 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 1998;15:167-172 STRUCTURAL ANALYSIS OF THE 14-3-3 PROTEINS BY LC-ESIMS: CHARACTERISATION OF SEVEN 14-3-3 ISOFORMS ISOLATED FROM HUMAN PLACENTA * Joel Madrazo,1 Steven Howell,2 David Jones, 2 Harry Martin2 and Alastair Aitken2 1Center for Genetic Engineering and Biotechnology. PO Box 6162, Havana, Cuba. Fax: (53-7) 21 8070; 33 6008; E-mail:protchem@cigb.edu.cu Received in November, 1997. Accepted for publication in January, 1998.
Code Number: BA98026
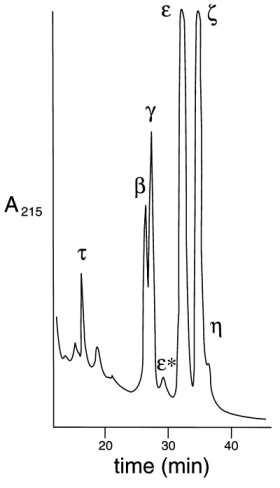
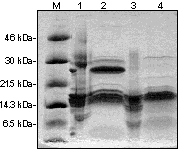
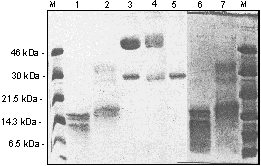
ABSTRACT The 14-3-3 is a family of acidic dimeric proteins with a pI of about 4.5 and a mass of 30 kDa on sodium dodecyl sulfate-polyacrylamide gel electrophoresis. There are seven major mammalian brain isoforms of 14-3-3 proteins, named a to h . 14-3-3 is highly conserved over a wide range of mammalian species. Homologues of 14-3-3 have also been found in plants, insects, amphibians and yeast. Here we present the structural analysis of 14-3-3 proteins with femtomole sensitivity by liquid chromatography-electrospray ionization mass spectrometry (LC-ESIMS). A reverse phase capillary column was set up for on-line trapping of peptides/proteins for concentration and desalting. Controlled digestion of native 14-3-3 with subtilisin was fractionated by gel filtration and fractions were analysed by ESIMS. It was shown that the N-terminus is an intact domain involved in the 14-3-3 dimerisation for all isoforms. Crosslinking experiments of the isolated N-terminal domain confirmed its dimeric association for all the isoforms isolated from sheep brain. The characterisation of 14-3-3 isolated from human placenta showed the presence of six known brain isoforms: t , b , g , e , z and h . A new isoform e * was found that was recognised by the antiserum specific to the e isoform. It had a molecular mass of 14 Da higher than e , but lower retention time on reverse-phase high performance liquid chromatography. Molecular masses of 14-3-3 monomers isolated from human placenta were measured. We found that a and d , which are in vivo post-translationally phosphorylated forms of b and z respectively in the mammalian brain, are absent in the placenta. Key words: 14-3-3 proteins, human placenta, dimers, protein domains, controlled digestion, ESIMS, femtomole sensitivity RESUMEN Las 14-3-3 constituyen una familia de proteínas acídicas diméricas con un pI cercano a 4,5 y una masa de 30 kDa en geles de poliacrilamida con dodecilsulfato de sodio. En cerebro de mamíferos existen siete isoformas mayoritarias de estas proteínas nombradas a , b , ..., h , las cuales son altamente conservadas en un amplio espectro de especies de mamíferos. Se han encontrado también homólogos de las 14-3-3 en plantas, insectos, anfíbios y levaduras. Aquí presentamos el análisis estructural de las 14-3-3 mediante espectrometría de masas con ionización por electronebulización acoplada a cromatografía líquida (LC-ESIMS) con sensibilidad femtomolar. Una columna capilar de fase inversa fue empleada en linea para la preconcentración y desalado de los péptidos y las proteínas analizados. Mediante digestiones controladas de las 14-3-3 nativas con subtilisina fue posible obtener un dominio que fue aislado