultados plantea: "...lo que hace a la viruela vacuna tan extremadamente singular es que la persona que ha sido afectada por esta, esta de por vida protegida de la infeccion con viruela humana; ni la exposicion a secreciones contaminadas ni la insercion en la piel del material contaminante producen la enfermedad" (5). En 1796, Jenner inoculo a un nino de 8 anos, James Phipps, con el virus de la viruela vacuna aislado de la mano infectada de Sarah Nelmes, una ordenadora (5). Jenner demostro que este nino fue resistente al reto con el virus de la viruela humana, lo cual fue corroborado por experimentos sucesivos. A partir de ese momento, se conocio este procedimiento como vacunacion. Este procedimiento, a pesar de ser acogido inicialmente con temor, ha sido ampliamente aceptado. Durante los ultimos 200 anos la vacunacion ha controlado nueve de las principales enfermedades, al menos en parte del mundo: la viruela, la difteria, el tetanos, la fiebre amarilla, la tosferina, la poliomelitis, el sarampion, la rubeola y las paperas. En el caso de la viruela se cumplio el sueno de la erradicacion de la enfermedad. El virus de la viruela vacuna fue sustituido posteriormente por el virus vaccinia (VV). Este virus produce una reaccion mas leve en el sitio de inoculacion. Jenner predijo la eliminacion de la viruela humana con la practica de la vacunacion. De acuerdo con esta prediccion, el ultimo caso endemico de viruela ocurrio en 1977 (6) y ya en 1979, la Asamblea de la Organizacion Mundial de la Salud declaro la erradicacion de la viruela humana y recomendo la eliminacion de la vacunacion contra su agente . A principios de la decada del 80 se empezo a aplicar con exito la tecnologia del ADN recombinante para el VV (7, 8). Este virus es la base actual de muchos de los vectores de expresion, candidatos vacunales, y en algunos casos vacunas contra agentes infecciosos heterologos como la rabia, el sarampion, la encefalitis equina, entre otros (9, 10). El uso exitoso del VV recombinante extendio esta tecnologia a otros poxvirus, los cuales han sido empleados en la vacunacion de diferentes especies animales, contra distintas enfermedades infecciosas (11-13) (Tabla 1).
Tabla 1. Poxvirus utilizados como vectores en la vacunacion. El objetivo de esta revision es brindar una vision general del uso de los poxvirus como vectores vacunales contra diferentes enfermedades animales y humanas. Tambien es su proposito exponer las direcciones fundamentales de las investigaciones en la actualidad. Se hace enfasis en la aplicacion de los poxvirus en ensayos preclinicos y clinicos de diversas enfermedades y en especial el sindrome de inmunodeficiencia adquirida (SIDA). Biologia molecular de los poxvirus La familia Poxviridae agrupa una serie de virus envueltos con un genoma de ADN de doble cadena, con replicacion citoplasmatica. Estos son los virus reconocidos como los de mayor genoma, entre 180 a 300 kb. El VV, prototipo del genero Ortopoxviridae, fue el primer virus animal en ser visto microscopicamente, en ser multiplicado en cultivo de tejido, en ser titulado, purificado fisicamente y analizado quimicamente. Presenta una bicapa lipidica que rodea a un nucleo biconcavo con dos cuerpos laterales, cuya funcion es discutida. Mas recientemente, se ha planteado que estos cuerpos laterales aparecen como artefactos de la tecnica usada para su visualizacion (Francisco Rodriguez, comunicacion personal). Las formas extracelulares del virus tienen una bicapa lipidica adicional (14). En el ano 1990, aparecio publicada la secuencia completa de los 191 kb de genoma de la cepa Copenhagen del virus, asi como la funcion de muchos de sus genes (15). Entre las caracteristicas mas relevantes del genoma viral se encuentran la no presencia de intrones y el aprovechamiento al maximo del genoma, aunque es muy raro el solapamiento de los genes. Ambas cadenas codifican para proteinas y los extremos se encuentran unidos covalentemente por un lazo. Se ha podido definir que los genes esenciales para la sobrevivencia del virus se encuentran ubicados hacia el centro del genoma, mientras que hacia los extremos se ubican la mayoria de los genes no esenciales, como los que definen el rango de hospedero. Tanto es asi, que diferentes cepas del VV se pueden diferenciar por deleciones o inserciones que ocurren en los extremos del genoma. El ciclo de vida del VV (14) comienza con la entrada viral y el desnudamiento primario del virus que da paso a la transcripcion de los genes tempranos. Las proteinas que aparecen durante este tiempo son generalmente requeridas para pasos posteriores del ciclo de vida. Unas horas despues el virus comienza a replicarse, y se "apaga" la transcripcion de los genes tempranos, entonces comienza la transcripcion de los genes tardios que, en su mayoria, codifican para proteinas estructurales. Entre las 8 a 12 h ya empiezan a aparecer los primeros viriones, que permanecen intracelularmente o emergen de la membrana celular, para dar las formas extracelulares. Los mecanismos que regulan el paso de la transcripcion de los genes tempranos a los genes tardios, mediados por la replicacion, no son bien conocidos. Se reconocen genes intermedios que se expresan un poco antes del comienzo de la replicacion y existen algunos que se expresan durante todo el ciclo de vida. Del estudio de estos genes, se han definido secuencias consenso para promotores tempranos y tardios, lo cual ha permitido el diseno de promotores sinteticos capaces de provocar altos niveles de transcripcion (16, 17). La definicion de genes no esenciales y de regiones intergenicas ha sido tambien importante en la busqueda de regiones de insercion en el genoma del VV (18). En anos mas recientes, el estudio tambien se ha extendido a los miembros del genero Avipoxviridae, que agrupa virus que infectan a las aves, y que tienen la ventaja potencial de tener un rango de hospedero restringido a celulas aviares, pues la replicacion en mamiferos es abortiva. El virus de la Viruela Aviar (VVA) es el prototipo de este genero. En muchos trabajos se han definido regiones genomicas del VVA con organizacion similar al VV (19-23) y en otros, se han encontrado segmentos con ciertas diferencias. Tal es el caso del gen de la timidinquinasa (TK) el cual esta translocado a otra posicion (24, 25). Se ha comprobado que es posible la transcripcion de genes en el VVA bajo promotores del VV y viceversa, aunque puedan existir diferencias en cuanto a la eficiencia de la transcripcion (26, 27). Obtencion de los vectores poxvirales Para la obtencion de poxvirus recombinantes generalmente se requiere de un plasmidio con una serie de elementos entre los que se encuentran (18):
. Una region promotora insertada en el plasmidio que dirija la transcripcion de los genes foraneos. Estos promotores deben ser originales de poxvirus o sinteticos, con las regiones consenso reconocidas para la expresion temprana y/o la tardia. . Un gen de interes clonado bajo el promotor de poxvirus con su ATG y sin intrones. . Una senal de terminacion de la transcripcion de los genes tempranos en caso de usarse un promotor para la transcripcion temprana. . En algunos casos se puede usar un gen reportero bajo otro promotor de poxvirus, y a continuacion del gen de interes, que permita la seleccion posterior de los recombinantes. . Parte o toda una region no esencial, o intergenica en el genoma original que flanquee los elementos anteriores.
Ventajas y desventajas del uso de los poxvirus como vectores vivos La replicacion citoplasmatica de estos virus, y la codificacion en su genoma de gran parte de las enzimas y factores requeridos para la transcripcion, replicacion y traduccion, son sus mayores ventajas para su uso como vectores. Son capaces de perder gran parte de la informacion (genes no esenciales) sin que se afecte su replicacion in vitro, y aceptar gran cantidad de informacion (hasta 25 kb). La expresion a traves de los vectores poxvirales permite la realizacion de las modificaciones postraduccionales y el trafico intracelular normal de las proteinas, las cuales son presentadas en su configuracion nativa. Esto es importante debido a que muchos anticuerpos neutralizantes estan dirigidos contra epitopos conformacionales que aparecen en la proteina nativa, y que pudieran no aparecer en las variantes recombinantes. Otra de sus ventajas es que con este sistema se puede lograr una buena respuesta citotoxica celular mediada por linfocitos CD8^+, ya que se asegura que el antigeno se procesa en el interior de la celula infectada y los epitopos se presentan en el contexto del Sistema Principal de Histocompatibilidad Tipo I. Esto es un elemento importante a tener en cuenta, en enfermedades en las que es preciso obtener este tipo de respuesta para combatir el agente infeccioso. El VV ha constituido un agente adecuado para la inmunizacion, sus preparaciones son estables, el costo de produccion es bajo, la administracion es facil, la induccion de la inmunidad es de larga duracion y tiene la capacidad de incorporar multiples genes foraneos. A pesar de lo anterior, el amplio uso del VV en la lucha contra la viruela humana, demostro la aparicion, en algunos casos, de encefalitis con desmielinizacion, e infeccion progresiva con el VV, asi como eczema vaccinatum (6, 31). Las complicaciones estuvieron asociadas con la edad, el estado inmunologico de los vacunados y las cepas del VV usadas. Por ejemplo, en Dinamarca las complicaciones aparecieron en aproximadamente 1/4 000 de los vacunados, mientras que en Estados Unidos, con una cepa diferente, las complicaciones aparecieron en 1/250 000 de los vacunados (32). Igualmente, es evidente que la base genetica de la cepa usada afecta la potencia de los recombinantes obtenidos. Un VV recombinante (cepa WR) expresando la proteina gp34 del Virus Epstein Barr (EBV) protegio monos contra el reto con el EBV, mientras que otro VV recombinante (cepa Wyeth) expresando el mismo antigeno fue incapaz de proteger contra un reto identico (33). Recientemente, se reporto la influencia de la cepa del virus parental en la virulencia e inmunogenicidad de un VV recombinante que expresa la proteina preS2-S del virus de la hepatitis B (34). Por otra parte, la previa exposicion de la poblacion al VV puede ser desventajosa cuando se quiere obtener una respuesta inmune contra un antigeno heterologo. Es debido a esto que, actualmente, se explora el uso de otros vectores atenuados derivados del VV y de vectores con rango de hospedero mas restringido, como los avipoxvirus. Aplicaciones de los poxvirus recombinantes en la vacunacion veterinaria Vaccinia Se han reportado numerosos ejemplos del uso del VV recombinante en la vacunacion contra diferentes patogenos, y en muchos casos se ha visto proteccion contra el reto posterior (Tabla 2). En numerosos ejemplos, la proteccion inducida por el VV recombinante se ha correlacionado con los anticuerpos neutralizantes contra proteinas de la envoltura de diferentes virus (95). Sin embargo, en algunos modelos animales la respuesta celular de linfocitos T citotoxicos (CTL) alcanzada, fue la responsable de la proteccion contra el reto viral ( 50, 87, 96, 97). Tabla 2: Vaccinias recombinantes protectores.
