 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
CARACTERIZACION DEL FONDO GENETICO DE LA LINEA DE SUPERTILAPIAS IG/03- F70. I. CARACTERES MORFOMETRICOS, MERESTICOS Y ANALISIS DE ADN Reynold Morales,^1 Amilcar Arenal,^2 Rafael Pimentel,^2 Irian Mendoza,^2 Asterio Cruz,^3 Rebeca Martinez,^1 Fidel Herrera,^1 Leopoldo Tapanes,^4 Mario P Estrada^1 y Jose de la Fuente^1
^1 Division de Genetica de Celulas de Mamiferos.
Recibido en marzo de 1997. Aprobado en julio de 1997.
Code Number: BA98002
ABSTRACT Morphological, meristic and DNA differences have been established in the characterization of various strains of tilapia (Oreochromis spp). The present study shows the characterization of the transgenic line of supertilapia IG/03-F70. Morphological and meristic data show a high correlation with the tilapia Oreochromis hornorum. The chromosomal number and DNA satellite analysis support the hypothesis that the parents of supertilapias IG/03-F70 progenitors are hybrids of an unknown origen with a high percentage of O. hornorum. Key words: tilapia, biometry, chromosome, DNA satellite RESUMEN En la caracterizacion de varias especies de tilapias Oreochromis spp han sido establecidas sus caracteristicas morfologicas, meristicas y a nivel de ADN. En el presente estudio se muestra la caracterizacion de la linea supertilapias IG/03-F70. Los datos morfologicos y meristicos obtenidos de la linea estudiada muestran una gran similitud con la especie Oreochromis hornorum. Los resultados del conteo de cromosomas y el analisis del ADN satelite de la linea IG/03-F70, apoyan la hipotesis de que los progenitores de esta linea son hibridos de origen desconocido con un alto por ciento de O. hornorum. Palabras claves: tilapia, biometria, cromosoma, ADN satelite Introduccion La tilapia pertenece a la familia de los ciclidos y constituye un grupo amplio de peces endemicos de frica, donde su origen se remonta a varios millones de anos (1). El interes por la ecologia y la sistematica de este grupo de peces data de principios de este siglo y con el tiempo se ha incrementado su estudio por la importancia que tiene esta especie para la acuicultura en zonas tropicales (2). Basandose en los habitos reproductivos y alimenticios de estas especies, se han clasificado en dos generos: Tilapia y Sarotherodon (3). En 1982, basado en los estudios sobre sus habitos reproductivos y en la forma de incubar los huevos, se decide separar a las tilapias en 4 generos: Tilapia, Sarotherodon, Oreochromis y Danakilia (4). En 1983, Trewavas y colaboradores separan a las tilapias en 6 generos, tomando en consideracion la denticion, asi como sus habitos reproductivos: Tilapia, Tristamella, Danakilia, Sarotherodon, Oreochromis y Pelmatochromis (5), este ultimo es un ciclido menos especializado (6). Entre los metodos utilizados para el estudio de individuos y poblaciones se encuentran el estudio de los caracteres morfometricos y meristicos (1), los patrones isoenzimaticos (7, 8), asi como el numero y morfologia de los cromosomas (2). La labor taxonomica ha servido para corroborar estudios preliminares en la caracterizacion y diferenciacion de especies de tilapia, asi como para registrar nuevas especies endemicas (6). Las tecnicas de cariotipaje se han aplicado ampliamente en peces para determinar el sexo genetico (9, 10), poliploidia (11-13), relacion evolutiva entre especies cercanas (14), asi como la ubicacion en el genoma de una informacion genetica determinada (10). Existen otros metodos que se apoyan en el estudio del ADN, no solo referente a su composicion, sino