por filtración en gel y analizado por ESIMS. Se demostró que el extremo NH2 de estas proteínas es un dominio que está involucrado en la dimerización de todas las isoformas de las 14-3-3. Los experimentos de entrelazamiento de este dominio permitieron confirmar su asociación dimérica para todas las isoformas aisladas de cerebro ovino. La caracterización de las 14-3-3 aisladas de placenta humana demostró la presencia de al menos seis de las isoformas de cerebro: t , b , g , e , z y h . Se encontró una nueva isoforma e *, la cual es reconocida por los antisueros específicos para e . El valor de la masa molecular medido por ESI es 14 Da mayor que esta última, mientras es menos retenida en la cromatografía de fase inversa. Se obtuvieron los valores de masas moleculares para las isoformas aisladas de placenta humana y encontramos que las isoformas a y d , que corresponden a las formas fosforiladas in vivo de b y z respectivamente en cerebro de mamíferos, están ausentes en placenta. Palabras claves: proteínas 14-3-3, placenta humana, dímeros, dominios, digestión controlada, ESIMS, sensibilidad femtomolar Introduction The 14-3-3 is a family of N-acetylated acidic proteins with a pI of about 4.5 and molecular masses of about 30 kDa on sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). They were named due to the migration position on diethylaminoethyl- (DEAE) cellulose chromatography and starch gel electrophoresis (1). The increasing interest in the literature to 14-3-3 is associated with the finding of their many diverse functions including critical roles in signal-transduction pathways by associating with at least three proteins which regulate signalling: 14-3-3 proteins were found to bind to the middle T transforming protein of polyoma virus (2), to the Raf-1 protein kinase (3-5) and to Bcr (6). Today they are more accepted as a novel type of chaperone protein that modulates interactions between components of the signalling pathways (7). They are also involved in exocytocis and cell-cycle control for DNA damage in fission yeast (8), and regulation of protein kinase C (PKC) in mammalian cells, reviewed by several authors (9-11). The 14-3-3 proteins are widely distributed in nature, with highly homologous proteins having been found in a variety of eukaryotic organisms including plants (12), insects (13), amphibians (14), birds (15) and yeast (16). To date, seven mammalian brain isoforms of 14-3-3 have been characterised, named a to h , according to their sequential elution position after reverse-phase high performance liquid chromatography (rp-HPLC) (17, 18). Five of these have been sequenced (8), and a /d isoforms are identical in primary structure to b / z isoforms respectively, but differ only in a post-translational modification (19). Comparison of all the 14-3-3 protein sequences reveal that they are highly conserved, some of them contain a few conservative substitutions over a wide range of mammalian species. These proteins all have an apparent molecular mass on SDS-PAGE from 29 to 33 kDa and exist as dimers (20). In Mammals, the 14-3-3 family of proteins have been found to be expressed in low levels in nearly all tissues examined to date, including pineal glands, retina (21), adrenal chromaffin cells (22), human platelets (23) and human placenta (as described