 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 1998;15:149-153 CHARACTERIZATION OF THE GENETIC BACKGROUND OF THE IG-91/03F70 STRAIN OF SUPERTILAPIA. II. ISOENZYMES *Reynold Morales,1 Georgina Espinosa,2 Yaisel J Borrell,2 Leopoldo Tápanes,3 Mario P Estrada1 and José de la Fuente1 1 Mammalian Cell Genetics Division, Center for Genetic Engineering and Biotechnology, PO Box 6162, Havana, Cuba. Phone: (53-7) 21 8164; Fax: (53-7) 21 8070; E-mail: jose.delafuente@cigb.edu.cu2Biology Faculty, University of Havana, Havana, Cuba. 3National Aquaculture Enterprise, Cotorro, Havana, Cuba. *Corresponding author Received in June, 1997. Accepted for publication in April, 1998.
Code Number: BA98022
ABSTRACT Considering the difficulties in the identification of tilapia species, electrophoretic analyses of enzymes and proteins enable aquaculturists to distinguish among species throughout the world. In the present study, we used a reliable method independent of the morphological characteristics to study the phenotype of the transgenic line of supertilapia IG-91/03F70. The results support the previous morphological, meristic and DNA studies, where the IG-91/03F70 tilapia line showed high correlations with Oreochromis hornorum and suggested other experiments to analyze a possible genetic input from O. niloticus. Genetic variation was observed at 26 loci among the supertilapia IG-91/03F70 population. Key words: tilapia, isoenzymes, electrophoretic markers RESUMEN Las dificultades para la identificación de las distintas especies de tilapias existentes en el mundo, son resueltas por los acuiculturistas mediante los análisis de los patrones electroforéticos de enzimas y proteínas. En el presente estudio se utiliza un método confiable independiente de las características morfológicas, para conocer la naturaleza de la línea de tilapias transgénicas IG-91/03F70. Los resultados obtenidos apoyan los encontrados previamente en los estudios morfológicos, merísticos y de ADN, donde la línea supertilapia IG-91/03F70 mostró alta correlación con la especie de tilapia Oreochromis hornorum y sugiere otros experimentos para aclarar el posible aporte génico de la especie O. niloticus. Se observó una variación genética en 26 loci entre la población de la línea supertilapia IG-91/03F70. Palabras claves: tilapia, isoenzimas, marcadores electroforéticos Introduction Aquaculture is one of the highly perspective ways of improving the protein balance in human feeding. In our country, hundreds of small reservoirs were constructed in which diverse fish species may be cultured (1). The species of high commercial value cultivated in Cuba were introduced in the country because there were no rapid growing native species with good reproductive performance. Among the species introduced, one of the largest populations was that of tilapia (2). Several introductions have taken place in Cuba since the '60s, with the farming of certain species (T. rendalli, Oreochromis mossambicus, O. aureus and O. niloticus). In 1979, two hybrids of tilapia known as "red tilapia" and "golden perch" were introduced from the Philippines. More recently, the O. hornorum was introduced (3). The expansion of intensive cultures and mainly of tilapias (Oreochromis spp.) has underlined the need of using different methods for controlling reproduction and attaining high yields. The main ones are: manual sex selection, sexual reversion using steroids and hybridization. The latter, either naturally, or directed by man, has widely propitiated the contamination of cultured stocks. The development of molecular biology and biotechnology has offered new methods to increase yields in aquaculture. Gene transfer, particularly that related with growth, has produced increases in size and weight of transgenic tilapia (4). The use of this technology in our country has enabled the production of a tilapia line which is known as the line of Supertilapia IG-91/03F70 (4). This line shows improved weight gain and a genetic background of hybrid species which have not been totally characterized. Several authors have demonstrated the use of electrophoretic methods for the identification of tilapia species and have reported diagnostic loci for certain species of this group (5-8). In this paper we complete the information on the genetic background of the line of Supertilapia including, besides the results of the morphometric studies and DNA analyses (9), the characterization of the electrophoretic patterns of several enzymes and proteins and the degree of genetic variability as well as its comparison with the reports from the literature. The results obtained support the hypothesis stated in previous studies by Morales et al (9), where we propose a large genetic contribution of the O. hornorum to this transgenic tilapia line. Other results do not allow to discard the genetic contribution of the O. nilotipus to the hybrid that constitutes the line of Supertilapia IG-91/03F70. Materials and Methods This study was performed with 20 non-sexed young fish of the Supertilapia line IG-91/03F70 randomly captured at the Guanimar experimental station in Havana province, Cuba. The enzymatic allelic variation was examined for 26 protein loci corresponding to 7 enzymes and 2 other proteins. After their extraction, the samples were stored at 20 ºC. The cold macerated protein extracts from the liver, white muscle, eyes, blood and plasma were prepared in a 1:2 ratio (weight:volume) in a Tris phosphate buffer solution (0.06-0.03 mol/L, pH 7) (10). The samples were submitted to a vertical electrophoresis according to Davis (11), using polyacrylamide gels that varied according to the system under study (Table 1). The electrophoresis were performed with Tris-lysine buffer (0.005-0.042 mol/L, pH 8.3), in a Truveller and Nefyodov equipment (12). Table 1. Enzymatic systems and loci studied in different tissues of the supertilapia line IG-91/03F70.