tambien a su organizacion (15). Teniendo en cuenta que en cada individuo el ADN es diferente en estos dos aspectos, hay rasgos comunes entre individuos de una misma especie y familia que los identifican como miembros de estas (16, 17). El estudio de las huellas digitales del ADN ha probado ser una herramienta extremadamente poderosa en la identificacion de individuos y pedigrees (18) en el estudio de lineas ginogeneticas y de la transmision paterna de algunos fragmentos de ADN durante la generacion de diploides ginogeneticos (19). Otros laboratorios han utilizado determinados marcadores de ADN nuclear en estudios filogeneticos, identificacion de cepas hibridas y para el manejo acuicola de estos peces (17). Con este trabajo nos propusimos hacer una caracterizacion de la linea de supertilapias IG/03-F70. Esta linea se formo a partir de tilapias transgenicas que portan una copia adicional del gen que codifica la hormona de crecimiento de tilapia, lo cual se mostro fenotipicamente con un crecimiento acelerado (20-22). A partir del estudio de los caracteres meristicos, morfometricos, preparaciones cromosomicas y el analisis de ADN satelite de animales de varias generaciones de peces de esta linea transgenica, se establecio con que especie de tilapia presenta mayor similitud, y se conocio que sus progenitores son de origen hibrido desconocido. Los resultados indicaron que el hibrido utilizado para la obtencion de tilapias transgenicas a partir de las cuales se genero la linea de supertilapias IG/03-F70, es mayoritariamente Oreochromis hornorum. Materiales y Metodos Caracteres morfometricos y meristicos Este trabajo se realizo con larvas y juveniles de la linea de supertilapias IG/03-F70, capturadas al azar en la estacion experimental de Guanimar, provincia La Habana, en las cuales no se considero el sexo. Los individuos se agruparon segun su peso corporal, y se constituyeron 4 grupos de animales (Tabla 1). Tabla 1. Variabilidad de las caracteristicas estudiadas por grupo de peces.
---------------------------------------------------------------------------
Caracteristicas Coeficientes de variacion (%)^a Variabilidad^b
---------------------------------------------
Grupo 1c Grupo 2c Grupo 3c Grupo 4c Media
---------------------------------------------------------------------------
Peso (g) 32,89 16,41 12,82 11,85 18,49 III
Longitud total (mm) 9,31 5,37 5,55 4,41 6,16 II
Longitud estandar (mm) 10,78 5,73 5,18 3,71 6,35 II
Longitud cabeza (mm) 11,77 15,11 7,49 10,91 11,32 III
Longitud preorbital(mm) 9,98 9,13 12,23 20,00 12,84 III
Longitud predorsal (mm) 8,86 6,34 9,22 8,64 8,27 II
Longitud preventral(mm) 10,67 4,87 6,44 7,37 7,34 II
Longitud preanal (mm) 11,75 6,10 3,85 3,55 6,31 II
Base aleta dorsal (mm) 11,95 8,32 7,74 2,04 7,51 II
Base aleta anal (mm) 16,87 13,13 15,92 8,76 13,67 III
Longitud aleta pectoral 18,44 9,78 2,80 7,88 9,72 II
(mm)
Altura maxima del 10,01 5,58 4,66 6,04 6,57 II
cuerpo (mm)
Rayos aleta dorsal 3,11 2,53 5,18 4,35 3,79 I
Radios aleta dorsal 7,59 4,08 11,19 3,97 6,71 II
Radios aleta caudal 5,02 0,00 5,00 5,58 3,90 I
Rayos aleta anal 0,00 0,00 0,00 0,00 0,00 I
Radios aleta anal 7,00 0,00 5,38 0,00 3,09 I
Rayos aleta ventral 0,00 0,00 0,00 0,00 0,00 I
Radios aleta ventral 0,00 0,00 0,00 0,00 0,00 I
Radios aleta pectoral 3,42 3,87 3,87 3,42 3,64 I
Escamas en una serie 3,32 3,37 2,38 0,00 2,27 I
longitudinal
Branquispinas en el 4,32 4,56 4,55 0,00 3,36 I
primer arco branquial
Longitud del hueso 5,09 2,08 7,20 14,82 7,30 II
faringeo inferior (mm)
Numero de vertebras 2,54 2,75 3,03 3,32 2,91 I