in this report); neural tissue appears to be somewhat more enriched (24, 25). It has been estimated that these proteins represent approximately 1 % of the total soluble brain protein (24). The 14-3-3 proteins have been reported to be localised in neurones, and to be axonally transported to nerve terminals in axonal ganglion cells (26). Two isoforms found, one in T-cells (t ) (27) and another in epithelial cells (stratifin), (28) or HME1 (29), have also been described. Although a global unifying physiological function of these proteins has not been clearly established, it has been postulated that these proteins are involved in a variety of cell functions (8). Firstly, Ichimura and co-workers in 1987 (30) showed that 14-3-3 was identical to an activator protein of tyrosine and tryptophan hydroxylases, the rate limiting enzymes involved in catecholamine and serotonin neurotransmitter biosynthesis, respectively. More recent studies indicate that purified sheep brain 14-3-3 isoforms do not activate recombinant tyrosine hydroxylase (31). Other functions proposed for the 14-3-3 family include the control of ADP-ribosyltransferase (5) and the modulation of PKC activity (8). A 14-3-3 protein called Exo1 stimulates calcium dependent exocytosis in permeabilized adrenal medulla cells (22). In mammalian pineal gland, N-acetyl transferase shows a 100-fold increase in activity at night, which increases serotonin to melatonin conversion, thereby suggesting a function for the 14-3-3 in regulating diurnal rhythm. In the present study, we isolated and purified the 14-3-3 proteins present in human placenta and identified the different isoforms using a battery of polyclonal antibodies raised against peptides corresponding to the N-terminus of brain 14-3-3 isoforms (19). Here we present the analysis with femtomole sensitivity by liquid chromatography-electrospray ionization mass spectrometry (LC-ESIMS) of enzymatically cleaved native N-acetylated 14-3-3 proteins. The 14-3-3 proteins present in human placenta were isolated, purified, and the isoforms were characterised. Materials and Methods All reagents were of analytical grade, from BDH, Sigma or Boehringer. Protein assay reagent was obtained from BioRad. HPLC and fast performance liquid chromatography (FPLC) solvents and water were obtained from Romil. Purification of 14-3-3 Expression and purification of the recombinant t 14-3-3 isoform was as described elsewhere (32). 14-3-3 was isolated from sheep brain and human placenta by a combination of anion-exchange and hydrophobic interaction chromatography, according to the methods of Toker et al. (20). A similar protocol with certain modifications was used in the case of human placenta. Freshly collected human placenta was cut into small pieces and stored frozen at -80 ºC until purification of the 14-3-3. Two pieces (200 g) of placenta were thawed and homogenised for 1 min at maximum setting in an ultraturrax homogeniser in 600 mL of extraction buffer [20 mM tris pH 7.5, 0.25 M sucrose, 2 mM ethylenediaminetetraacetic acid (EDTA), 10 mM ethylene glycol- O,O'-bis (2-aminoethyl) -N,N,N',N'-tetraacetic acid (EGTA) and 1 mM 1,4-dithio-DL- threitol (DTT)] to which a pool of protease inhibitors (phenylmethanesulfonyl fluorid (PMSF) 10 mM, leupeptin 10 mg/L, pepstatin A 10 mg/L, soybean trypsin