 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
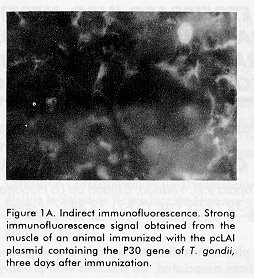
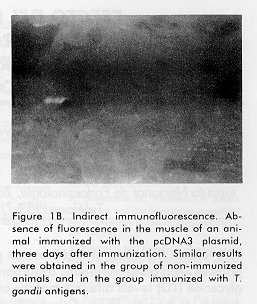
Biotecnología Aplicada 1998;15:159-161 EXPRESSION OF THE MAJOR SURFACE ANTIGEN OF Toxoplasma gondii IN THE MUSCLE OF BALB/C MICE * Alberto Fachado,1 Jorge L González,1 Carlos Hidalgo,2 Dinorah Torres,3 Teresita Vidal,2 Lisset Fonseca,3 Esteban Alberti,3 Ana M Montalvo,3 Léster Leal,4 Marcelo Nazábal,5 Regina Amendoeira5 and Armando Acosta2 1Departamento de Inmunología, Instituto Superior de Ciencias Básicas y Preclínicas
"Victoria de Girón", ave 31 y 146, No. 3102, Playa, Ciudad de La Habana 16, Cuba. E-mail: fachado@Vgiron.sld.cu Received in February, 1998. Accepted for publication in March, 1998.
Code Number: BA98024
ABSTRACT The gene, corresponding to the mature protein, of the major surface trophozoites antigen of Toxoplasma gondii (P30) was cloned in the pcDNA3 plasmid under the cytomegalovirus intermediate early promoter and used for the intramuscular immunization of Balb/c mice. The expression of the P30 antigen in the muscle of immunized animals was evaluated by indirect immunofluorescence. The expression of P30 antigen in the muscle of immunized animals was detected three days after immunization whereas in animals immunized with pcDNA3 plasmid and in non-immunized animals the results were negative. Key words: Toxoplasma gondii , P30 antigen, DNA, immunization RESUMEN El gen del principal antígeno de superficie del trofozoito de Toxoplasma gondii (P30) fue clonado en el plásmido pcDNA3 bajo el promotor temprano de citomegalovirus y usado para la inmunización intramuscular de ratones Balb/c. La expresión del antígeno P30 en el músculo de los animales inmunizados fue evaluada tres días después por inmunofluorescencia indirecta. Se detectó la presencia del antígeno en los animales inmunizados, no así en los otros grupos. Palabras claves: Toxoplasma gondii, antígeno P30, ADN, inmunización Introduction Infection with the Toxoplasma gondii parasite is responsible for toxoplasmosis in humans and other warm-blooded animals (1). This infection is benign for healthy people although it may cause abortion or neonatal malformations if contracted during pregnancy (2). This disease is often lethal for immunocompromised patients as those with AIDS, neoplastic diseases or transplanted organs (2). In veterinary medicine, toxoplasmosis has economic importance due to cattle abortion and neonatal loss (3). Thus, the development of an effective vaccine against T. gondii would be of great value to both human and veterinary medicine. Cell-mediated immunity is an important mechanism of host protection against the obligate intracellular parasite T. gondii as evidenced by the high incidence of toxoplasma infection in immunosupressed individuals. The major surface antigen of T. gondii (P30) has the ability to induce antigen-specific cytotoxic T cells in mice (4). In this study, these cells were directly lytic for extracellular RH strain tachyzoites. P30 (SAG1) induces a wide polyisotypic antibody response during infection (IgM, IgA, IgG) (5, 6). Two reports highlighted the prophylactic potential of P30, since they have simultaneously demonstrated that immunization with this protein in the presence of liposomes (7) or Saponin Quil A (8) induces nearly total protection of mice infected with a moderately virulent T. gondii strain. One of the most promissory strategies for the development of new generation vaccines against parasitic infections is DNA immunization (9-11). The first step in that goal is to achieve the expression of the specific gene in the target tissue. In the present paper, we demonstrated the expression of the P30 antigen in the muscle of Balb/c mice immunized with the gene coding for this antigen. Materials and Methods Animals Fifteen Balb/c mice, six to eight weeks old, supplied by the National Center for the Production of Laboratory Animals (CENPALAB, Havana, Cuba) and maintained in standard conditions, were used in the experiment. Parasites Tachyzoites of the highly virulent RH strain of T. gondii (kindly provided by Dr. Pascal Herion, Institute of Biomedical Science, Mexico City) were used for the experiment. T. gondii tachyzoites were obtained from the peritoneal exudate of 3 to 4 days infected Balb/c mice as described elsewhere (12). Bacterial strain The Escherichia coli GC-366 strain (71/18 dam-. P1 