---------------------------------------------------------------------------
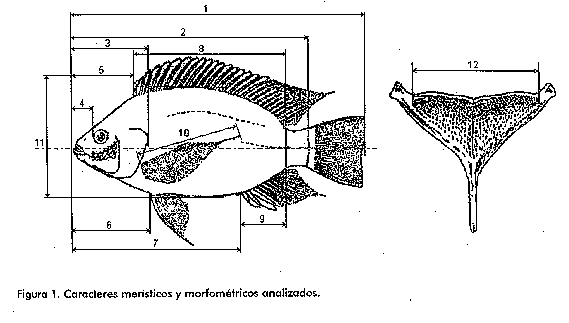
^a: Desviacion estandar/media x 100; ^b: los caracteres biometricos se clasificaron en 3 grupos atendiendo al coeficiente de variacion. 1) de 1 a 5: poco variable, 2) entre 5 y 10: variable y 3) mayor de 10: muy variable; ^c: los individuos se agruparon segun su peso corporal. Grupo 1: entre 1 y 5 g, N = 7; Grupo 2: entre 5,1 y 20,9 g, N = 6; Grupo 3: entre 21 y 30,9 g, N = 8 y Grupo 4: mayores de 31 g, N = 7. Para realizar las mediciones y los conteos se utilizo una balanza, un ictiometro y, en algunos casos, fue necesario emplear un microscopio estereoscopico con ocular micrometrico. Caracteristicas meristicas analizadas Numero de escamas en una serie longitudinal. Son las que se pueden contar desde el borde posterior del operculo hasta el pliegue vertical de articulacion en la base de la aleta caudal. Conteo de rayos y radios de las aletas. El tronco y la cola presentan las aletas pares (pectorales y ventrales) e impares (dorsal, anal y caudal). Las aletas pueden estar formadas por espinas, elementos de sosten rigidos y puntiagudos, no ramificados y por radios, elementos de sosten tambien, pero de consistencia blanda y ramificados, o por ambos a la vez. Numero de branquispinas en el primer arco branquial inferior. Se realiza en el primer arco branquial y, en nuestro caso, en el limbo inferior solamente. La union de los limbos se puede hallar en un punto de division, y si una branquispina se encuentra entre ambos angulos del arco, esta se incluye generalmente en el conteo del limbo inferior, asi como todas las branquispinas rudimentarias en el final anterior. Numero de vertebras. Para realizar el conteo fue necesario sacrificar el animal, hervirlo y proceder a la diseccion de sus tejidos; de este modo quedaron expuestas las vertebras. Medidas morfometricas analizadas (Figura 1)
. Peso en gramos (la pesada se realizo con los animales vivos)
Analisis estadistico El procesamiento estadistico se realizo en hojas de trabajo de Excel (Microsoft Excel for Windows 95, version 7.0). Antes del procesamiento estadistico de los datos se comprobo la homogeneidad de varianza y la normalidad. Las relaciones entre los parametros en los grupos de peces fueron contrastadas mediante un analisis de varianza de una via y se realizo la prueba de Duncan para los casos en que dio significacion. Obtencion de tejidos en division celular Para realizar este trabajo se emplearon tilapias adultas de la linea de supertilapias IG/03-F70, a las que se les realizo un corte transversal en la parte posterior de la aleta caudal. Al cabo de los 3-7 dias la porcion regenerada del tejido de la aleta, de aproximadamente 2 mm, fue utilizada para la preparacion cromosomica. Preparacion de metafases y conteo de cromosomas Se realizaron modificaciones a la tecnica descrita por Chourrout e Itskovich (11) para larvas de tilapias. Una vez cortado el fragmento de aleta regenerado, se coloco en una placa Petri de 60 mm de diametro, a la cual se le anadio colchicina (SIGMA) al 0, 002 %, y se incubaron a 28 C entre 4 a 6 h. Posteriormente, se le dio un shock hipotonico con agua destilada durante 90 min. Despues se fijo con metanol - acido acetico (3:1). Se realizaron 4 fijaciones, cada una de 20 min. Una vez eliminado el exceso de fijador, se coloco la muestra en un portaobjetos excavado, se le adicionaron 2 o 3 gotas de acido acetico al 60 %, en presencia del cual se trituro el tejido por 1 min, y se dejo reposar 20 min. Transcurrido ese