inhibitor 10 mg/L, and benzamidine 10 mM) had been added immediately before use. All procedures were carried out at 4 ºC. After centrifugation at 40,000 g for 30 min the supernatant was applied onto a DEAE-Sepharose column (300 mL), using a Pharmacia FPLC system, previously equilibrated with buffer A (20 mM tris pH 7.5, 1 mM each EDTA, EGTA and DTT). The column was extensively washed with buffer A and bound proteins were eluted by the application of a linear gradient (0-0.5 M NaCl) from buffer A to B (buffer A with 0.5 M NaCl) at a flow rate of 3.5 mL/min. Fractions of 14 mL were collected and each fraction of 0.2 to 0.4 M NaCl was analysed by SDS-PAGE. Fraction containing 14-3-3 (30 and 33 kDa bands) were pooled, NaCl was added to a final concentration of 2.5 M and loaded on a 50 mL phenyl-sepharose column equilibrated with 2.5 M NaCl in buffer A. Proteins were eluted by applying a 60 min step gradient (2.5 M-1.5 M-0.5 M-0.0 M NaCl in buffer A) at 2 mL/min. Collected fractions of 5 mL were analysed by SDS-PAGE; 14-3-3 was eluted from the column only when buffer A (0.0 M NaCl) was applied. Pooled fractions were desalted by a Sephadex G25 column and finally applied on a Pharmacia mono Q HR column equilibrated with buffer A. A 60 min gradient (from 0.0 to 0.5 M NaCl) at 2 mL/min was run to elute the proteins, fractions were analysed by SDS-PAGE and pooled 14-3-3 were separated by reverse phase HPLC. Separation on reverse phase HPLC of 14-3-3 isoforms Separation was carried out in a Waters gradient HPLC system with a UV detector at 215 nm wavelength. A purified mixture of 14-3-3 proteins from sheep brain (or partially purified from placenta) from mono-Q fractions was applied on a reverse phase analytical column (C4 Bakerbond 4.6 x 250 mm). Proteins were eluted at 0.8 mL/min flow rate by a pre-programmed convex gradient (No.4) of 35-55 % (water/ acetonitrile/ TFA- reverse phase solvent system) during 40 min. This gradient allows a rapid increase of acetonitrile concentration from 35 % to approximately 45 %, and then a very slow increase from 45 to 55 % B (0.05 % TFA in acetonitrile). Peaks were collected manually and different isoforms of 14-3-3 were identified by dot and Western blot techniques using a battery of specific antibodies against the N-terminus of the 14-3-3 isoforms (19). Controlled digestion of 14-3-3 The recombinant purified t isoform as well as the mixture of 14-3-3 purified from sheep brain was digested with subtilisin at a ratio 1/50 (enzyme / substrate) at 4 ºC. After 20 h, the subtilisin was inactivated by addition (two times) of PMSF freshly dissolved in dimethylsulfoxide (DMSO). The digested proteins were analysed by 15 % SDS-PAGE and Western blot. Digestions were also run on gel filtration chromatography on a Superose 12 column (Pharmacia) with phosphate buffer saline (PBS) pH 7.2, flow rate 0.3 mL/min, and detection by absorbance at 280 nm, 0.05 AU. Several peaks corresponding to 60, 30 and less than 30 kDa were collected and analysed directly on the LC-ESIMS coupled through a reverse phase capillary column. An additional separation of the obtained peptide mixtures to get the mass spectra of the component peptides was not necessary. Acrylamide gel electrophoresis of proteins and Western blot analysis Proteins were electrophoresed in denaturing