---------------------------------------------------------------------------
Patogeno Cepa recombinante Proteccion en animales
---------------------------------------------------------------------------
Rabia Copenhagen Raton, mapache, zorro (35-38)
NYCBH Raton, perro (39)
NYVAC Raton (40)
Sarampion Copenhagen Raton, rata, perro (41-44)
HBV WR Chimpances (45)
HSV WR Raton, curiel (46-49)
MCMV Copenhagen Raton (50, 51)
EBV WR Mono (33)
Influenza WR Hamster, huron, raton (52-55)
MVA Raton (56)
Dengue WR Raton (57, 58)
Fiebre amarilla WR, Copenhagen Raton (59, 60)
JEV WR, Copenhagen Raton (61-63)
NYVAC Cerdo (64)
Virus sincicial respiratorio WR Rata, raton, chimpances (65-67)
HPV Copenhagen Raton, rata (68-70)
Parainfluenza humana WR Mono (71)
Fiebre lassa Wyeth, Lister Curiel (72, 73)
Peste bovina WR, Wyeth,
LC16mo Conejo, reses (74-78)
Virus equino del herpes Copenhagen Hamster (79)
Virus de la pseudorrabia Copenhagen Raton, cerdo (80)
EIV NYVAC Caballo (81)
Virus de leucemia bovina Copenhagen, WR,
LC16mo Oveja (82-84)
Virus del papiloma bovino Copenhagen Rata (85)
Virus polioma Copenhagen Rata (86)
Virus de leucemia Friend WR Raton (87)
Virus colera del cerdo WR Cerdo (88)
VEE Wyeth Raton, mono, caballo (89-91)
Virus Sendai WR Raton (92)
Influenza aviar Wyeth, WR Pollo (32, 93, 94)
--------------------------------------------------------------------------
HBV: Virus de la hepatitis B, HSV: Virus del herpes simplex, MCMV. Virus de
la coriomeningitis murina, EBV: Virus Epstein Barr, JEV: Virus de la
encefalitis japonesa, HPV: Virus papiloma humano, EIV: Virus de la
influenza equina, VEE: Virus de la encefalomiocarditis equina
venezolana.Vaccinia salvaje. La cepa del VV mas ampliamente usada ha sido la WR, cepa altamente virulenta, por lo que su uso como vacuna veterinaria o humana no representa una buena opcion. Otras cepas alternativas a la WR han sido la NYCBH y la Wyeth (DRYVAX). La cepa Copenhagen se ha usado ampliamente en Europa (35), y es muy efectiva como vehiculo de inmunizacion. Cepas atenuadas de vaccinia. Mas recientemente, la cepa MVA ha sido propuesta para su desarrollo como candidato vacunal (98-100). Esta ultima se derivo de la cepa Ankara por mas de 570 pases en cultivos primarios de fibroblastos de embrion de pollo, lo cual resulto en la delecion de regiones del genoma que han alterado el rango de hospedero de la cepa, y la perdida de su capacidad replicativa en cultivos de celulas no aviares (101). Esta cepa, al igual que las virulentas, mostro una inmunogenicidad elevada en ratones (100). Otra cepa, la LC16m8 derivada de la cepa del VV Lister, presento una neurovirulencia reducida aunque no tan atenuada como la MVA. Su uso como vacuna recombinante confirio inmunidad a los animales inmunizados (102). La insercion de genes de linfocinas en algunas regiones del genoma del VV, disminuye la virulencia del mismo, sin que se afecte la inmunogenicidad. Los virus recombinantes que expresan la IL2 (103) o IFN gamma (104) humanos, o de raton, son menos patogenicos para ratones atimicos desnudos inmunodeficientes, que los virus originales. El fenotipo del VV tambien ha sido atenuado por la delecion especifica de ciertos genes que codifican para TK (105), factor de crecimiento (106), hemaglutinina (102, 103), ribonucleotido reductasa (107), proteinas de la envoltura (108), proteinas para el control del complemento (109), y varios genes que controlan el rango de hospedero (110). En 1992, Tartaglia y cols (111) describieron la obtencion de un VV altamente atenuado, y mas seguro, al que llamaron NYVAC y se derivo de la cepa Copenhagen del VV por la delecion precisa de 18 marcos de lecturas, codificantes para funciones vinculadas con determinantes de patogenicidad, rango de hospederos y replicacion. Entre sus principales ventajas se encuentran (40, 111):
. Habilidad de replicarse con alta eficiencia en cultivos primarios de fibroblastos de embrion de pollo. . Ausencia de endurecimiento y ulceracion en el sitio de inoculacion en la piel de conejos. . Eliminacion rapida del virus infeccioso en el sitio de inoculacion intradermico en piel de conejos. . Ausencia de inflamacion testicular en ratones desnudos. . Virulencia altamente reducida despues de un reto intracraneal, tanto en ratones de 3 semanas como recien nacidos. . Patogenicidad altamente reducida y fallo en la diseminacion de la infeccion, en ratones inmunocomprometidos. . La atenuacion del NYVAC hizo que el Comite Supervisor del ADN Recombinante del Instituto Nacional de Salud de los Estados Unidos, redujera el nivel de contencion biologica para su manipulacion, de BSL2 a BSL1 (112). Este es el unico ortopoxvirus que se puede trabajar bajo estas condiciones.
La inoculacion con NYVAC-JEV, que expresa la proteina preM y proteinas de la envoltura del virus de la encefalitis japonesa (JEV), protegio cerdos contra el reto con JEV. Este inmunogeno fue capaz de generar anticuerpos neutralizantes y anticuerpos inhibidores de la hemaglutinacion (64). La expresion de las hemaglutininas (HA) tipo A1 y A2 del virus de la influenza equina (EIV) por NYVAC, protegio caballos contra el reto con EIV tipo A2 y desarrollo solo sintomas atenuados de la enfermedad. En este estudio se detectaron anticuerpos inhibitorios de la hemaglutinacion (81). En otro experimento, donde se inocularon cerdos con NYVAC, expresando las glicoproteinas gB y gD del virus de la pseudorrabia (PRV), estos se protegieron al ser retados con PRV (113, 114). En articulos mas recientes, se ha ensayado un NYVAC que codifica para el antigeno de superficie del HBV, y una proteina de fusion, en la que preS1 y preS2 estan fusionadas al N-terminal del antigeno de la capsida viral. La inoculacion con este vector indujo una respuesta inmune alta contra todos los antigenos, en animales de laboratorio (115). De igual forma, los genes para las proteinas gp34, gp110 o gB, y gp85 del EBV y antigenos del HBV, se combinaron en una sola preparacion vacunal basada en NYVAC. Esta forma de expresion de los antigenos fue capaz de inducir una respuesta de anticuerpos, contra el EBV y el HBV (115). Estos ejemplos demuestran que NYVAC retiene la inmunogenicidad y la potencia de un vector del VV replicativo, y representa una opcion mas segura respecto a las cepas existentes. Avipoxvirus Aunque los avipoxvirus recombinantes no se replican en celulas de mamiferos, existe una sintesis de novo de las proteinas recombinantes, que induce una respuesta inmune contra la misma. En un experimento muy ilustrativo, animales inoculados con VVA recombinante para la glicoproteina de la rabia, generaron anticuerpos contra este virus. Sin embargo, animales inoculados con un virus recombinante inactivo, produjeron anticuerpos contra el vector viral pero no contra el antigeno del virus de la rabia. Esto indica que la respuesta inmune especifica contra el virus de la rabia, fue inducida por las proteinas sintetizadas de novo en las celulas infectadas y no por las proteinas introducidas al inocular el virus (116). Virus de la Viruela Aviar. El VVA es el prototipo de este genero y desde 1920 se ha usado una vacuna atenuada con el VVA para su control en aves (117). La eficacia de este virus como vector para la inmunizacion, fue demostrada en 1988 por Taylor y cols (116, 118), tanto en especies aviares como no aviares. Especialmente en especies aviares, la inmunizacion con VVA que expresa diferentes antigenos, ha sido eficaz para combatir multiples enfermedades. La inoculacion de pollos y pavos con VVA, que expresa la HA del virus de la influenza aviar, los protegio contra un reto con el virus salvaje (119). La inmunizacion con VVA, recombinante para la HA neuraminidasa (120-122) y la proteina de fusion F (123, 124) del virus Newcastle protegio a pollos contra un reto viral (125). La coexpresion de ambas proteinas tambien ha inducido una respuesta inmune capaz de proteger a pollos contra el reto viral (122). El uso de VVA recombinantes, para la proteina gB del virus Marek s, protegio a pollos contra la aparicion del linfoma caracteristico de esta enfermedad, y contra la muerte, ante el reto con el virus salvaje (126). Este virus tambien se ha usado para combatir la leucosis aviar y el virus sarcoma (127). La utilidad de los avipoxvirus en la inmunizacion de mamiferos se demostro con el uso de un VVA recombinante que expresa la glicoproteina del virus de la rabia. La inoculacion con este recombinante protegio a gatos, perros y ratones ante el reto con el virus salvaje (116). En 1993, una patente presentada por la compania Virogenetics (128), describio la obtencion de un VVA modificado geneticamente a partir de una placa aislada de VVA, al que llamaron TROVAC. En 1994, Radaelli y cols (129) reportaron la inoculacion de conejos con este vector, recombinante para el virus de la inmunodeficiencia humana.