tiempo, se dejaron caer 2 gotas de este preparado con una pipeta Pasteur sobre un portaobjetos, previamente conservado en metanol al 50 % y secado a 50 C. Ambas gotas se extendieron con la parte fina de la pipeta, el portaobjetos se puso sobre una plancha caliente durante un periodo corto de tiempo hasta que se evaporo el acido acetico. Antes de tenir la lamina, se dejo secar al aire durante 2 dias. La tincion se realizo con Giemsa (SIGMA) al 10 % en tampon fosfato 10 mM, pH 7, durante 8 min. Se lavo con agua destilada y se dejo secar al aire. Una vez secas las laminas, se observaron al microscopio optico a un aumento de 100 X y los detalles se observaron a un aumento de 1 000 X con lente de inmersion en aceite. Durante la observacion se presto atencion al numero de cromosomas de cada metafase, asi como a su morfologia. Solo se tuvieron en cuenta aquellas metafases en las cuales sus cromosomas se pudieron observar con nitidez. Se realizo el conteo de los cromosomas de 46 metafases. Obtencion y analisis del ADN satelite Se seleccionaron 5 tilapias al azar dentro de una poblacion de la linea de supertilapias IG/03-F70, sin tener en cuenta el sexo. Como fuente de ADN se tomo un fragmento con un peso de 30 mg de la aleta caudal, el cual fue digerido con Proteinasa K (100 g/mL; Boehringer Mannheim) en tampon de lisis (Tris-HCl, 10 mM; SDS, 0, 5 %; EDTA, 100 mM, pH 7, 5) durante 5 h a 55 C. La muestra se trato sucesivamente con fenol, fenol: cloroformo y cloroformo. La fase acuosa se incubo 1 h a 55 C para eliminar los restos de cloroformo; posteriormente se incubo 1 h a 37 C con Ribonucleasa A (100 g/mL; Boehringer Mannheim). Se aplicaron extracciones sucesivas de fenol: cloroformo y cloroformo, respectivamente. Finalmente, el ADN fue precipitado con 2 volumenes de etanol en presencia de acetato de sodio 300 mM, lavado con etanol 70 % y resuspendido en 200 L de agua destilada esteril. Se tomaron 10 g de ADN genomico de cada pez y la digestion se realizo con la enzima de restriccion EcoR I (2 unidades/ g de ADN; New England Biolabs) por 3 h a 37 C. El ADN digerido con EcoR I se aplico en un gel de agarosa de bajo punto de gelificacion (LGT, SIGMA) al 0, 9 %, utilizando un marcador de peso molecular (pGEM^ DNA Markers; Promega), que consiste en 15 fragmentos de ADN en un rango entre 36 y 2 645 pb. Se hizo una extraccion fenolica de la zona del gel entre 200 y 350 pb y las bandas extraidas se resuspendieron en 10 L de agua. De estas bandas, se tomaron 3 L y se separaron a 10 v/cm durante 8 h en un gel de poliacrilamida al 12 % en condiciones no desnaturalizantes. Una vez terminada la corrida, se tomo el gel adherido a uno de los cristales y se trato con acido acetico al 12 % durante 30 min, luego, 3 veces en agua desionizada durante 2 min, 30 min en solucion de tincion (nitrato de plata 0,1 %; formaldehido 0,05 %) y 10 s en agua desionizada. Para el revelado, se mantuvo en solucion de formaldehido 0,05 %; tiosulfato de sodio 2 g/mL y carbonato de sodio 3 % hasta que aparecieron las bandas. Por ultimo, se fijo en acido acetico 10 % y se seco en un horno de microondas a 250 C por 7 min. La talla de las bandas que se observaron en el gel se estimo mediante la interpolacion de los valores de la distancia de migracion de las bandas en cuestion en una curva obtenida ploteando el logaritmo decimal del peso molecular contra la distancia de migracion de las bandas del patron, tomando como 0 el punto de aplicacion. Resultados y Discusion Caracteres morfometricos y meristicos