conditions by the method of Laemmli (33) and the Western blot analysis was performed as described by Martin et al. (19). ElectroSpray ionization mass spectrometry ElectroSpray ionization mass spectrometry (ESIMS) was performed on a Fisons VG Platform instrument. On-line trapping was used to purify and desalt peptides/proteins before introduction to the ESIMS source (34). This comprised a 0.7 mm microbore column (slurry-packed in house) with Polymer Labs. (UK) poly (styrene/divinylbenzene) PLRP-S, 8 m m particle, 300Å pore size. The sample was loaded on this trapping column in a low concentration of organic solvent, washed free of interfering salts, TFA, etc. with acetonitrile/water/acetic acid 15:84:1 (v/v/v) at a flow rate of 2-500 m L min-1. Proteins were eluted (in ca. 80 % yield) with acetonitrile/water/acetic acid 50:49:1 (v/v/v) at a flow rate of 10 m L min-1 by switching a Rheodyne valve to set this column on-line with the source. This resulted in a very large increase in sensitivity, down to a few (50-100) femtomoles of protein. Crosslinking of the 14-3-3 proteins For crosslinking studies, two water-soluble homobifunctional reagents of different spacer lengths were used: bis (sulfosuccinimidyl) suberate (BS3, 11.4 Å) or ethylene glycobis (sulfosuccinimidylsuccinate) (Sulfo-EGS, 16.1 Å), both from Pierce (USA). The crosslinker was added to the protein solution (1-1.5 mg/mL) in phosphate buffer, pH 8, to reach a final concentration of 2-3 mM. Protein solution was vigorously agitated and left for 40 min at room temperature. Then the reaction was stopped by the addition of 1 M tris buffer pH 7.5 to a final concentration of 50 mM. As a control experiment, the undigested 14-3-3 proteins and a monomeric protein (carbonic anhydrase) were evaluated under the same reaction conditions. Results and Discussion Phosphorylated forms of 14-3-3 are absent in placenta Figure 1 shows the separation by rp-HPLC of 14-3-3 proteins isolated from human placenta. Most of the isoforms that have been characterised in mammalian or avian brains are present in the placenta. Seven peaks were identified as isoforms of 14-3-3 proteins (t , b , g , e , e *, z , h ), by the dot blot technique with a battery of specific antibodies against the N-terminal decapeptide of each brain isoform (19). Some other peaks present in this fraction correspond to contaminant proteins from the placenta tissue. This was verified by dot- and Western blot with a non isoform-specific 14-3-3 polyclonal antiserum, which recognises all 14-3-3 isoforms (data not shown). Here the first peak identified as a 14-3-3 protein corresponded to t . Although its mass spectra showed that it is N-acetylated as all native 14-3-3 (42 Da higher than the theoretical mass value, Table 1), it elutes on rp-HPLC just before the b isoform (Figure 1), earlier than the recombinant t and the native N-acetylated t isoform found in the brain, (19). This could be due to one or a combination of aminoacid substitutions with similar masses but lower hydrophobicity. Figure 1. Separation of 14-3-3 proteins isolated from human placenta on rp-HPLC. The small peak that eluted just before e (Figure 1) (identified as e *) was recognised by the antiserum that is specific to the N- and C-terminus of e isoform (not shown). It showed no readable sequence on the