ALVAC. Tartaglia y cols, en 1992 (111), describieron tambien la obtencion de un virus altamente atenuado, a partir de la cepa vacunal del virus de la viruela en canarios, al que llamaron ALVAC. En analogia al NYVAC, ALVAC exhibe una menor patogenicidad en ratones recien nacidos e inmunocomprometidos. La respuesta inmune inducida por un VVA que expresa la glicoproteina de la rabia, no fue tan potente como la inducida por el VV-RG. Sin embargo, el ALVAC-RG (virus de la viruela en canarios recombinante para la glicoproteina de la rabia) indujo una respuesta inmune tan potente como el VV-RG. En un estudio de reto con el virus de la rabia (40), el ALVAC-RG tuvo una PD50 de 3,86, similar al VV-RG (3,74) y al NYVAC-RG (3,7). Las razones de las diferencias entre ambos avipoxvirus no son conocidas. ALVAC- RG protegio a perros y gatos contra la rabia (130). Once de doce perros resistieron el reto con el virus de la rabia despues de 36 meses de la inoculacion con ALVAC-RG, a pesar de no detectarse anticuerpos neutralizantes antes del reto (131). Perros inmunizados con ALVAC recombinante para la HA, y el antigeno F del sarampion desarrollaron anticuerpos neutralizantes. Estos animales resistieron el reto con un virus salvaje, de forma similar a los perros inoculados con un VV replicativo, expresando los mismos antigenos. Los perros inmunizados con este preparado contra sarampion, mostraron una respuesta de anticuerpos neutralizantes y fueron protegidos contra el reto (132) con el virus del moquillo canino (Morbillivirus especifico de perros, semejante al del sarampion). Los hurones han sido usados recientemente como modelo animal para investigar la proteccion contra el moquillo canino. Virus recombinantes basados en ALVAC y NYVAC han protegido a estos animales contra la infeccion sintomatica (133). La inoculacion con ALVAC, que expresa las HA de tipo 1 y 2 de EIV protegio a caballos contra el EIV. Una sola inoculacion indujo una respuesta de anticuerpos inhibitorios de la hemaglutinacion contra A1 y contra A2 (81). Los sintomas fueron atenuados respecto a los provocados por la infeccion con el virus salvaje. Se midieron altos titulos de anticuerpos contra la HA tipo A2 en los caballos retados con un virus con este subtipo de HA. Por otra parte, la inmunizacion con ALVAC- FeLV, que expresa las proteinas Gag y Env del virus de la leucemia felina, protegio al 100 % de los gatos contra el reto oronasal con el virus salvaje. Cuando se le deleciono una region inmunosupresora a la proteina Env, los recombinantes ALVAC- FeLV protegieron al 50 % de los gatos. El analisis serologico demostro que ninguno de los 12 gatos desarrollaron anticuerpos neutralizantes antes del reto con el virus, pero todos los gatos protegidos generaron anticuerpos neutralizantes despues del reto (134). Estos resultados sugieren que la proteccion esta asociada a una respuesta inmune primaria que es rapidamente amplificada una vez retados los animales. No obstante, la respuesta celular puede jugar un papel importante en la proteccion. Ratones inoculados con ALVAC-JEV, que expresa las proteinas preM, E, NS1 de JEV fueron protegidos ante el reto con JEV (135). Aplicacion de los poxvirus en la vacunacion humana La historia de la vacunacion en humanos con estos virus, comenzo con la inoculacion contra la viruela. A partir de los anos 80 se han probado diferentes vectores, en animales de laboratorio. Algunos de estos se encuentran en la fase preclinica, como un candidato vacunal basado en ALVAC contra el citomegalovirus humano (136), y otros han llegado a ensayos clinicos en humanos. Uno de esos ejemplos es la vacuna contra la rabia, ALVAC-RG ensayada en humanos (Fase I) en Francia y Estados Unidos. Con esta vacunacion se obtuvo una respuesta de anticuerpos neutralizantes y respuesta celular contra el virus de la rabia (137, 138). El ALVAC que expresa la HA y la proteina de fusion del sarampion esta siendo evaluado en ensayos clinicos en humanos. Igualmente, los vectores NYVAC-JEV y ALVAC-JEV estan siendo probados en ensayos clinicos (Fase I) en humanos. Entre los ejemplos mas notables encontramos los ensayos de Fase I con ALVAC expresando proteinas del virus de inmunodeficiencia humana tipo 1 (VIH-1), vCP125 y vCP205 (139-142); los cuales seran discutidos con posterioridad. Por ultimo, existen reportes del uso de NYVAC para combatir enfermedades parasitarias como la malaria (143). Actualmente esta en curso un estudio de Fase I en humanos con un vector NYVAC, que expresa proteinas del Plasmodium falciparum. La alta inmunogenicidad de este preparado fue demostrada originalmente en macacos (144). Poxvirus recombinantes y el desarrollo de vacunas contra el VIH Debido a las caracteristicas peculiares del VIH como agente etiologico del SIDA se han ensayado multiples estrategias vacunales para su prevencion. Entre estas encontramos: el uso de la proteina de la envoltura viral (gp160 o gp120) obtenida por via recombinante, peptidos sinteticos, adenovirus recombinantes, poxvirus recombinantes, entre otros (145). Una de las caracteristicas del SIDA es que no se conoce que tipo de respuesta inmune es necesaria para lograr proteccion contra el virus, al no existir un modelo animal adecuado que reproduzca la enfermedad ante el reto con el VIH. Vaccinia salvaje El VV ha sido ampliamente usado para expresar un gran numero de productos genicos del VIH, con el objetivo de investigar sus propiedades bioquimicas, funcionales e inmunologicas. En relacion con el desarrollo de vacunas, los VV recombinantes para genes de VIH han sido ampliamente usados para caracterizar la respuesta inmune; particularmente la respuesta CTL (95), y para inducir una respuesta humoral y celular en diferentes modelos animales como macacos y chimpances; tambien en ensayos clinicos en humanos (145). La inmunizacion de ratones con algunos de estos recombinantes, ha inducido respuesta humoral. En especial, recombinantes que expresan la glicoproteina de la envoltura (146, 147) han inducido respuesta de anticuerpos neutralizantes. Existen algunos trabajos donde se protegen macacos al ser inoculados con el VV que expresan genes del virus de inmunodeficiencia de simios (VIS), analogo al VIH (148, 149). Macacos inmunizados con dosis iniciales de VV recombinante que expresa la glicoproteina de la envoltura de VIS y reactivados con una preparacion recombinante de la glicoproteina de la envoltura del SIVmne fueron protegidos contra un reto con el virus SIV (aislamiento MNE) (149). Sin embargo, la inmunizacion de macacos con VV, que expresan diferentes proteinas de VIH-2, como Gag, Pol, Vif, Nef, Env, seguido de una reactivacion con proteinas purificadas de la cepa ROD de VIH-2, no fueron capaces de proteger a los macacos contra un reto con la cepa SBL de VIH-2 (150). Lo anterior sugiere que la respuesta inmune inducida contra la cepa vacunal fue dirigida principalmente contra epitopos no conservados en la cepa que se empleo en el reto. Como resultado de la expresion del gen gag en las celulas infectadas con VV, se producen particulas virales semejantes al VIH pero de caracter no infeccioso (VLP) (95, 151). La produccion in vivo de estas VLP pudiera aumentar la inmunogenicidad de los VV, pero como su formacion depende del tipo celular (151), no se conoce si se producen con la administracion in vivo. Un VV recombinante (HIVAC-1e) de las companias Oncogen/Bristol-Myers Squibb, que expresa la glicoproteina de la envoltura de VIH-1 (aislamiento IIIB), ha entrado en Fase I en humanos (152-154). Los resultados con esta preparacion sola, o en combinacion con la proteina gp160 recombinante expresada en baculovirus, son los siguientes:
2. Los regimenes de inmunizacion en los que vacunan primeramente con HIVAc- 1e y amplifican la respuesta inmune con la gp160 recombinante adyuvada en alumina, han dado mejores resultados que cuando se usan los inmunogenos por separado. 3. Los regimenes combinados dieron como resultado una respuesta citotoxica especifica contra VIH-1, de tipo CD4^+ y CD8^+, mientras que los inmunizados con las subunidades recombinantes solo exhibieron una respuesta celular de tipo CD4^+. 4. La respuesta inmune fue mas pronunciada en individuos virgenes para el VV, en comparacion con individuos con una respuesta previa contra el virus.