Transduction from GC-352 dam 13 : : Tn9 (cloranfenicolr) D (lac-pro) F' {lac Iq D lac Z-M15 pro+} was used for genetic manipulation. Transformed and non transformed GC-366 E. coli strain cells were cultured in a Luria Bertaini medium (LB) (1 % Triptone, 0.5 % yeast extract and 0.5 % NaCl) with and without ampicillin (50 mg/mL), respectively. The cultures were incubated at 37 ºC overnight with agitation (200 rpm). Restriction and modification enzymes The enzymes Pst I, Sma I, Xba I, T4 DNA Ligase, Klenow, TAq DNA polymerase and Alkaline phosphatase (Boehringer Mannheim, Biochemical) were used according to the manufacturer's instructions. DNA purification Genomic DNA of T. gondii tachyzoites was obtained as follows: 0.5 x 107 tachyzoites were resuspended in 1 mL of lysis buffer (100 mM tris-HCl pH 8, 100 mM EDTA pH 8, 2 % SDS, 150 mM NaCl and 200 mg/mL of proteinase K) and incubated 2 h at 56 ºC. After phenol-chloroform extraction, the DNA was precipitated with absolute ethanol and then resuspended in the Tris-EDTA (TE) buffer (Tris 10 mM, EDTA 1 mM pH 8). The DNA integrity was evaluated by agarose gel electrophoresis 0.8 % with ethidium bromide (13). DNA concentration was estimated with the Gene Quant Apparatus (Pharmacia Biotech). Polymerase chain reaction The PCR was performed as described by Joss et al. (14). Each PCR reaction contained 1 U of TAq polymerase enzyme, 1x buffer (10x concentrated) (20 mM Tris-HCl, 0.1 mM dithiothriotol, 0.1 mM EDTA, 0.1 M KCl, 0.5 % nonidit P40 (v/v), 0.5 % Tween 20 (v/v), 50 % glycerol (v/v) pH 8), 0.2 mM each of dATP, dCTP, dGTP, dTTP (Advanced Biotechnologies) and 40 pmols of each primer. The 0.794 Kb fragment of the P30 gene, corresponding to the mature protein, was amplified from 1 m L (100 ng/µL) of the T. gondii genomic DNA in a final volume of 50 m L as described (14) using the following set of primers: 5'-GAAGATCTAGACTTCACTCTCAAGTGCCCT AAAACAGCGCTC-3' (upper primer) and 5'-GGACTTATTGGCTCTATCGCAGCTTGTGTC CCGGGATA-3' (lower primer). All primers were from the Center for Genetic Engineering and Biotechnology (CIGB, Havana, Cuba). The reaction was carried out with the Gene ATAQ Controller Apparatus (Pharmacia) with one cycle of 95 °C for 5 min, 55 °C for 40 s and 72 °C for 1.30 min, followed of 30 cycles of 94 °C for 50 s, 55 °C for 40 s and 72 °C for 1.30 min, with a final cycle of 72 °C for 5 min. pcLAI plasmid construction The PCR product was digested with the restriction enzymes Xba I and Sma I and then ligated to the previously Xba I digested pcDNA3 plasmid (INVITROGEN), which carried the intermediate early promoter of the cytomegalovirus. After filling the remaining Xba I site of the plasmid with the Klenow enzyme, a second ligation reaction was carried out to obtain the pcDNA3-P30 recombinant plasmid (pcLAI). Competent E. coli cells were transformed with the ligation mixture (13). The plasmid with the correct insert orientation was detected by restriction analysis with the Pst I enzyme. Once identified, the pcLAI plasmid was highly purified by the alkaline lysis method (13). The purified material was diluted in sterile NaCl 0.9 % at a final concentration of 250 mg/mL, aliquoted and stored at - 20 ° C until use. Immunization protocol Animals were divided into 4 groups and inoculated as follows: 100 mg of purified pcLAI (n = 5), 100 mg of plasmid pcDNA3 (n = 5), 100 mg of T. gondii antigen (n = 5) and an additional group of 5 animals that remained non-immunized. All animals received the inoculation in a volume of 0.1 mL of NaCl 0.9 % by the intramuscular route in the internal region of the right thigh. Indirect immunofluorescence study Three days after the immunization, the animals were slaughtered and tissue sections of 4 mm of the inoculated muscle were cut by cryostat (1720 Leitz). The samples obtained from each animal were incubated with a high titer human serum against the T. gondii antigenic extract determined by ELISA (15), or with a pool of negative sera tested by the same method as a negative control. Both sera were used diluted 1:16 in phosphate buffered saline solution, pH 7.2 (PBS). After 30 min the slides were washed three times for 5 min with PBS and were incubated for 30 min with a goat anti-human immunoglobulin G fluorescein isothiocyanate conjugate (BIOCEN, Cuba), diluted 1:40. After three washes of 5 min in PBS, the samples were mounted with a solution of glycerol PBS (9:1) and