 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 1998;15:183-187 EXPRESIÓN POR INGENIERÍA GENÉTICA DE LA FIMBRIA K99 DE Escherichia coli ENTEROTOXIGÉNICA EN DIFERENTES CEPAS DE E. coli K12 *Ángela E Sosa,1 Roberto Basulto,2 José A Ramón2 y José de la Fuente3 1División de Desarrollo Biofarmacéutico, 3División de Genética de Células de Mamíferos, Centro de Ingeniería Genética y Biotecnología, AP 6162, Ciudad de La Habana, Cuba, Telefax: (53-7) 21 8070 / 33 6008; E-mail: alberto.agraz@cigb.edu.cu Recibido en enero de 1998. Aprobado en abril de 1998. Code Number: BA98030 ABSTRACT The fimbriae is used for immunization of lambs, pigs and calves, against gastrointestinal infection with Escherichia coli K99. Due to low expression levels of fimbriae in natural strains, it is attractive its obtainment by recombinant technology. However, it is difficult to achieve a stable expression in E. coli K12, because of the lisis and the plasmid unstableness which provoke the accumulation of the proteins that constitute the fimbriae inside a bacterium. We previously reported the cloning of the gene fanC on the plasmid pUC19. In this work, we report the cloning of the complete operon under its own regulating region on the plasmid pUC19. This genetic construction was evaluated in 14 strains of E. coli K12, and several differences were found in terms of: the efficiency of transformation, growth and levels of expression of fimbriae (determined by a specific ELISA) in the Luria-Bertani, minimum and saline media. We also evidenced that the presence of the genotype D (ara-leu) favored the expression of fimbriae. These observations permitted us to adequately select the host, which is an important step in the large-scale expression of recombinant fimbriae. Keywords: fimbriae, pili, K99, enterotoxigenic E. coli RESUMEN La fimbria se utiliza para la inmunización de corderos, cerdos y terneros contra la infección gastrointestinal de Escherichia coli K99. Debido a los bajos niveles de expresión de fimbria a partir de cepas naturales es atractiva su obtención por vía recombinante. Sin embargo, es difícil lograr una expresión estable en cepas de E. coli K12, por la lisis celular y la inestabilidad plasmídica que provoca la acumulación de las proteínas que constituyen la fimbria en el interior de la bacteria. Previamente se reportó el clonaje del gen fanC en el plásmido pUC19. En este trabajo se reporta el clonaje del operón bajo su propia región reguladora en el plásmido pUC19. Esta construcción genética se evaluó en 14 cepas de E. coli K12 y se encontraron diferencias en cuanto a: la eficiencia de transformación, crecimiento y niveles de expresión de fimbria (determinada por un ELISA específico) en los medios Luria-Bertani, mínimo y salino. También se evidenció que la presencia de un genotipo D (ara-leu) donde se deleciona el operón leu completo favoreció la expresión. Estas observaciones permitieron hacer una selección adecuada del hospedero, paso importante para la expresión de fimbria recombinante a gran escala. Palabras claves: fimbria, pili, K99, E. coli enterotoxigénica Introducción Las cepas patógenas de Escherichia coli logran establecerse en el hospedero por una fuerte adherencia a la superficie de las mucosas y tienen la capacidad de infectar el tracto respiratorio, urinario o gastrointestinal. El proceso de adherencia está basado en el reconocimiento de componentes específicos de la membrana plasmática en las células del tejido del huésped y está mediado por una estructura adhesiva llamada fimbria o pili, localizada en la superficie de la bacteria (1). La fimbria de cepas K99 es el principio activo de vacunas contra la enterocolibasilosis porcina, ovina y bovina (2). La biosíntesis de la fimbria es un proceso complejo que requiere de una serie de eventos moleculares secuenciales que sólo ocurren durante la infección (3). En las cepas K99 naturales, el operón fan que contiene los ocho genes necesarios para la formación de la fimbria es portado en una simple copia en un episoma conjugativo de aproximadamente 52 MDa (4). La regulación local del operón fan ocurre por las proteínas FanA y FanB que actúan como antiterminadores de la transcripción (5). Debido a la actividad de estas proteínas, el gen fanC que codifica la subunidad antigénica más importante de la fimbria K99 es expresado de forma constitutiva a muy bajos niveles. Este operón se encuentra dentro de la familia de los operones que están bajo el control del regulador global de la proteína de respuesta a leucina (LRP) (5). Este regulador estimula la expresión de la fimbria al unirse al promotor fanA; es por esto que la expresión de fimbria como una estructura es sólo posible bajo ciertas condiciones ambientales (6). Se ha evidenciado que la expresión de Lrp se induce en la fase logarítmica del crecimiento celular en medio mínimo y, por