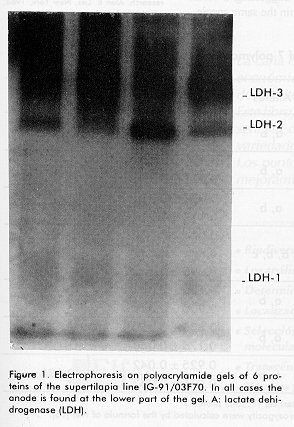
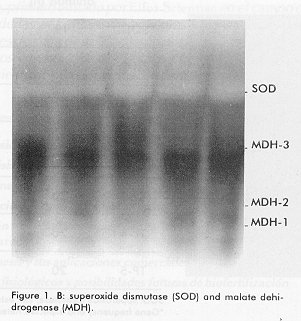
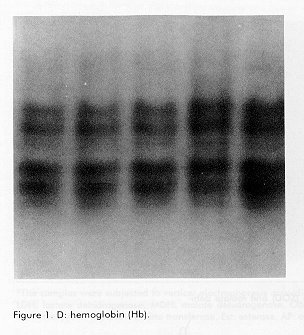
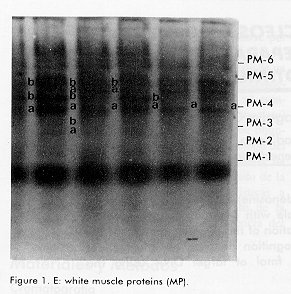
*The samples were subjected to vertical electrophoresis according to Davis (11), using polyacrylamide gels. The gels were incubated with specific stains for the development of enzymatic activity according to Shaw and Prassad (13). Gene frequencies and the Hardy Weinberg genetic equilibrium for the polymorphic system, comparing the observed and expected phenotypes by a chi square test for polymorphic systems, were determined, as well as the observed and expected heterozygocity, the percentage of polymorphic loci and the number of alleles/locus. Results and Discussion The enzymes and proteins studied with reproducible and clear allelic expressions are shown in Figure 1 and Table 1. In the case of systems with more than one locus, the fastest one was designated with the number 1. In the case of polymorphic systems, the alleles were named with a small letter starting by "a" for that of the greatest mobility. All bands analyzed migrated towards the anode. Figure 1. A: Electrophoresis on polyacrylamide gels of 6 proteins of the supertilapia line IG-91/03F70. In all cases the anode is found at the lower part of the gel. A: lactate dehidrogenase (LDH). Figure 1. B: superoxide dismutase (SOD) and malate dehidrogenase (mdh). Figure 1. C: white muscle esterase (Est). Figure 1. D: hemoglobin (Hb). Figure 1. E: white muscle proteins (MP). Of the 26 loci analyzed, 19 were monomorphic, which corresponded to the lactate, malate and glucose-6-phosphate dehydrogenases, aspartate amino transferase, alkaline phosphatase and esterase enzymes. The hemoglobin and certain total protein bands of the muscle also showed to be monomorphic. The lactate dehydrogenase enzyme had three loci (Figure 1A, Table 1), the most anodal one, characteristic of the eye, was also present in the plasma. A similar number of loci have been reported in the literature for representatives of the tilapines (5, 6, 8). Three loci were identified in the malate dehydrogenase enzyme (Figure 1B, Table 1), all monomorphic. McAndrew and Majundar (5) found for the majority of the species, a pattern of 5 bands and a pattern of 3 bands for Tilapia zillii and O. niloticus (5). Also, other authors have reported three loci for this enzyme in different species among which are O. niloticus and O. mossambicus (3, 6). Sodsuk and McAndrew (5, 8) have reported only 2 loci for this enzyme. The fact that the pattern obtained in the line of Supertilapias IG-91/03F70 for the malate dehydrogenase, agrees with that reported for O. niloticus and that, on the other hand, the electrophoretic behavior of the glucose-6-phosphate dehydrogenase with only one locus (Table 1) agrees with that reported by Macaranas et al (6) in populations of O. niloticus and O. mossambicus, leads to the idea that the line studied carries O. niloticus genes. Alternanively, the pattern of O. hornorum for these enzymes (which has not been reported), could be similar to the pattern described for O. niloticus. The aspartate amino transferase showed an electrophoretic pattern of 2 loci. The most anodal one was present in the muscle and the one with the least mobility was found in the liver (Table 1). The distribution of the loci in tissues differed from the results obtained by Macaranas et al (6), which can be attributed to the use of different experimental systems in these two studies. The alkaline phosphatase presented a poorly active locus that may agree with the acid phophatase reported by Sodsuk and McAnrew (8). The change in pH in the staining may be the cause of the low activity found for this enzyme in the present study. A complex pattern of bands were found in the esterases, that were clearly interpreted as 8 loci, five of which were monomorphic (Figure 1). Other authors (5, 6, 8) only report 2 or 3 systems of esterases in tilapia species, possibly due to the resolution of the electrophoretic support they use which does not allow the clear detection of the rest. The electrophoretic pattern for hemoplobin was of 4 bands, which agrees with that reported by Camacho et al for O. hornorum and O. mossambicus (Figure 1D) (3). These authors found a different distribution of bands for the 2 species mentioned, with the pattern of the line of Supertilapia IG-91/03F70 agreeing