Edman-degradation automatic sequencer. This suggests that the N-terminus is blocked (acetylated), as are all 14-3-3 native isoforms studied to date. On SDS-PAGE it migrates as e (33 kDa band) and ESIMS analysis showed that it is approximately 14 Da higher in mass than e (Table 1). These results are consistent with a Thr/Asp change at position 38 observed for this isoform isolated from the sheep spleen or from its brain (...DVELThr/AspVE...), (37). These evidences exclude the possibility that e * is a proteolytic product of e by an exopeptidase during purification. Further analysis has to be done to confirm the structural differences between the two homologue (e ) isoforms. Table 1. Molecular masses of 14-3-3 isoforms isolated from the placenta.
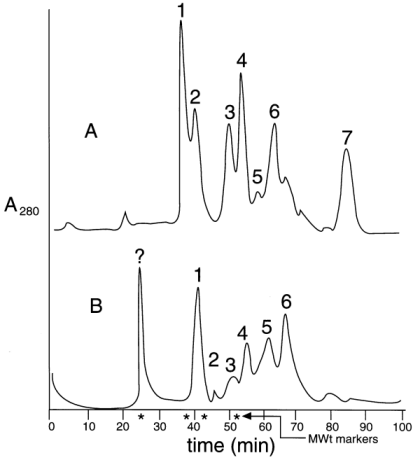
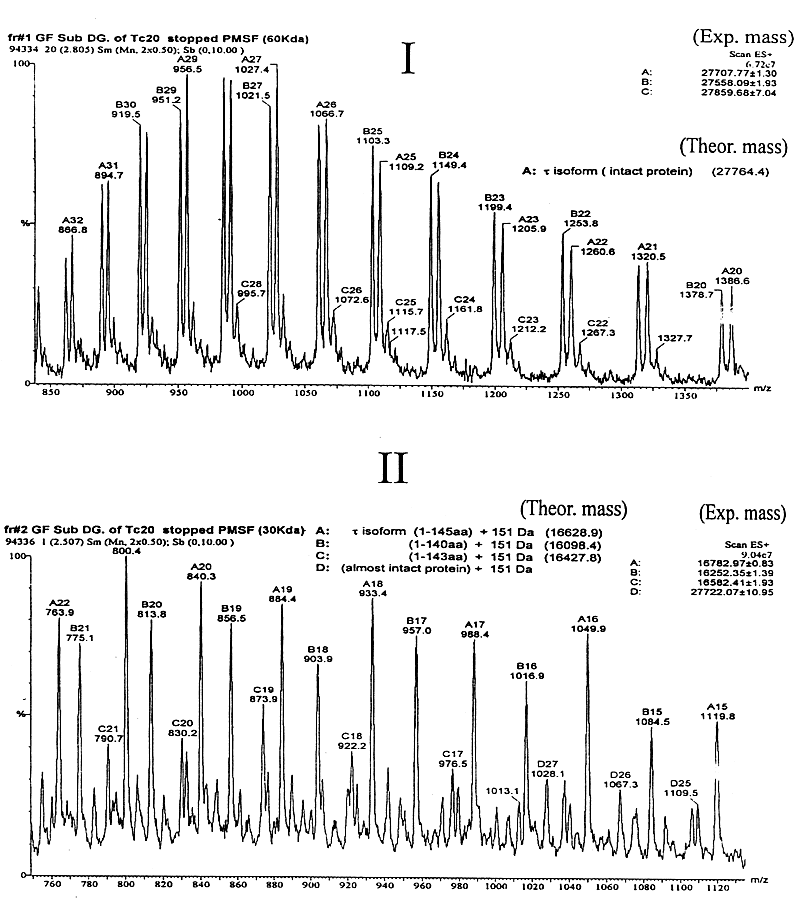
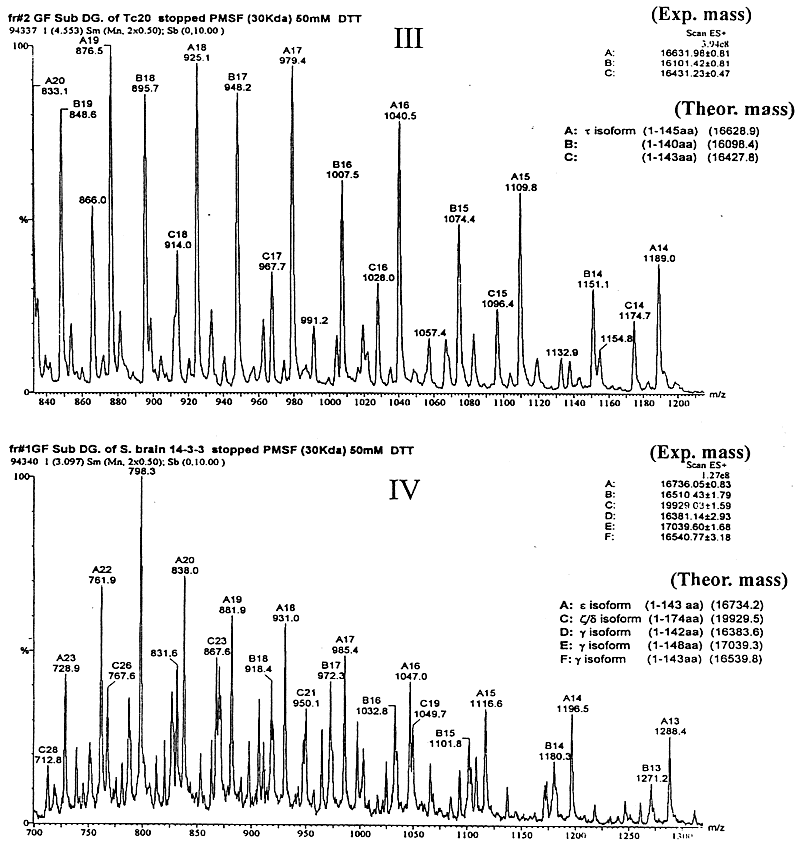
*: calculated masses from DNA sequences; ** b T is a form of b (see text). It has been previously established by protein sequencing that two isoforms, a and b (as well as d and z ) from mammalian brain 14-3-3 are identical in primary structure, but differ only in a post-translational modification (phosphorylation), which is responsible for the mass difference of 80 Da and for the observed earlier elution of a and d (with respect to b and z ) isoforms on rp-HPLC (19, 38). In addition, this relationship between a / b and d / z 14-3-3 isoforms in both mammalian and avian brains had been confirmed by ESIMS (38). The measured masses of the human placenta 14-3-3 isoforms (which are all N-acetylated proteins) are shown in Table 1. b T is a form of b that uses the alternative methionine codon six nucleotides upstream of the major initiation site which is processed to give an N-terminal N-acetyl-Thr as the first residue instead of N-acetyl-Met (28). This accounts for the mass difference of 101 Da (Thr residue) between them. These two forms of b co-elute in one peak on rp-HPLC (38).From the results of dot- and Western blot, rp-HPLC separation and ESIMS data we can conclude that there is no evidence of an a or d isoform, which are in vivo post-translationally phosphorylated forms of b and z respectively. They have been found in brain tissue (19, 38) but are absent in human placenta. 14-3-3 N-terminus is involved in dimerisation Most globular proteins in their native conformation are relatively resistant to the attack of proteases at low concentrations. Controlled digestion under certain experimental conditions can give valuable information on the conformation and structure of the substrate protein or a complex of proteins. The digestion of sheep brain 14-3-3 with a variable concentration of subtilisin at 37 ºC produced mainly a fragment of 17-kDa (32). Under those conditions, it was shown that an antibody specific for the N-terminus of e isoform was able to recognise this 17-kDa fragment by Western blot, but the digestion to isolate this domain in native conditions was impossible to control. Complementary experiments carried out in different conditions for the digestion (described in Materials and Methods) allowed the isolation and analysis of this resistant domain of the 14-3-3 proteins. Separation by gel filtration of the digested recombinant t isoform and the digested 14-3-3 protein mixture isolated from sheep