Por ultimo, un VV recombinante que expresa la proteina de la envoltura de VIH-1 de la cepa IIIB, y las proteinas Gag y Pol esta en estudio en humanos (145). Con este candidato se realizo un ensayo clinico en individuos inmunes a vaccinia, llevado a cabo por la compania Therion. Se planea un proximo estudio, en individuos virgenes para vaccinia, a fin de comparar la respuesta inmune con respecto al experimento anterior. NYVAC y ALVAC en animales Abimiku y cols, en 1995 (155), inmunizaron macacos con NYVAC o ALVAC, que expresaban las proteinas de la envoltura viral del VIH-1: gp120 o gp160 de las cepas MN o IIIB. La reactivacion de la respuesta inmune se realizo con peptidos octamericos, gp120 nativa de la cepa IIIB u otra subunidad proteica, adyuvados en alumina. El reto intravenoso con VIH-2 protegio a 3/8 macacos. Franchini y cols, en 1995 (156), inmunizaron macacos con alguno de los siguientes inmunogenos: NYVAC o ALVAC que expresaban los genes env, gag, pol de VIH-2; NYVAC que expresaba gp160 o gp120 de VIH-2 en combinacion con gp160 recombinante de VIH-2 adyuvada en alumina. El reto intravenoso con VIH-2 protegio a 7/8 macacos. De los siete protegidos solo cinco de ellos siguieron protegidos al ser retados por segunda vez, seis meses despues. Lo anterior demuestra la duracion de la respuesta inmune. Biberfield y cols, en 1996 (157), usaron varios esquemas de inmunizacion en macacos. Los inmunogenos usados fueron: ALVAC recombinante para las proteinas de la envoltura, Gag y Pol de VIH-2 o ALVAC que expresaba la proteina de la envoltura de VIH-2 solamente. En estos esquemas se administro el virus solo o en combinacion con la proteina de la envoltura viral de VIH-2 adyuvada en QS21, o con un peptido sintetico en el mismo adyuvante. En general, se indujo una respuesta de anticuerpos neutralizantes pobre, pero alta respuesta linfoproliferativa en los grupos donde se uso la combinacion con la glicoproteina. Solo tres de doce macacos mostraron una respuesta citotoxica. A pesar de que el 40 % de los macacos fueron protegidos contra el reto con VIH-2, en los grupos donde se amplifico con la glicoproteina no se encontro una correlacion entre la respuesta inmune y la proteccion. Las diferencias encontradas en los trabajos de Abimiku y cols (155) y Franchini y cols (156), se deben a diferencias en la procedencia de los inmunogenos usados y los virus empleados para el reto viral. En el primer caso, se usan inmunogenos con proteinas de VIH-1, mientras que el reto se realiza con VIH-2. En el segundo trabajo, la inmunizacion es con proteinas de VIH-2 y el reto con este mismo virus. Tanto el VIH-1 como el VIH-2, son los agentes etiologicos del SIDA, pero existen diferencias genomicas y epitopicas entre ambos virus; tanto es asi, que el VIH-2 guarda un mayor por ciento de homologia con varias cepas de VIS, que con el propio VIH-1. Ademas, los macacos como modelo animal, tienen la desventaja que el VIH-1 no es capaz de infectarlos, por lo que hay que usar un modelo alternativo como el VIH-2 para llegar a conclusiones, que en algunos casos pueden no ser extrapolables a humanos. De cualquier manera estos trabajos corroboran en macacos, que se requieren regimenes combinados de inmunizacion, con el virus recombinante y un inmunogeno de naturaleza proteica, para lograr proteccion contra la infeccion (158). Girard en 1995 (159) reporto la inoculacion de dos chimpances con altas dosis (4 x 10^8 unidades formadoras de placas) de ALVAC-HIV-1(vCP205). Este recombinante expresa la gp120 de la cepa MN, la proteina Gag y la proteasa de la cepa LAI. Los chimpances fueron retados intravenosamente con linfocitos de sangre periferica infectados con la cepa IIIB de VIH-1. Se indujeron anticuerpos de union a la region variable 3 (V3) de la gp120 y neutralizantes, 3-4 veces mayor en magnitud en uno de los chimpances con respecto al otro, en el momento del reto. El chimpance que mostro mayores titulos de anticuerpos fue protegido contra el reto viral. En un reporte previo (160) se inocularon dos chimpances con vCP125, recombinante que solo expresa gp160 de la cepa MN, y se amplifico la respuesta inmune con gp160MN/LAI (gp120 de MN y gp41 de LAI). Este regimen de inmunizacion no los protegio contra el reto con VIH-1 de la cepa SF2. La explicacion que le dan los autores a este resultado es que la dosis utilizada era muy baja (10^6.1 unidades formadoras de placas). Es de senalar que los experimentos con chimpances, al ser con tan pocos animales, pueden no dar una informacion definitiva al comparar diferentes inmunogenos. Ademas, en este modelo animal solo se puede investigar la proteccion contra la infeccion viral inicial, pues los chimpances no desarrollan SIDA, al menos en un tiempo corto. En este modelo animal no es posible medir la proteccion contra el desarrollo de la enfermedad. Radaelli y cols, en 1994 (129), usaron los recombinantes del VVA, vFP62 y vFP63, que expresan la gp160 de SF2. Se les identifico como FPIS^- y FPIS^+ y se les comparo con los recombinantes a partir del virus de la viruela en canarios, que expresan la misma proteina, llamados CPIS^- y CPIS^+. Los anticuerpos presentes en los sueros de los conejos inmunizados con CPIS^-, CPIS^+ y FPIS^-, neutralizaron la cepa SF2 in vitro . Sin embargo, no se detecto actividad neutralizante en los sueros de los inmunizados con FPIS^+. Los experimentos de neutralizacion cruzada de las cepas de laboratorio MN, IIIB, o RF mostraron que, ninguno de los sueros, tenian anticuerpos capaces de neutralizar las mismas. Se detecto una respuesta de linfocitos T, en todos los animales. Los titulos de anticuerpos, y la actividad neutralizante de los mismos, fueron mayores en los conejos inoculados con los virus de viruela en canarios con respecto a los inoculados con los VVA. Estas diferencias en cuanto a respuesta inmune, entre ambos recombinantes aviares, se pueden deber a que la transcripcion de los genes o la sintesis proteica pueda ser mas eficiente en un virus con respecto a otro. El modelo animal empleado puede tambien influir en los resultados obtenidos. Explicaciones adicionales, no se pueden descartar. ALVAC en humanos Los mejores resultados en ensayos de Fase I, en cuanto a respuesta celular y humoral, se han obtenido con los candidatos vacunales basados en ALVAC. El virus recombinante vCP125, expresa la gp160 de la envoltura de VIH-1 y el recombinante vCP205, expresa la gp120 de la cepa MN, la proteina Gag, la proteasa y la region transmembranica de la cepa IIIB. Estos virus se han usado en regimenes combinados con un inmunogeno proteico recombinante, generalmente gp120 o gp160. Algunos resultados se exponen a continuacion. El primer articulo aparecio publicado en 1995 (139), si bien en la X Conferencia Internacional sobre SIDA, en Japon (1994) se habian presentado ya algunos datos alentadores en cuanto al uso de ALVAC como vector vacunal contra el VIH. En el mismo, se uso un regimen combinado de inmunizacion con vCP125 y la gp160 MN/LAI recombinante adyuvada en alumina o Montanide ISA51 (adyuvante oleoso semejante al Incompleto de Freund). El 90 % de los individuos inoculados desarrollaron anticuerpos neutralizantes contra la cepa MN, y de estos solo el 50 % desarrollaron anticuerpos neutralizantes contra SF2, cepa muy parecida en su secuencia aminoacidica al MN. Sin embargo, no se detectaron anticuerpos neutralizantes contra la cepa IIIB, cepa mas divergente que la SF2, ni contra aislamientos virales primarios, provenientes de pacientes infectados. Se obtuvieron lineas celulares de CTL de las cuales el 39 % estaban dirigidas contra epitopos presentes en la proteina de la envoltura. La mayoria de la respuesta CTL era debida a linfocitos T CD8^+ (64-90 %) y solo el 4 % a CD4^+. Clements y cols, 1996 (140), en la XI Conferencia Internacional sobre SIDA, en Vancouver, expusieron los resultados de la combinacion de vCP125 con gp120 (Compania Chiron) recombinante de la cepa SF2 adyuvada a MF59. Los resultados expuestos en ese evento se presentan en la Tabla 3. Tabla 3. Resultados de la combinacion de vCP125 con gp120 recombinante de la cepa SF2.