observed in a fluorescence microscope (Leizt, Germany) with a magnification of 25X. Results and Discussion Cell-mediated immunity is an important mechanism of host protection against the strictly intracellular parasite T. gondii. The major surface antigen of T. gondii (P30) has the ability to induce antigen-specific cytotoxic T cells in mice (4). The intramuscular injection of naked DNA in saline solution allows the long term expression of the expressed antigen. We injected intramuscularly the P30 mature T. gondii protein under the control of the cytomegalovirus intermediate early promoter. It is known that antigens that triger CTL response induce an immune-mediated destruction of transfected muscular cells, and after 5 days destruction is of 80 % with HBsAg (16). The detection of the expression of P30 in the muscle was carried out after 3 days of injection to allow the expression and avoid the possible effect of a CTL response. In the muscle of the animals immunized with the pcLAI plasmid strong fluorescent staining was observed (Figure 1A). The signal observed varied among the different cross-sections of the same animal, the deeper ones having the strongest signal. No significant differences were observed between the animals of this group. In contrast, the remaining control groups gave negative results (Figure 1B), as expected. Figure 1A. Indirect immunofluorescence. Strong immunofluorescence signal obtained from the muscle of an animal immunized with the pcLAI plasmid containing the P30 gene of T. gondii, three days after immunization. Figure 1B. Indirect immunofluorescence. Absence of fluorescence in the muscle of an animal immunized with the pcDNA3 plasmid, three days after immunization. Similar results were obtained in the group of non-immunized animals and in the group immunized with T. gondii antigens. The samples of the animals of all groups gave negative results after incubation with the pool of negative sera (data not shown). The fact that negative results were obtained with the muscle of the animals of the control groups after incubation with the high titer anti-toxoplasma serum, excludes the possibility that the positive results obtained in the group of animals immunized with pcLAI were due to the presence of cross reactive antibodies directed to normal constituents of the muscular cells in this serum. On the other hand, the negative results found in all groups with the pool of negative sera rule out the possibility that the result obtained in the group immunized with the fragment P30 gene were associated with non-specific reactions. The level of expression depends on the antigen, and the expression vector. We used the complete open reading frame of the protein, 336 amino acids and the pcDNA3 vector to obtain a wider immune response and a long term expression in the muscle. The results obtained demonstrates the expression of the P30 antigen of T. gondii in the muscle of the immunized animals. The humoral response of immunized mice with a DNA fragment containing the carboxyl-terminal 288 amino acids of P30 in the pCMVint vector was recently reported (17). In this short report they only measured the humoral response in the immunized mice, after 12 weeks, against the T. gondii total extract. There was no direct evidence of P30 expression in the muscle. The vector and gene fragment used are different to ours, so the immune response and the expression level might be different. The expression of other parasitic antigens in the muscle of mice immunized with naked DNA has been demonstrated (9, 10). Our group showed, by indirect immunofluorescence, the expression of Trypanosoma cruzi antigens in the muscle after the intramuscular immunization of mice with a genomic expression library of this parasite (11). The humoral response is elicited between 20 to 60 days and the cellular response could take longer. We are currently evaluating the capability of the polynucleotide vaccination with a plasmid encoding the complete P30 protein to elicit humoral and cellular response in mice. Acknowledgements The authors are grateful to Dr. Ricardo Silva for his helpful suggestions, Dr. Gerardo Guillén for reviewing the manuscript and to Jorge L. Leyva for technical support. References
Copyright 1998 Elfos Scientiae The following images related to this document are available:Photo images[ba98024b.jpg] [ba98024a.jpg] |
| |||||||||

{kind=link}
{kind=link}