De acuerdo con el coeficiente de variacion, las medidas muy variables fueron el peso, longitud de la cabeza, longitud preorbital y la base de la aleta anal (Tabla 1). Los parametros variables fueron la longitud total, longitud estandar, longitud predorsal, longitud preventral, longitud preanal, base de la aleta dorsal, longitud de la aleta pectoral, altura maxima del cuerpo, radios de aleta dorsal y longitud del hueso faringeo inferior (Tabla 1). Por ultimo, fueron poco variables los radios de la aleta caudal, los rayos de la aleta dorsal, rayos y radios de la aleta anal, rayos y radios de la aleta ventral, radios de la aleta pectoral, escamas en una serie longitudinal, branquispinas en el primer arco branquial, asi como el numero de vertebras (Tabla 1). Resultados semejantes fueron obtenidos por Gourene y Teugels (1). De las relaciones del cuerpo analizadas, las de longitud total/longitud estandar, longitud preanal/longitud estandar, longitud preventral/longitud estandar, base de la aleta dorsal/longitud estandar y la altura maxima del cuerpo/longitud estandar fueron poco variables (Tabla 2). Tabla 2. Variabilidad de las relaciones de parametros por grupo de peces.
---------------------------------------------------------------------------
Relacion de parametros Coeficientes de variacion (%)^a Variabilidad^b
--------------------------------------
Grupo Grupo Grupo Grupo Media
1c 2c 3c 4c
Longitud total/longitud estandar 2,14 1,91 0,97 1,57 1,65 I
Longitud cabeza/longitud estandar 3,29 11,97 7,49 8,29 7,76 II
Longitud preorbital/
longitud estandar 3,52 7,79 8,84 17,25 9,35 II
Longitud preorbital/longitud cabeza4,38 15,34 11,93 9,42 10,26 III
Longitud predorsal/longitud
estandar 12,74 1,73 5,17 6,56 6,55 II
Longitud preanal/longitud estandar 2,80 1,62 2,58 2,71 2,43 I
Longitud preventral/longitud
estandar 3,25 4,00 5,90 4,95 4,53 I
Base de la aleta dorsal/longitud
estandar 2,57 8,11 3,77 2,93 4,34 I
Base de la aleta anal/longitud
estandar 9,07 15,15 13,59 7,04 11,21 III
Longitud de la aleta pectoral/
longitud estandar 8,75 8,01 3,65 6,23 6,66 II
Altura maxima del cuerpo/longitud
estandar 7,01 2,41 3,10 4,99 4,38 I
Longitud del hueso faringeo/
longitud cuerpo 7,43 14,54 8,43 4,95 8,84 II
--------------------------------------------------------------------------
^a: Desviacion estandar/media x 100; ^b: los caracteres biometricos se clasificaron en 3 grupos atendiendo al coeficiente de variacion. 1) de 1 a 5: poco variable, 2) entre 5 y 10: variable y 3) mayor de 10: muy variable; ^c: los individuos se agruparon segun su peso corporal. Grupo 1: entre 1 y 5 g, N = 7; Grupo 2: entre 5,1 y 20,9 g, N = 6; Grupo 3: entre 21 y 30,9 g, N = 8 y Grupo 4: mayores de 31 g, N = 7. Se catalogaron como variables, las de longitud de la cabeza/longitud estandar, longitud preorbital/longitud estandar, longitud predorsal/longitud estandar, longitud de la aleta pectoral/longitud estandar y longitud del hueso faringeo/longitud de la cabeza (Tabla 2). Se consideraron parametros muy variables la longitud preorbital/longitud de la cabeza y la base de aleta anal/longitud estandar (Tabla 2). El analisis de varianza entre los 4 grupos de tilapias estudiados, respecto a las relaciones de parametros (Tabla 2), muestra que varian significativamente las relaciones longitud total, longitud preorbital, longitud preanal y base de la aleta dorsal, referidas a la longitud estandar. Al contrastarlas con la prueba de Duncan, se observa que existen diferencias significativas entre algunos grupos (Tabla 3). Tabla 3. Comparacion multiple de las medias de los grupos de peces utilizando la prueba de Duncan.