-----------------------------------------------------------------------
Esquema Neutralizacion contra CTL CTL LP
MN(%) SF2(%) CD8^+(%) CD4^+(%) (%)
-----------------------------------------------------------------------
4XALVAC 10^6 4/10(40) 2/10(20) 10 10 86
2XALVAC10^6+2Xrgp120 9/9(100) 9/9(100) 25 25 100
4XALVAC 10^7 12/18(67) 2/18(11) 17 17 50
2XALVAC10^7+2Xrgp120 29/29(100) 28/28(100) 42 21 80
4Xrgp120 5/9(56) 8/8(100) 10 0 -
-----------------------------------------------------------------------
CTL: Respuesta celular citotoxica, LP: Respuesta linfoproliferativa,
rgp120:gp120 recombinante, 4X y 2X: 4 y 2 inoculaciones.En general, se logro una respuesta de anticuerpos neutralizantes, CTL y LP. Los esquemas con inmunizacion inicial con vCP125 (sobre todo el de mayor dosis) y la amplificacion con gp120 recombinante indujeron una respuesta de anticuerpos neutralizantes y CTL mas frecuentemente que los otros esquemas. Lawrence y cols (142), investigaron la inmunogenicidad del virus recombinante vCP205 solo o en combinacion con gp120 recombinante de la cepa SF2 (Compania Chiron). Solo el 10 % de los individuos mostraron anticuerpos contra peptidos de VIH-1, contra proteinas purificadas o contra lisados de celulas infectadas con el virus. Se detecto actividad CTL (CD8^+) Gag- dependiente en el 70 % de los individuos, mientras solamente el 12 % mostro actividad CTL contra epitopos presentes en la proteina de la envoltura. Recientemente se anuncio el comienzo de estudios de Fase II en humanos con el virus recombinante, vCP205 (Jose Esparza, OMS, comunicacion personal). Consideraciones finales El papel jugado por el VV en la erradicacion de la viruela humana sugirio que los poxvirus pueden tener un uso importante como vectores vivos en la vacunacion. En el campo veterinario algunas vacunas basadas en poxvirus han sido licenciadas para su uso y se comercializan. En humanos se dan los primeros pasos, basados en los resultados alentadores obtenidos con la vacuna contra la rabia. En algunas enfermedades infecciosas sin cura hasta este momento, como el SIDA, el uso de recombinantes que expresan antigenos del VIH-1 han mostrado resultados promisorios en experimentos de Fase I en humanos. Los problemas concernientes a la seguridad del VV se han eliminado con el desarrollo de los vectores atenuados, ALVAC y NYVAC. Los mismos, pueden expresar multiples genes del mismo patogeno o de diferentes patogenos. Actualmente, el estudio de estos vectores para su uso como vacuna en humanos y en animales sigue avanzando y se encuentra en la mirilla de la investigacion cientifica. Sin embargo, esta por demostrar que la respuesta inmune, tanto humoral como celular, inducida en humanos sea capaz de protegerlos contra los agentes infecciosos o el desarrollo de las enfermedades. Esta respuesta unicamente la daran el tiempo y la experimentacion. Agradecimientos Quisiera agradecer al licenciado Rolando Pajon y al doctor Carlos Duarte por la revision critica de este articulo. References 1. Fenner F. Poxviruses. In: Fields BN, Knipe DM, editors. Virology, New York: Raven Press, 1990:2113-2133. 2. Hume EH. The Chinese way in medicine. Baltimore: Johns Hopkins Press, 1940. 3. Wong KC, Wu LT. History of Chinese medicine. Tientsin Press, 1932. 4. Parish HJ. A history of immunization. London: E&S Livingstone, 1965. 5. Jenner E. An inquiry into the causes and effects of the variole vaccine or cowpox. In: Charles W. Eliot, LL.D, editor. The Harvard Classics, New York: Collier & Son corporation, 1938;38:145-223. 6. Fenner F, Henderson DA, Arita I, Jezek Z, Ladnyi ID. Smallpox and its eradication. Geneva, World Health Organization. 1988: 1460. 6. Wittek R, Moss B. Tandem repeats within the inverted terminal repetition of vaccinia virus DNA. Cell 1980;21:277-284. 8. Wittek R, Cooper J, Barbosa E, Moss B. Expression of vaccinia virus genome: analysis and mapping of mRNAs encoded within the inverted terminal repetition. Cell 1980;21: 487-493. 9. Panicali D, Paoletti E. Construction of poxviruses as cloning vectors: insertion of the thymidine kinase gene from herpes simplex virus into the DNA of infectious Vaccinia virus. Proc Nat Acad Sci USA 1982;79:4927- 4931. 10. Mackett M, Smith GL, Moss B. Vaccinia virus: a selectable eukaryotic cloning and expression vector. Proc Nat Acad Sci USA 1982;79:7415-7419. 11. Letellier C, Burny A, Meulemans G. Construction of a pigeonpox virus recombinant: expresion of the NDV fusion glycoprotein and protection of chickens against NDV challenge. Arch Virol 1991;118:43-56. 12. Wasmoen TL, Kadakia NP, Unfer RC, Fickbohm BL, Cook CP Chu HJ, Acree WM. Protection of cats from infectious peritonitis by vaccination with a recombinant raccoon poxvirus expressing the nucleocapsid gene of feline infectious peritonitis virus. Adv Exp Med Biol 1995;380:221-228. 13. Hu L, Esposito JJ, Scott FW. Raccoon poxvirus feline panleukopenia virus VP2 recombinant protects cats against FPV challenge. Virology 1996;218:248-252. 14. Moss B. Poxviridae and their replication. In: Fields BN, Knipe DM, editors. Virology, New York: Raven Press, 1990:2079-2111. 15. Goebel SJ, Jonhson GP, Perkus ME, Davis SW, Winslow JP, Paoletti E. The complete DNA sequence of vaccinia virus. Virology 1990;179:247-266. 16. Davinson AJ, Moss B. Structure of vaccinia virus early promoters. J Mol Biol 1989;210: 749-769. 17. Davinson AJ, Moss B. Structure of vaccinia virus late promoters. J Mol Biol 1989; 210:771-784. 18. Hruby DE. Vaccinia virus vectors: new strategies for producing recombinant vaccines. Clinical Microbiology Reviews 1990; 3:153-170. 19. Drillien R, Spehner D, Villeval D, Lecocq JP. Similar genetic organization between a region of fowlpox virus DNA and the vaccinia virus HindIII J fragment despite divergent location of the thymidine kinase gene. Virology 1987;160:203-209. 20. Binns MM, Tomley FM, Campbell J, Bournsell MG. Comparison of a conserved region in fowlpox virus and vaccinia virus genomes and the translocation of the fowlpox virus thymidine kinase gene. J Gen Virol 1988;69:1275-1283. 21. Binns MM, Britton BS, Mason C, Boursnell MG. Analysis of fowlpox virus genome region corresponding to the vaccinia virus D6 to A1 region: location of, and variation in, non-essential genes in poxviruses. J Gen Virol 1990;71:2873-2881. 22. Tartaglia J, Winsloww J, Goebel S, Johnson GP, Taylor J, Paoletti E. Nucleotide sequence analysis of 10.5 kbp HindIII fragment of fowlpox virus: relatedness to the central portion of the vaccinia virus HindIIID region. J Gen Virol 1990;71:1571-1524. 23. Zantinge JL, Krell PJ, Derbyshire JB, Nagy E. Partial transcriptional mapping of the fowlpox virus genome and analysis of EcoRI L fragment. J Gen Virol 1996;7:603-614. 24. Weir JP, Moss B. Nucleotide sequence of the vaccinia virus thymidine kinase gene and the nature of spontaneous frameshift mutations. J Virol 1983;46:530-537. 25. Boyle DB, Couper BEH, Gibbs AJ, Seigman LJ, Both GW. Fowlpox virus thymidine kinase: nucleotide sequence and relationships to other thymidine kinases. Virology 1987;156:355-365. 26. Tripathy DN, Wittek R. Regulation of foreign gene in fowlpox virus by a vaccinia virus promoter. Avian Diseases 1990;34:218-220.
27. Kumar S, Boyle DB. Activity of fowlpox virus late gene promoter in vaccinia and fowlpox virus recombinants. Ach Virol 1990;112:139-148. 28. Nazerian K, Dhawale S. Structural analysis of unstable intermediate and stable forms of recombinant fowlpox virus. J Gen Virol 1991;72:2791- 2795. 29. Scheiflinger F, Dorner F, Falkner FG. Construction of chimeric vaccinia viruses by molecular cloning and packaging. Proc Nat Acad Sci USA 1992;89:9977-9981. 30. Pfleiderer M, Falkner FG, Dorner F. A novel vaccinia virus expression system allowing construction of recombinants without the need for selection markers, plasmids and bacterial hosts. J. Gen Virol 1995;76:2957-2962.
31. Lane JM, Ruben FL, Neff JM, Millar JD. Complications of smallpox vaccination, 1968. National surveillance in the United States. New Engl J Med 1969;281:1201-1208. 32. Neff J. Vaccinia virus vaccines: virulence and attenuation of vaccinia strain variation. In: Quinnan G, editor. Vaccinia virus as vectors for vaccine antigens, New York: Elsevier Science Publishing Co, 1985:69-75. 33. Morgan AJ, Mackett M, Finerty S, Arrand JR, Scullion FT, Epstein MA. Recombinant vaccinia virus expressing Epstein Barr virus glycoprotein gp34 protects cottontop tamarins against EB virus induced malignant lymphomas. J Med Virol 1988;25:189-195. 34. Kutinova L, Ludvikova V, Krystofova J, Otavova M, Simonova V, Nemeckova S et al. Influence of the parenteral virus strain on the virulence and immunogenicity of recombinant vaccinia viruses expressing HBV preS2-S protein or VZV glycoprotein. Vaccine 1996; 14:1045-1052. 35. Brochier B, Kieny MP, Costy F, Coppens P, Bauduin B, Lecocq JP et al. Large-scale eradication of rabies using recombinant vaccinia-rabies vaccine. Nature 1991;354: 520-522. 36. Kieny MP, Lathe R, Drillien R, Spehner D, Skory S, Schmitt D et al. Expression of rabies glycoprotein from a recombinant vaccinia virus. Nature 1984;312:163-166. 37. Ruprecht CE, Wiktor TJ, Johnston DH, Hamir AN, Dietzchold B, Wunner WH, Glickman LT, Koprowski H. Oral immunization and protection of raccoons (Procyon lotor) with a vaccinia-rabies glycoprotein recombinant virus vaccine. Proc Nat Acad Sci USA 1986;83:7947-7951. 38. Blancou J, Kieny MP, Lathe R, Lecocq JP, Pastoret PP, Soulebot JP, Desmettre P. Oral vaccination of the fox against rabies using a live recombinant vaccinia virus. Nature 1986;322:373-376. 39. Esposito J, Brechling K, Baer G, Moss B. Vaccinia virus recombinants expresing rabies virus glycoprotein protect against rabies. Virus Gene 1987;1:7-21. 40. Tartaglia J, Perkus ME, Taylor J, Norton EK, Audonnet JC, Cox WI et al. NYVAC: a highly attenuated strain of vaccinia virus. Virology 1992;188:217- 232. 41. Wild TF, Bernard A, Spehner D, Drillien R. Construction of vaccinia virus recombinants expressing several measles virus protein and analysis of their efficacy in vaccination of mice. J Gen Virol 1992;73:359-367. 42. Taylor J, Pincus S, Tartaglia J, Richardson C, Alkhatib G, Briedis D et al. Vaccinia virus recombinants expressing either the measles virus fusion or hemagglutinin glycoprotein protect dogs against canine distemper virus challenge. J Virol 1991;65:4263-4274. 43. Drillien R, Spehner D, Kirn A, Giraudon P, Buckland R, Wild F, Lecocq JP. Protection of mice from fatal measles encephalitis by vaccination with vaccinia virus recombinants encoding either the hemagglutinin or the fusion protein. Proc Nat Acad Sci USA 1988; 85:1252-1256. 44. Bankamp G, Brinckmann UG, Reich A, Niewiesk S, terMeulen V, Liebert U. Measles virus nucleocapsid protein protects rats from encephalitis. J Virol 1991;65:1695-1700. 45. Moss B, Smith GL, Gerin JL, Purcell RH. Live recombinant vaccinia virus protects chimpanzees against hepatitis B. Nature 1984;311:67-69. 46. Paoletti E, Lipinska BR, Samsonoff C, Mercer S, Panicali D. Construction of live vaccines using genetically engineered poxviruses: biological activity of vaccinia virus recombinants expressing the hepatitis B virus surface antigen and the herpes simplex virus glycoprotein D. Proc Nat Acad Sci USA 1984;81:193-197. 47. Cremer KJ, Mackett M, Wohlenberg C, Notkins AL, Moss B. Vaccinia virus recombinant expressing herpes simplex virus type 1 glycoprotein D prevents latent herpes in mice. Science 1985;228:737-740. 48. Cantin EM, Eberle R, Baldick JL, Moss B, Willey DE, Notkins AL, Openshaw H. Expression of herpes simplex virus 1 glycoprotein B by a recombinant vaccinia virus and protection of mice against lethal herpes simplex virus 1 infection. Proc Nat Acad Sci USA 1987;84:5908-5912. 49. Wachsman M, Aurelian L, Smith CC, Lipinskas BR, Perkus ME, Paoletti E. Protection of Guinea pigs from primary and recurrent herpes simplex virus (HSV) type 2 cutaneous disease with vaccinia virus recombinants expressing HSV glycoprotein D. J Infect Dis 1987;155:1188-1197. 50. Jonjic S, DelVal M, Keil GM, Reddehase MJ, Koszinowski UH. A nonstructural viral protein expressed by a recombinant vaccinia virus protects against lethal cytomegalovirus infection. J Virol 1988;62:1653- 1658. 51. Reddehase MJ, Balthesen M, Papp M, Jonjic S, Pavic I, Koszinowski UH. The conditions of primary infection define the load of latent viral genome in organs and the risk of recurrent cytomegalovirus disease. J Exp Med 1994;179:185-193. 52. Smith GL, Murphy BR, Moss B. Construction and characterization of an infectious vaccinia virus recombinant that expresses the influenza hemagglutinin gene and induces resistance to influenza virus infection in hamster. Proc Nat Acad Sci USA 1983;80: 7155-7159.