brain on a Superose 12 column are shown in Figure 2. Peak 1 (Figure 2A) corresponds to the almost undigested dimeric t isoform (» 60 kDa). ESIMS analysis (Figure 3I) of this fraction showed mass values of PMSF-adducts to the t free Cys residues, which are relatively close to the theoretical mass for the intact monomeric t isoform (27764.2 Da). SDS-PAGE (Figure 4) and Western blot with the antiserum raised against the N-terminal decapeptide of t isoform (not shown) confirmed the integrity of the t N-terminus. Peak 2 (Figure 2A), eluting at 30 kDa on gel filtration is, according to the ESIMS analysis (Figure 3II, III) and SDS-PAGE (Figure 4), a mixture of at least three sTable peptide products of about 16 kDa. All of them had an intact N-terminus as determined by dot blot and were dimeric, eluting on gel filtration with a mass twice (» 30 kDa) the amount measured by ESIMS (» 16 kDa). Peaks 3, 4 (Figure 2A) and others were identified as mixtures of many small peptides from the C-terminal domain of the protein (data not shown). Figure 2. Separation of 14-3-3 digested with subtilisin by gel filtration on a Superose-12 column. A: subtilisin digestion of an t isoform; B: separation of the digested 14-3-3 mixture of sheep brain isoforms. Molecular weight markers: 2000 kDa blue dextran, 67 kDa bovine serum albumin (BSA), 30 kDa carbonic anhydrase, 12.5 kDa cytochrome-C. Figure 2B shows similar results for the subtilisin digestion of the 14-3-3 mixture of brain isoforms. In this case, the digestion went on for 30 h and the 60 kDa peak that corresponds to the dimeric intact 14-3-3 proteins could not be observed. Here peak 1 (» 30 kDa) is, according to the ESIMS analysis (Figure 3IV), a mixture of N-terminal fragments of several isoforms with a range of masses between 16 and 19 kDa. Many of them matched closely the theoretical masses of the N-terminal peptides of the most abundant isoforms in the mixture: e , z (d ) and g , respectively (Figure 3IV). SDS-PAGE (Figure 4) and Western-blot analysis with each isoform-specific antiserum also confirmed this result (not shown). Figure 3. Mass spectra of 14-3-3 fragments obtained by controlled subtilisin digestion and separated by gel filtration FPLC. I: spectrum of peak 1 (Figure 2A), eluting as a 60 Da protein, corresponds to some PMSF adducts (+151 Da) of the monomeric t isoform almost intact; II: spectrum of peak 2 (Figure 2A), which eluted as a 30 Da protein, represents PMSF adducts (+151 Da) of a mixture of at least three N-terminal peptides 1-140, 1-143 and 1-145 of t . Figure 3. Cont. III: spectrum of peak 2 (Figure 2A) after treatment with 50 mM DTT for 30 min to reduce the PMSF adducts from Cys residues. Here, measured masses match closely (+/- 3 Da) the expected values for peptides terminating at Q140, D143 and S145 of t isoform; IV: spectra of DTT treated peptides from peak 2 (Figure 2B), eluting as a 30-kDa protein on gel filtration. Mass values obtained crorrespond to a mixture of N-terminal peptides from the most represented isoforms in sheep brain: three N-terminal g peptides, one e N-terminal peptide and one d /x N-terminal peptide. Figure 4. Analysis by 15 % SDS-PAGE of digested 14-3-3 proteins with subtilisin in order to generate and isolate a resistant domain. Lane M: molecular weight markers; lanes 1, 2: t isoform