---------------------------------------------------------------------------
Grupo Longitud total/ Longitud Longitud preanal/ Base aleta dorsal/
longitud estandar preorbital/ longitud estandar longitud estandar
longitud estandar
---------------------------------------------------------------------------
Media S Media S Media S Media S
---------------------------------------------------------------------------
1 127,11 * 12,21 * * 72,06 * 54,51 *
2 126,03 * 11,07 * 71,91 * 57,95 * *
3 25,04 * 11,26 * 73,44 * 60,40 *
4 76,42 * 13,25 * 68,35 * 57,12 * *
---------------------------------------------------------------------------
S: Una posicion distinta de los asteriscos denota diferencia significativa
entre dos promedios. Cuando aparecen dos asteriscos en una media se debe a
que la misma puede ser igual a sus medias adyacentes, pero estas ultimas
entre si son diferentes.El conteo de los rayos y radios de las aleta dorsal, anal y ventral en la linea de supertilapias IG/03-F70 permitio calcular la relacion rayo/radio de esta linea (Tabla 4), parametro que juega un papel importante al comparar con las claves para identificar especies de tilapias publicadas por otros autores (6). Tabla 4. Relacion en por ciento de rayos/radios en las aletas dorsal, anal y ventral en la linea de supertilapias IG/03-F70.
------------------------------
Media^1 DS^2
------------------------------
Aleta dorsal 56,7 2,36
Aleta anal 23,4 0,84
Aleta ventral 16,7 0,00
------------------------------
^1: Media (N = 28) de la relacion rayos/radios en las aletas dorsal, anal y
ventral de la linea de supertilapias IG/03-F70; ^2: desviacion estandar. Relacion rayos/radios = (numero rayos/(numero rayos + numero de radios)) x 100. En la Tabla 5 se muestra la semejanza de la linea de supertilapias IG/03-F70, en porcentajes, con otras especies de tilapias, respecto a varios parametros estudiados, comparados con los rangos publicados en las claves de Trewavas para identificar especies de tilapias (6). Analizando el porcentaje de semejanza global, se puede concluir que la linea de supertilapias IG/03-F70 es un hibrido con un alto contenido de O. hornorum. Tabla 5. Coincidencia en por ciento de los parametros estudiados entre la linea de supertilapias IG/03-F70 y otras especies de tilapias del genero Oreochromis.
---------------------------------------------------------------------------
Parametros de la Coincidencia Coincidencia Coincidencia Coincidencia
linea de supertilapias con O. con O. con O. con O.
IG/03-F70 mossambicus^a aureus^a niloticus^a hornorum^a
---------------------------------------------------------------------------
Rayos aleta dorsal 100 21 79 100
Radios aleta dorsal 57 100 89 100
Rayos aleta anal 100 100 100 100
Radios aleta anal 96 82 100
Escamas en una serie
longitudinal 89 89 7 NR
Branquispinas en el primer
arco branquial NR 50 50 100
Por ciento de semejanza global 74 72 68 100
--------------------------------------------------------------------------
^a: Por ciento del total de los animales estudiados (4 grupos, N = 28,
Tabla 1) de la linea de supertilapias IG/03-F70 que coinciden con el rango,
para cada especie, publicado por Trewavas en las claves para identificacion