53. Jakeman KJ, Smith H, Sweet C. Mechanisms of immunity to influenza: maternal and passive neonatal protection following immunization of adults ferrets with a live vaccinia-influenza virus hemagglutinin recombinant but not recombinants containig other influenza virus proteins. J Gen Virol 1989;70:1523-1531.
54. Rota PA, Shaw MW, Kendal AP. Cross-protection against microvariants of influenza virus type B by vaccinia virus expressing hemagglutinin from egg- or MDCK cell derived subpopulations of influenza virus type B/England/222/82. J Gen Virol 1989; 70: 1533-1537. 55. Rota PA, De BK, Shaw MW, Black RA, Gamble WC, Kendal AP. Comparison of inactivated live and recombinant DNA vaccines against influenza virus in mouse model. Virus Res 1990;16:83-94.
56. Sutter G, Wyatt LS, Foley PL, Bennink JR, Moss B. A recombinant vector derived from the host range restricted and highly attenuated MVA strain of vaccinia virus stimulates protective immunity in mice to influenza virus. Vaccine 1994;12:1032-1040. 57. Bray M, Zhao B, Markoff L, Eckels KH, Chanock RM, Lai C-J. Mice immunized with recombinant vaccinia virus expressing dengue 4 virus structural proteins with or without nonstructural protein NS1 are protected against fatal dengue 4 virus encephalitis. J Virol 1989;63:2853-2856. 58. Falgout B, Bray M, Schlesinger JJ, Lai CJ. Immunization of mice with recombinant vaccinia virus expressing authentic dengue virus nonstructural protein NS1 protects against lethal dengue virus encephalitis. Virology 1990;64:4356-4363. 59. Putnak JR, Schlesinger JJ. Protection of mice against yellow fever virus encephalitis immunization with a vaccinia virus recombinant encoding the yellow fever virus nonstructural proteins, NS1, NS2a, NS2b. J Gen Virol 1990;71:1697-1702. 60. Pincus S, Mason PW, Konishi E, Fonseca BAL, Shope RE, Paoletti E. Recombinant vaccinia virus producing the prM and E proteins of yellow fever virus protects mice from lethal yellow fever encephalitis. Virology 1992;187:290-297. 61. Yasuda A, Kimura-Kuroda J, Ogimoto M, Miyamoto M, Sata T, Sato T, Takamura C, Kurata C, Kojima A, Yasui K. Induction of protective immunity in animals vaccinated with recombinant vaccinia viruses that express preM and E glycoproteins of Japanese encephalitis virus. J Virol 1990;64:2788- 2795. 62. Konishi E, Pincus S, Fonseca BAL, Shope RE, Paoletti E, Mason PW. Comparison of protective immunity elicited by recombinant vaccinia viruses that synthesize E or NS1 of Japanese encephalitis virus. Virology 1991; 185:401-410. 63. Mason PW, Pincus S, Fournier MJ, Mason TL, Shope RE, Paoletti E. Japanese encephalitis virus vaccinia recombinants produce particulate forms of the structural membrane proteins and induce high levels of protection against lethal JEV infection. Virology 1991; 180:294-305. 64. Konishi E, Pincus S, Paoletti E, Laegreid WW, Shope RE, Mason PW. A highly attenuated host range-restricted vaccinia virus strain, NYVAC, encoding the prM, E, NS1 genes of japanese encephalitis virus prevents JEV viremia in swine. Virology 1992;190: 454-458. 65. Stott EJ, Taylor G, Ball LA, Anderson K, Young KKY, King AMQ, Wertz GW. Immune and histopathological responses in animals vaccinated with recombinant viruses that express individual genes of human respiratory syncytial virus. J Virol 1987;61:3855-3861. 66. Kulkarni AB, Connors M, Firestone CY, Morse III HC, Murphy BR. The cytolytic activity of pulmonary CD8+ lymphocytes induced by infection with a vaccinia virus recombinant expressing the M2 protein of respiratory syncytial virus(RSV) correlates with resistance to RSV infection in mice. J Virol 1993;67: 1044-1049. 67. Crowe Jr JE, Collins PL, London WT, Chanock RM, Murphy BR. A comparison in chimpanzees of the immunogenicity and efficacy of live attenuated RSV temperature-sensitive mutant vaccines and vaccinia virus recombinants that express the surface glycoproteins of RSV. Vaccine 1993;11:1395-1404. 68. Chen L, Thomas EK, Hu SL, Hellstrom I, Hellstrom KE. Human papillomavirus type 16 nucleoprotein E7 is a tumor rejection antigen. Proc Nat Acad Sci USA 1991;88:110-114. 69. Chen L, Mizuno MT, Singhal MC, Hu SL, Galloway DA, Hellstrom I, Hellstrom KE. Induction of cytotoxic T lymphocytes specific for syngeneic tumor expressing the E6 oncoprotein of HPV 16. Immunol 1992;148: 2617- 2621. 70. Meneguzzi G, Cerni C, Kieny MP, Lathe R. Immunization against HPV type 16 tumor cells with recombinant vaccinia viruses expressing E6 and E7. Virology 1991;181:62-69. 71. Spriggs MK, Collins PL, Tierny E, London WT, Murphy BR. Immunization with vaccinia virus recombinants that express the surface glycoproteins of human parainfluenza virus type 3 (PIV3) protects patas monkeys against PIV3 infection. J Virol 1988;62:1293-1296. 72. Auperin DD, Esposito JJ, Lange JV, Bauer SP, Knight J, Sasso DR, McCormick JB. Construction of a recombinant vaccinia virus expressing the Lassa virus glycoprotein gene and protection of Guinea pigs from a lethal Lassa virus infection. Virus Res 1988;9:233-248. 73. Clegg JCS, LLoyd G. Vaccinia recombinant expressing Lassa virus internal nucleocapsid protein protects Guinea pigs against Lassa fever. Lancet 1987;8552:186-189. 74. Barret T, Belsham GJ, Subbarao SM, Evans SA. Immunization with a vaccinia recombinant expressing the F protein protects rabbits from challenge with a lethal dose of rinderpest virus. Virology 1989;170:11- 18. 75. Yilma T, Hsu D, Jones L, Owens S, Grubman M, Mebus C, Yamanaka M, Dale B. Protection of cattle against rinderpest with vaccinia virus recombinant expressing the HA or F genes. Science 1988;242:1058-1061. 76. Giavedoni L, Jones L, Mebus C, Yilma T. A vaccinia virus double recombinant expressing the F and H genes of rinderpest virus protects cattle against rinderpest and causes no pock lesions. Proc Nat Acad Sci USA 1991;88:8011-8015. 77. Asano K, Tsukiyama K, Shibata S, Yamaguchi K, Momoki T, Maruyama T. Immunological and virological characterization of improved construction of recombinant vaccinia virus expressing rinderpest virus haemaglutinin. Arch Virol 1991;116:81-90. 78. Yamanouchi K, Inui K, Sugimoto M, Asano K, Nishimaki F, Kitching RP. Immunization of cattle with a recombinant vaccinia vector expressing the hemagglutinin gene of rinderpest virus. Vet Res 1993;132:152-156. 79. Guo P, Goebel S, Perkus ME, Taylor J, Norton E, Allen G et al. Coexpression by vaccinia virus recombinants of equine herpes virus-1 glycoproteins gp13 and gp14 results in potentiated immunity. J Virol 1990;64: 2399-2406. 80. Riviere M, Tartaglia J, Perkus ME, Norton EK, Molnar-Bongermino C, Desmettre P, Paoletti E. Immunological evaluation in mice and swine of pseudorabies virus glycoproteins gp50, gpII and gpIII expressed by vaccinia virus recombinants. J Virol 1992;66: 3424-3434. 81. Taylor J, Tartaglia J, Moran T, Webster RG, Bouquet JF, Quimby FW et al. The role of poxvirus vectors in influenza vaccine development. In: Proceedings of the Third International Symposium on Avian Influenza. University of Wisconsin-Madison Extension Duplicating Services, 1992:311- 335. 82. Portetelle D, Limbach K, Burny A, Mammerickx M, Desmettre P, Riviere M et al. Recombinant vaccinia virus expression of the bovine leukaemia virus envelope gene and protection of immunized sheep against infection. Vaccine 1991;9:194-200. 83. Gatei MH, Naif HM, Kumar S, Boyle DB, Daniel RCW, Good MF, Lavin MF. Protection of sheep against bovine leukemia virus (BLV) infection by vaccination with recombinant vaccinia viruses expressing BLV envelope glycoproteins: correlation of protection with CD4 T-cell response to gp51 peptide 51-70. J Virol 1993;67:1803-1810. 84. Ohishi K, Suzuki H, Yamamoto T, Maruyama T, Miki K, Ikawa Y et al. Protective immunity against BLV induced in carrier sheep by inoculation with a vaccinia virus-BLV env recombinant: association with cell-mediated immunity. J Gen Virol 1991;72: 1887-1892. 85. Meneguzzi G, Kieny MP, Lecocq JP, Chambon P, Cuzin F, Lathe R. Vaccinia recombinants expressing early bovine papilloma virus (BPV1) proteins: retardation of BPV1 tumour development. Vacine 1990; 8:199-204. 