with that published for O. hornorum. Three of the systems detected with general protein staining in the white muscle were monomorphic and the general pattern obtained largely corresponded with that reported by Camacho (3) for tilapia rendali, golden perch, O. hornorum, red tilapia, O. aureus and O. mossambicus. Uribe et al reported in 1989 a similar pattern for the species O. mossambicus, O. hornorum and their hybrids (14). The other 7 loci obtained with the superoxide dismutase and esterase enzymes and with the total proteins of the muscle showed a polymorphic behavior. In all cases, comparing the phenotypes observed with those expected according to the Hardy-Weinberg law, the differences were non significant, thereby confirming the genetic interpretation given to the behavior of these loci. The superoxide dismutase had only one locus (Figure 1B) on the same gel as the malate dehydrogenase. This locus showed the greatest heterozygocity of all the polymorphic loci. In general, this result was similar to that published by other authors (3, 5, 8). The three esterase polymorphic loci identified as Est-2, Est-4 and Est-5 presented 2 and 3 alleles and their frequencies are shown in Table 2. The esterase polymorphic loci were reported by McAndrew and Majumdar (5) but only for the species O. aureus, O. jipe and O. niloticus. Macaranas et al also refered to an esterase polymorphic locus in O. niloticus but not in O. mossambicus, that were the two species studied by these authors (6). The other 3 polymorphic loci corresponded to the total muscle proteins (Figure 1E, Table 2). Macaranas reported in 1986 (6) 2 polymorphic systems for the species O. niloticus and O. mossambicus for this marker and Camacho reported in 1984 the possibility of finding polymorphism in the central region of the pattern of total proteins (3). Gene frequencies of 7 polymorphic loci in the supertilapia line IG-91/03F70.
SOD: superoxide dismutase; Est: esterase; TP: total protein. The average heterozygocity attained for the line IG-91/03F70 was of 0.063 and that expected was of 0.060. These heterozygocity values were calculated for several tilapia species by McAndrew et al (5), who obtained values ranging from 0.020 to 0.058. The latter value corresponds to the O. niloticus. On the other hand, the average heterozygocity obtained by Macaranas was of 0.073 for O. niloticus and 0.022 for O. mossambicus (6), suggesting a certain variability in this parameter within the same specie. The average heterozygocity obtained for the line IG-91/03F70 is found between the values obtained for O. niloticus by the groups of McAndrew and Macaranas (5, 6), suggesting a certain relationship between both species, without excluding the possibility that the values for O. niloticus and O. hornorum may be very similar since for the latter no values have been reported. The heterozygocity obtained in the line of Supertilapias IG-91/03F70 was also similar to those re-ported by Shaklee in 1983 for other groups of fish (16). The percentage of polymorphic loci was of 26, which is inferior to that of 42.8 found by Macaranas in O. niloticus and O. mossambicus (5). Finally, we can conclude that the results obtained from the study of the electrophoretic patterns in hemoglobin and proteins of the white muscle for the line IG-91/03F70 support the hypothesis stated in previous studies by Morales et al (9), where we state the high genetic contribution of the O. hornorum to the line studied. On the other hand, the results of the analysis of other proteins, enzymes and parameters as average heterozygocity and percentage of polymorphic loci, do not lead to the rejection of a genetic contribution of the O. niloticus to the hybrid under study (Supertilapia IG-91/03F70), which requires the comparison of the values obtained with those of the pure O. hornorum. Acknowledgments The authors wish to thank the workers of the "Acuadique enterprise" of Havana Province for the selection and harvest of the animals used for this study. This work was partially supported by the International Centre of Genetic Engineering and Biotechnology Collaborative Research Program (Project CRP/CUB93-05). References
Copyright 1998 Elfos Scientiae The following images related to this document are available:Photo images[ba98022d.jpg] [ba98022c.jpg] [ba98022e.jpg] [ba98022b.jpg] [ba98022a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}