de especies de tilapias (6). NR: No reportado.Numero de cromosomas de la linea estudiada En la linea de supertilapias IG/03-F70 se estudio el numero de cromosomas por celula en 3 generaciones distintas (F1, F2 y F3), y se obtuvo una distribucion estrecha de la frecuencia de cromosomas por celula, con un pico bien definido en 44, lo que se corresponde con el numero de cromosomas reportado por otros autores para el genero Oreochromis (12, 13) (Tabla 6). En algunas celulas se observo hipoploidia y en ningun caso hiperploidia; ello demostro que la tecnica utilizada provoca la perdida de algunos cromosomas, la cual es la causa de la aparicion de celulas con menos de 44 cromosomas como han descrito otros autores (12, 13) (Tabla 6). Tabla 6. Numero de cromosomas por celula en la linea de supertilapias IG/03-F70. --------------------------------------------------------------------------- Numero del Numero total de Numero de metafases Numero de metafases con animal metafases con 44 cromosomas menos de 44 cromosomas (generacion) analizadas (frecuencia %) (frecuencia %) --------------------------------------------------------------------------- 28 (F1) 46 36 (78,3) 10 (21,7) 43 (F2) 24 22 (91,6) 2 (8,4) 5 (F3) 16 14 (87,5) 2 (12,5) --------------------------------------------------------------------------- Morfologia de los cromosomas Mediante esta tecnica fue posible observar que 2 de los 44 cromosomas fueron de 4 a 5 veces mayores que el resto (Figura 2). Los restantes son poco diferenciables entre si, en tamano y forma (Figura 2). No se observaron cromosomas metacentricos, siendo la mayoria de ellos acrocentricos y subtelocentricos, ni se identificaron cromosomas sexuales (Figura 2).
Al digerir el ADN de tilapias de la linea de supertilapias IG/03-F70 con EcoR I y aplicarlo en una electroforesis en agarosa 0,9 % (LGT, SIGMA), se observo una banda difusa (resultado no mostrado) en una zona cuya talla fue superior a los 222 pb e inferior a los 350 pb. Una vez purificadas las bandas de cada una de las muestras procesadas y efectuada la tincion con plata en un gel de poliacrilamida al 12 %, se pudo observar la presencia de 2 bandas en la zona mencionada (Figura 3). Se determino la ecuacion de la curva que expresa el logaritmo del peso molecular en funcion de la distancia de migracion de los fragmentos de ADN, a partir de un patron de peso molecular conocido (carril 1, Figura 3). Sustituyendo en la ecuacion los valores de migracion de las 2 bandas obtenidas en la digestion del ADN de la linea de supertilapias IG/03-F70, se obtuvo que el valor estimado de los pesos moleculares de dichas bandas fueron de 260 y 231 pb, respectivamente. Los valores obtenidos se asemejan a los publicados por Franck y colaboradores en 1992 (16), que fueron de 237 y 230 pb para las especies de O. hornorum y O. placidus. Teniendo en cuenta el patron que muestra el ADN satelite de la linea de supertilapias IG/03-F70, la no existencia de reportes sobre la introduccion de O. placidus en Cuba y los resultados obtenidos en los estudios de morfometria que se presentan en el presente articulo, consideramos que el fondo genetico de la linea estudiada es basicamente de la especie O. hornorum. Agradecimientos Los autores agradecen el trabajo realizado por el personal de la empresa Acuadique en la seleccion y coleccion de los animales utilizados para el estudio. Este trabajo ha sido financiado parcialmente por el Centro Internacional de Ingenieria Genetica y Biotecnologia. Collaborative Research Programme (proyecto CRP/CUB93-05). References 1. Gourene G, Teugles G. Differenciation morphologique de souches des tilapias Oreochromis niloticus et O. aureus (Teleostei; Cichlidae) utilisees en pisciculture. Cybium 1993;17(4):343-355. 2. Majumdar KC, McAndrew BJ. Relative DNA content of somatic nuclei and chromosomal studies in three genera: Tilapia, Sarotherodon and Oreochromis of the tribe Tilapiini (Pisces, Cichlidae). Genetica 1986;68:175-188. 3. Trewavas E. On the cichlid fishes of the genus Pelmatochromis with proposal of a new genus for P. congicus on the relationship between Pelmatochromis and Tilapia and recognition of Sarotherodon as a distinct genus. Bull Brit Mus Natur Hist Zool 1973;25:1-26. 