86. Lathe R, Kieny MP, Gerlinger P, Clertan P, Guizani I, Cuzin F, Chambon P. Tumour prevention and rejection with recombinant vaccinia. Nature 1987;326:878-880. 87. Miyazama M, Nishio J, Chesebro B. Protection against Friend retrovirus- induced leukemia by recombinant vaccinia viruses expressing the gag gene. J Virol 1992;66: 4497-4507. 88. Rumenapf T, Stark R, Meyers G, Thiel HJ. Structural proteins of hog cholera virus expressed by vaccinia virus: further characterization and induction of protective immunity. J Virol 1991;65:589-597. 89. Kinney RM, Esposito JJ, Mathews JH, Johson BJB, Roehrig JT, Barrett ADT, Trent DW. Recombinant vaccinia virus/Venezuelan equine encephalitis(VEE) virus protects mice from peripheral VEE virus challenge. J Virol 1988;62:4697-4702. 90. Monath TR, Cropp CB, Short WF, Bowen RA, Kinney RM, Roehrig JT, Trent DW. Recombinant vaccinia-Venezuelan equine encephalitis (VEE) vaccine protects non-human primates against parenteral and intranasal challenge with virulent VEE virus. Vaccine Res 1992;1:55-68. 91. Bowen RA, Short WA, Cropp CB, Mathews JH, Roehring JT, Kinney RM et al. Protection of horses immunized with recombinant vaccinia-Venezuelan equine encephalitis vaccine. Vaccine Res 1992;1:111-121. 92. Sakaguchi T, Takao S, Kiyotani K, Fujii Y, Nakayama T, Yoshida T. Expression of the HN, F, NP, and M proteins of Sendai virus by recombinant vaccinia viruses and their contribution to protective immunity against sendai virus infection in mice. J Gen Virol 1993; 74:479-484. 93. De BK, Shaw MW, Rota PA, Harmon MW, Esposito JJ, Rott R et al. Protection against virulent H5 avian influenza virus infection in chickens by an inactivated vaccine produced with recombinant vaccinia virus. Vaccine 1988;6:257-261. 94. Chambers TM, Kawaoka Y, Webster RG. Protection of chickens from lethal influenza infection by vaccinia-expressed hemagglutinin. Virology 1988;167:414-421. 95. Cox WI, Tartaglia J, Paoletti E. Poxvirus recombinants as live vaccines. In: Binns MM and Smith GL, editors. Recombinant Poxviruses, Boca Raton: CRC Press, 1992:123-162. 96. Klavinska LS, Whitton L, Joly E, Oldstone MBA. Vaccination and protection from a lethal viral infection: identification, incorporation, and use of a cytotoxic T lymphocyte glycoprotein epitope. J Virol 1990;178:393-400.
97. Connors M, Kulkarni AB, Collins PL, Firestone CY, Holms KL, Morse HC III, Murphy BR. Resistance to respiratory syncytial virus (RSV) challenge induced by infection with a vaccinia virus recombinant expressing the RSV M2 protein (Vac-M2) is mediated by CD8+ T cells while that induced by Vac-F or Vac-G recombinants is mediated by antibodies. J Virol 1992;66:1277- 1281. 98. Scheiflinger F, Falkner FG, Dorner F. Evaluation of the thymidin kinase (tk) locus as an insertion site in the highly attenuated vaccinia MVA strain. Arch Virol 1996;141: 663-669. 99. Antoine G, Scheiflinger F, Holzer G, Langmann T, Falkner FG, Dorner F. Characterization of the vaccinia MVA HA gene locus and its evaluation as an insertion site for foreign genes. Gene 1996;177:43-48. 100. Sutter G, Wyatt LS, Foley PL, Bennink JR, Moss B. A recombinant vector derived from the host range restricted and highly attenuated MVA strain of vaccinia virus stimulates protective immunity in mice to influenza virus. Vaccine 1994;12:1032-1040. 101. Sutter G, Moss B. Non-replicating vaccinia vector efficiently expresses recombinant genes. Proc Nat Acad Sci USA 1992;89: 10847-10851. 102. Shida H, Hinuma Y, Hatanaka M, Morita M, Kidokora M, Suzuki K et al. Effects and virulence of recombinant vaccinia viruses derived from attenuated strain that express the human T cell leukemia virus type 1 envelope gene. J Virol 1988;62:4474- 4480. 103. Flexner C, Hugin A, Moss B. Prevention of vaccinia virus infection in immunodeficient nude mice by vector-directed IL2 expression. Nature 1987;330:259-262. 104. Kohone-Corish MRJ, Long NJC, Woodhams CE, Ramshaw IA. Immunodeficient mice recover from infection with vaccinia virus expressing interferon gamma. Eur J Immunol 1990;172:1495-1503. 105. Buller RML, Smith GL, Cremer K, Notkins AL, Moss B. Decreased virulence of recombinant vaccinia virus expression vectors is associated with a thymidine kinase-negative phenotype. Nature 1985;317:813-815. 106. Buller RML, Chakrabarti S, Cooper JA, Twardzik DR, Moss B. Deletion of the vaccinia virus growth factor gene reduces virus virulence. J Virol 1988;67:866-874. 107. Child SJ, Palumbo G, Buller RM, Hruby D. Insertional inactivation of the large subunit of ribonucleotide reductase encoded by vaccinia virus is associated with reduced virulence in vivo. Virology 1990;174:625-629. 108. Rodriguez JR, Rodriguez D, Esteban M. Insertional inactivation of the vaccinia virus 32kD gene is associated with attenuation in mice and reduction of viral gene expression in polarized epithelial cells. J Virol 1992; 66:183-189. 109. Isaacs SN, Kotwal GJ, Moss B. Vaccinia virus complement-control protein prevents antibody-dependent complement-enhanced neutralization of infectivity and contributes to virulence. Proc Nat Acad Sci USA 1992;89: 628-632. 110. Lee SL, Roos JM, McGuigan LC, Smith KA, Cormier N, Cohen LK, Roberts BE, Payne LG. Molecular attenuation of vaccinia virus: mutant generation and animal characterization. J Virol 1992;66:2617-2630. 111. Tartaglia J, Cox WI, Taylor J, Perkus M, Riviere M, Meigner B, Paoletti E. Highly attenuated poxvirus vectors. AIDS Res Hum Retroviruses 1992;8:1445-1447. 112. US Federal Register. September 13, 1994;58 (175). 113. Brockmeier SL, Lager K, Tartaglia G, Riviere M, Paoletti E, Mengeling WL. Vaccination of pigs against pseudorabies with highly attenuated vaccinia (NYVAC) recombinant viruses. Vet Microb 1993;38:41-58. 114. Brockmeier SL, Mengeling WL. Comparison of the protective response by NYVAC vaccinia recombinants expressing either gp50 or gpII and gp50 of pseudorabies virus. Can J Vet Res 1996;60:315-317. 115. Pincus S, Tartaglia J, Paoletti E. Poxvirus-based vectors as vaccine candidates. Biologicals 1995;23:159-164. 116. Taylor J, Weinberg R, Languet B, Desmettre B, Paoletti E. Recombinant fowlpox virus inducing protective immunity in non-avian species. Vaccine 1988;6:497-503. 117. Fatunmbi OO, Reed WM. Evaluation of a commercial modified live virus fowlpox vaccine for the control of "variant" fowl poxvirus infections. Avian Dis 1996;40:582-587. 118. Taylor J, Weinberg R, Kawaoka Y, Webster R, Paoletti E. Protective immunity against avian influenza induced by a fowlpox virus recombinant. Vaccine 1988;6:504-508. 119. Webster RG, Kawaoka Y, Taylor J, Weinberg R, Paoletti E. Efficacy of nucleoprotein and hemagglutinin antigens expressed in fowlpox virus as vaccine for influenza in chickens. Vaccine 1991;9:303-308. 120. Ogawa R, Yanagida N, Saeki S, Saito S, Ohkawa S, Gotoh H et al. Recombinant fowlpox viruses inducing protective immunity against Newcastle disease and fowlpox viruses. Vaccine 1990;8:486-490. 121. Boursnell MEG, Green PF, Samson ACR, Campbell JIA, Deuter A, Peters RW et al. A recombinant fowlpox virus expressing the haemagglutinin- neuraminidase gene of Newcastle disease virus (NDV) protects chickens against challenge by NDV. Virology 1990;178:297-300. 122. Edbauer C, Weinberg R, Taylor J, Rey-Senelonge A, Bouquet JF, Desmettre P, Paoletti E. Protection of chickens with a recombinant fowlpox virus expressing the Newcastle disease virus hemagglutinin-neuraminidase gene. Virology 1990;179: 901-904. 123. Taylor J, Edbauer C, Rey-Senelonge A, Bouquet JF, Norton E, Goebel S et al. Newcastle disease virus fusion protein expressed in a fowlpox virus recombinant confers protection in chickens. J Virol 1990;64: 1441-1450. 124. Boursnell MEG, Green PF, Campbell JIA, Deuter A, Peters RW, Tomley FM et al. Insertion of the fusion gene from Newcastle disease virus into a non-essential region in the terminal repeats of fowlpox virus and demonstration of protective immunity induced by the recombinant. J Gen Virol 1990;71: 621-628. 125. Taylor J, Christensen L, Getting R, Goebel J, Bouquet JF, Mickle TR, Paoletti E. Efficacy of a recombinant fowlpox-based Newcastle disease virus vaccine candidate against velogenic and respiratory challenge. Avian Dis 1996;40:173-180. 