digested at 4 ºC for 20 h; lanes 3, 4: 14-3-3 proteins from sheep brain digested at 4 ºC for 30 h; and lanes 1, 3: samples were run with a non reducing sample buffer. In Figure 2B, a peak eluting at the void volume of the column (marked as "?") was collected. It was undetecTable on SDS-PAGE. This probably corresponds to insoluble PMSF, added in excess to inactivate the subtilisin, which aggregates from the oversaturated solution. Crosslinking experiments Two water-soluble homobifunctional crosslinking reagents of different spacer lengths were evaluated: BS3 and Sulfo-EGS. From them, we selected BS3 because, under the evaluation conditions this reagent produced better yields from crosslinking. Figure 5 shows the analysis by SDS-PAGE of the crosslinked 14-3-3 N-terminal fragments that were isolated by gel filtration chromatography after the subtilisin digestion. The appearance of bands above the monomeric N-terminal fragments (at the height of their double mass) confirms their dimeric state in native conditions as observed by gel filtration. Control experiments with native dimeric 14-3-3 proteins showed similar results, that is the appearance of the band at the height of the dimeric form. For a known monomeric protein (carbonic anhydrase) used as a negative control for crosslinking experiments, no band was observed at the height of its double mass. Figure 5. Analysis by 15 % SDS-PAGE of the crosslinked 14-3-3. Lanes M: molecular weight markers; lane 1: isolated N-terminal domain of the t isoform; lane 2: crosslinked N-terminal domain of t ; lane 3: crosslinked non digested t isoform; lane 4: crosslinked non digested 14-3-3 mixture from the brain; lane 5: crosslinked carbonic anhydrase as a control; lane 6: N-terminal domain generated from the mixture of all 14-3-3 isoforms isolated from sheep brain; and lane 7: crosslinked N-terminal of all 14-3-3 isoforms isolated from sheep brain. All 14-3-3 proteins show high levels of identity in sequence and so they are likely to have nearly identical tertiary structures. We can state that the 14-3-3 N-terminus corresponds to a domain involved in the dimerization of this family of proteins. A few months later after this study was completed, the crystal structure of two of the isoforms (t and z ) of the 14-3-3 family was published (35, 36). For both isoforms, the crystal structure shows that they, in fact, exist as dimers, in which each monomer is composed of an unusual arrangement of nine antiparallel a -helices organised as two structural domains. The N-terminus is, in fact, one domain that is involved in the dimerisation of the proteins, with residues 5-21 in one monomer being in contact with residues 58-89 in the opposing monomer. These results show that the strategy to obtain a 14-3-3 resistant domain (involving the interacting site) was valid and could be applied for other structural protein studies where the tridimensional structure is not yet known. Acknowledgements We would like to acknowledge the financial assistance from Wellcome Trust in the fellowship for J. M. References
Copyright 1998 Elfos Scientiae The following images related to this document are available:Line drawing images[ba98026d.gif] [ba98026b.gif] [ba98026c.gif] [ba98026f.gif] [ba98026e.gif] [ba98026a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}