4. Trewavas E. Generic grouping of Tilapiini used in aquaculture. Aquaculture 1982;27:79-81 5. Trewavas E. Tilapiine fishes of genera Sarotherodon, Oreochromis and Danakilia. 1983;583 pp. British Museum Natural History, London. 6. Morales A. La tilapia en Mexico. Biologia, cultivos y pesquerias. AGT Editor S.A. 1991;3, 15. 7. McAndrew BJ, Majumdar KC. Evolutionary relationship within three tilapiine genera (Pisces: Cichlidae). Zool J Linn Soc 1984;80:421- 435. 8. Sodusk P, McAndrew BJ. Molecular systematic of three tilapiine genera: Tilapia, Sarotheredon and Oreochromis using allozyme data. J Fish Biol 1991; 39(SA):301-308. 9. Carbone P, Vitturi R, Catalano E, Macaluso M. Chromosome sex and Y- autosome fusion in Blennius tentacularis Brunnich, 1765 (Pisces: Blennidae).J Fish Biol 1987;31:597-602. 10. Nanda I, Feichtinger W, Schmid M, Schr”der JH, Zischler H, Epplen T. Simple repetitive sequences are associated with differentiation of the sex chromosomes in the Guppy fish. J Mol Evol 1990;30:456-462. 11. Chourrout D, Itskovich J. Three manipulations permitted by artificial insemination in tilapia: induced diploid gynogenesis, production of all triploid populations and intergeneric hybridization. In: Fishelson L, Yaron Z, eds. International Symposium on Tilapia in Aquaculture, Tel Aviv University 1983; 246-55. 12. Chourrout D. Induction of gynogenesis, triploidy, and tetraploidy in fish. ISI ATLAS of Science: Animal and Plant Science 1988;65-70. 13. Varadaraj K, Pandian TJ. Production of all female-sterile-triploid Oreochromis mossambicus. Aquaculture 1990; 84:117-123. 14. Rab P, Roth P, Mayr B. Karyotype studdy of eight species of European Percid fishes (Pisces: Percidae). Caryologia 1987;40(4):307-318. 15. Turner TF, Ahlquist JE, White MM, Kelsey R. Variation in genome organization and composition of repeated DNAs among three species of fish (Family: Percidae). The Journal of Heredity 1993; 84(1):70-73. 16. Franck JPC, Wright JM, McAndrew BJ. Genetic variability in a family of satellite DNAs from tilapia (Pisces: Cichlidae). Genome 1992;35:719- 725. 17. Franck JPC, Wright JM. Conservation of a satellite DNA sequence (SATB) in the tilapiine and haplochromine genome (Pisces: Cichlidae). Genome 1993; 36:187-194. 18. Harris AS, Bieger S, Doyle RW, Wright JM. DNA fingerprinting of tilapia, Oreochromis niloticus, and its application to aquaculture genetics. Aquaculture 1991;92:157-163. 19. Carter RE, Mair GC, Skibinski DOF, Parkin DT, Beardmore JA. The application of DNA fingerprinting in the analysis of gynogenesis in tilapia. Aquaculture 1991;95:41-52. 20. Martinez R, Estrada MP, Berlanga J, Guillen I, Hernandez O, Cabrera E et al. Growth enhancement in transgenic tilapia by ectopic expression of tilapia growth hormone. Molecular Marine Biology and Biotechnology 1996;5(1):62-70. 21. De la Fuente JM, Martinez R, Estrada MP, Hernandez O, Cabrera E., Garcia del Barco D et al. Towards growth manipulation in tilapia (Oreochromis sp.):generation of transgenic tilapia with chimeric constructs containing the tilapia growth hormone cDNA. J of Marine Biotechnology 1995;3:216-219 22. Guillen I, Martinez R, Hernandez O et al. Aquaculture Biotechnology Symposium Proceedings. Edited by Edward M. Donalson and Don. Mackinlay. International Congress on the Biology of Fishes. San Francisco State University. July 14-18. 1996:63-72. Copyright 1998 Elfos Scientiae The following images related to this document are available:Line drawing images[ba98002a.gif] |
| |||||||||

{kind=link}
{kind=link}