126. Nazerian K, Lee LF, Yanagida N, Ogawa R. Protection against Marek s disease by a fowlpox virus recombinant expressing the glycoprotein B from Marek's disease Virus. J Virol 1992;66:1409-1413. 127. Nazerian K, Yanagida N. A recombinant fowlpox virus expressing the envelope antigen of subgroup A avian leukosis/sarcoma virus. Avian Dis 1995;39:514-520. 128. Paoletti E, Tartaglia J, Cox WI, inventors. Virogenetics corporation, assignee. Immunodeficiency virus recombinant poxvirus vaccine. WO9222641. 1992 Dec 23. 129. Radaelli A, Gimelli M, Cremonesi C, Scarpini C, deGiuliMorghe C. Humoral and cell mediated immunity in rabbits immunized with live non- replicating avipox expressing the HIV-1 SF2 env gene. Vaccine 1994;12: 1110-1117. 130. Taylor J, Trimarchi C, Weinberg R, Languet B, Guillemin F, Desmettre P, Paoletti E. Efficacy studies on a canarypox-rabies recombinant virus. Vaccine 1991;9:190-193. 131. Taylor J, Tartaglia J, Riviere Y, Duret C, Languet B, Chappui G, Paoletti E. Application of canarypox (ALVAC) vectors in human and veterinary vaccination. In: Brown F, editor. Recombinant vectors in vaccine development. Dev Biol Stand Basel: Karger, 1995:165-170. 132. Taylor J, Weinberg R, Tartaglia J, Richardson C, Alkhatib G, Briedis D et al. Non-replicating viral vectors as potential vaccines: recombinant canarypox virus expressing measles virus fusion (F) and hemagglutinin (HA) glycoproteins. Virology 1992;187:321-328. 133. Stephensen CB, Welter J, Thaker SR, Taylor J, Paoletti E. CDV infection of ferrets as a model for testing morvillivirus vaccine strategies: NYVAC and ALVAC based CDV recombinants protects against symptomatic infection. J Virol 1997;71:1506-1513. 134. Tartaglia G, Jarrett O, Neil JC, Desmettre P, Paoletti E. Protection of cats against feline leukemia virus by vaccination with a canarypox virus recombinant, ALVAC-FL. J Virol 1993;67:2370-2375. 135. Konishi E, Pincus S, Paoletti E, Shope RE, Mason PW. Avipox virus- vectored japanese encephalitis virus vaccine: use as vaccine candidate in combination with purified subunit immunogens. Vaccine 1994;12:633-638. 136. Gonczol E, Berencsi K, Pincus S, Endresz V, Meric C, Paoletti E, Plotkin S. Preclinical evaluation of an ALVAC(canarypox)- human cytomegalovirus glycoprotein B vaccine candidate. Vaccine 1995;13:1080- 1085. 137. Cadoz M, Strady A, Meigner B, Taylor J, Tartaglia J, Paoletti E, Plotkin S. Immunization with canarypox virus expressing rabies glycoproteins. Lancet 1992;339:1429-1432. 138. Taylor J, Meigner B, Tartaglia J, Languet B, Vanderhoeven J, Franchini G et al. Biological and immunological properties of a canarypox-rabies recombinant, ALVAC RG (vCP65) in non-avian species. Vaccine 1995; 13:539- 549. 139. Pialoux G, Excler JL, Riviere Y, Gonzalez-Canalis G, Feuillie V, Coulaud P et al. A prime-boost approach to HIV preventive vaccine using a recombinant canarypox virus expressing glycoprotein 160 (MN) followed by a recombinant glycoprotein 160 (MN/LAI). AIDS Res Hum Retroviruses 1995;11:373-381. 140. Clements ML, Weinhold KJ, Siliciano RF, Schwartz D, Mattews T, Graham B et al. HIV immunity induced by canarypox (ALVAC)-gp160/MN, rgp120/SF2 or both [Abstract]. XI Internatinal Conference on AIDS, Vancouver. 1996;Mo. A 281. 141. Lawrence C, Weinhold KJ, McElrath J, Excler JL, Duliege AM, Clements ML et al. Safety and immunogenicity of live recombinant canarypox virus vector containing the envelope, gag, and the protease genes of HIV-1 in seronegatives adult volunteers [Abstract]. XI International Conference on AIDS, Vancouver. 1996;Mo. A282. 142. Egan MA, Pavlat WA, Tartaglia J, Paoletti E, Weinhold KJ, Clements ML, Siliciano RF. Induction of HIV-1 specific cytolytic T lymphocyte responses in seronegative adults by a replicating, host-range-restricted canarypox (ALVAC) carrying the HIV-1/MN env gene. J Infect Dis 1995;171:1623-1627. 143. Lanar DE, Tine JA, deTaisne C, Seguin MC, Cox WI, Winslow JP et al. Attenuated vaccinia virus- circumsporozoite protein recombinants confer protection against rodent malaria. Infect Immun 1996;64:1666-1671. 144. Tine JA, Lanar DE, Smith DM, Wellde BT, Schultheiss P, Ware LA et al. NYVAC-Pf7: a poxvirus-vectored, multiantigen, multistage vaccine candidate for Plasmodium falciparum malaria. Infection and Immunity 1996;64: 3833- 3844. 145. Fast P. HIV vaccine clinical trials: experience of the NIAID AIDS vaccine evaluation group. In: Girard M and Dodet B, editors. X Colloque des cent gardes. Retroviruses of human AIDS and related animal diseases, Francia, 1995:283-290. 146. Hu SL, Klaniecki J, Dykers T, Sridhar P, Travis BM. Neutralizing antibodies against HIV-1 BRU and SF2 isolates generated in mice immunized with recombinant vaccinia virus expressing HIV-1(BRU) envelope glycoproteins and boosted with homologous gp160. AIDS Res Hum Retroviruses 1991;7:615-620. 147. Dallo S, Maa JS, Rodriguez JR, Rodriguez D, Esteban M. Humoral immune response elicited by highly attenuated variants of vaccinia virus and by an attenuated recombinant expressing HIV-1 envelope protein. Virology 1989;173:323-329. 148. Hu SL, Zarling JM, Chinn J, Travis BM, Moran PA, Sias J et al. Protection of macaques against simian AIDS by immunization with a recombinant vaccinia virus expressing the envelope glycoprotein of simian type D retrovirus.Proc Nat Acad Sci USA 1989; 86:7213-7217. 149. Hu SL, Abrams K, Barber GN, Moran P, Zarling JM, Langlois AJ et al. Protection of macaques against SIV infection by subunit vaccines of SIV envelope glycoprotein gp160. Science 1992;255:456-459. 150. Vogt G, le Grand R, Vaslin B, Boussin F, Bayon-Auboyer MH, Rivier Y et al. Heterologous HIV-2 challenge of rhesus monkeys immunized with recombinant vaccinia viruses and purified recombinant HIV-2 proteins. Vaccine 1995;13:202-208. 151. Dru A, Ludosky MA, Cataud J, Beaud G. Cell-line dependent release of HIV-like gag particles after infection of mammalian cells with recombinant vaccinia virus. AIDS Res Hum Retrov 1994;10:383-390. 152. Cooney EL, McElrath MJ, Corey L, Hu SL, Collier AC, Arditti D et al. Enhanced immunity to human immunodeficiency virus (HIV) envelope elicited by a combined vaccine regimen consisting of priming with a vaccinia recombinant expressing HIV envelope and boosting with gp160 protein. Proc Nat Acad Sci USA 1993;90:1882-1886. 153. Graham BS, Matthews TJ, Belshe RB, Clements ML, Dolin R, Wright PF et al. Augmentation of HIV-1 neutralizing antibody by priming with gp160 recombinant vaccinia virus and boosting with rgp160 in vaccinia-naive adults. J Infect Dis 1993;167:533-537. 154. Cooney EL, Collier AC, Greenberg PD, Coombs RW, Jorling J, Arditti DE et al. Safety and immunological response to a recombinant vaccinia virus vaccine expressing HIV envelope glycoprotein. Lancet 1991;337: 567-572. 155. Abimiku A, Franchini G, Tartaglia J, Aldrich K, Myagkikh M, Markham PD et al. HIV-1 recombinant poxvirus against HIV-2 challenge in rhesus macaques. Nature Med 1995;1:321-329. 156. Franchini G, Robert-Guroff M, Tartaglia J, Aggarwal A, Abimiku A, Benson J et al. Highly attenuated HIV-2 recombinant poxviruses, but not HIV-2 recombinant Salmonella vaccines, induce long-lasting protection in rhesus macaques. AIDS Res Hum Retroviruses 1995;11:909-919. 157. Biberfield G, Thorstensson R, Putkonen P. Protection against HIV-2 and SIV in macaques vaccinated against HIV-2. AIDS Res Hum Retroviruses 1996;12:443-446. 158. Myagkikh M, Alipanah S, Markham PD, Tartaglia J, Paoletti E, Gallo RC et al. Multiple immunizations with attenuated poxvirus HIV type 2 recombinants and subunit boosts required for protection of rhesus macaques. AIDS Res Hum Retroviruses 1996;12:985-992.
159. Girard M. Homologous and heterologous protection from HIV-1 infection in chimpanzees. In: Girard M and Dodet B, editors. X Colloque des cent gardes. Retroviruses of human AIDS and related animal diseases, Francia, 1995:179-183. 160. Girard M, Meigner B, Barre-Sinuossi F. Vaccine protection of chimpanzees against infection by heterologous HIV-1. J Virol 1995;69:6239- 6248. Copyright 1998 Elfos Scientiae The following images related to this document are available:Line drawing images[ba98001a.gif] |
| |||||||||

{kind=link}