el contrario, se reduce notablemente en medio rico o en medio salino suplementado con 1 % de casaminoácidos (5). Por ello, es muy difícil obtener grandes cantidades de biomasa fimbriada cuando se utilizan cepas naturales de E. coli con fines productivos. Por otra parte, los inconvenientes propios de la manipulación de patógenos hacen de la vía recombinante una alternativa adecuada para la expresión de fimbrias para ser empleadas en vacunas. La estrategia más común para la expresión de proteínas recombinantes a gran escala es la clonación en vectores plasmídicos de alto número de copia y/o bajo el control de la regulación de promotores metabólicos fuertes que permitan la síntesis de altos niveles de transcriptos, como primera etapa en lograr altos niveles de la proteína de interés. Cuando se clonó el gen fanC bajo la regulación del promotor triptófano (ptrp) y en el vector pUC19 (500-700 copias/célula), se observó que la acumulación de la proteína provoca inestabilidad plasmídica y lisis celular en las cepas HB101, JM103 y W3110 (7, 8). Sin embargo, no se conoce si alguna característica en el genoma del hospedero pudiera favorecer la expresión de la fimbria. En este trabajo se reporta el clonaje del operón completo bajo su propia región reguladora en el plásmido pUC19. Además, se estudiaron 14 cepas de hospederos de E. coli K12 con diferentes genotipos con respecto al operón leucina, con el objetivo de evaluar si este genotipo influye en el rendimiento final de la biomasa fimbriada. La comparación se hizo en cuanto a: la eficiencia de transformación, el crecimiento y los niveles de expresión de fimbria en los medios Luria-Bertani (LB), mínimo y salino suplementado con casaminoácidos. Materiales y Métodos Cepas de E. coli K12 utilizadas en este trabajo
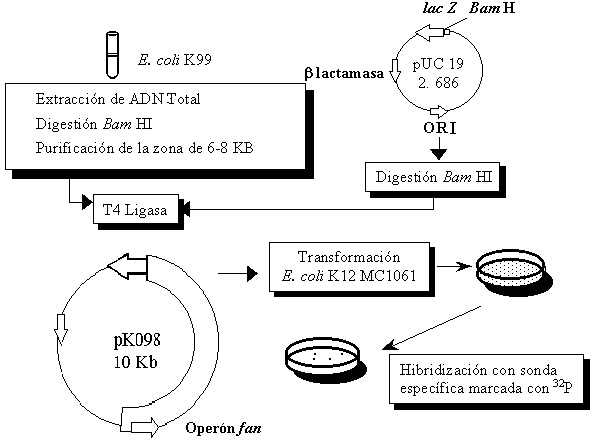
Clonaje del operón fan en el plásmido pUC19 A partir de una cepa de E. coli K99 positiva se extrajo el ADN total, se digirió con la enzima de restricción BamHI y se construyó una minilibrería en el plásmido pUC-19. La estrategia de clonaje se describe en la Figura 1. Todas las enzimas fueron adquiridas de New England BioLabs, Inc. Los procedimientos para el aislamiento y manipulación del ADN se hicieron de acuerdo a Sambrook et al. (9). La construcción final se llamó pK098. Figura 1. Estrategia seguida para el clonaje del operón fan (expresión de la fimbria K99) en el plásmido pUC-19. La cepa de E. coli enterotoxigénica K99 positiva fue aislada de cerdos enfermos (17), el ADN total fue aislado y digerido con la enzima BamHI siguiendo el método convencional. La zona entre 6 y 8 Kb se aisló de un gel de agarosa de bajo punto de fusión y se extrajo por el método de fenol/cloroformo. Paralelamente se digirieron 10 µg de pUC 19 con la enzima BamHI. Ambos fragmentos se ligaron y se transformaron en células de MC1061 hechas competentes por el método de CaCl2. Los transformantes que contenían la nueva construcción (pK098) se seleccionaron por hibridación con una sonda específica (17) marcada con 32P de las colonias transferidas a un filtro de nitrocelulosa. Los procedimientos para el aislamiento y manipulación del ADN se hicieron de acuerdo a Sambrook et al. (9). Transformación de cepas de E. coli K12 Las 14 cepas se transformaron simultáneamente con el plásmido pK098 que contiene el operón fan. Se utilizaron 100 m L de los cultivos crecidos a 37 ºC toda la noche en medio LB (9) para inocular 5 mL de medio LB y se crecieron 2 h a 37 ºC; luego 1 mL de los cultivos se centrifugó a 3 000 rpm, se lavaron con 250 m L de una solución fría de CaCl2 0,1 M, se decantó el sobrenadante y las células se resuspendieron en 100 m L de esta misma solución. A la suspensión celular se le adicionó 1 m g de ADN de la nueva construcción, se incubaron 30 min en hielo, luego se adicionó 1 mL de medio LB frío y se incubó sin agitación a 37 ºC por 1 h. Después de este tiempo se sembraron 100 m L de la suspensión celular por extensión en placas de medio LB al cual se adicionó ampicillina a una concentración de 100 mg/mL (LBA) para seleccionar los transformantes. Como control de viabilidad diluciones de 10-6 de todas las transformaciones se sembraron en placas LB. Como control del método se transformaron todas las cepas con el plásmido pUC19 sin el inserto. Condiciones de cultivo y expresión Los transformantes se crecieron en medios M9 y LB preparados según Sambrook et al. (9) y en medios Minca y salino preparados según Isaacson (16). Todos los medios se suplementaron con ampicillina (100 mg/mL) para la selección de los transformantes (M9A, Minca A y salino A y LBA). Determinación de la expresión y purificación de la fimbria La expresión se evidenció por un ensayo de aglutinación celular semicuantitativo a células intactas. Para este ensayo la cantidad de células en los cultivos fueron igualadas a densidades ópticas equivalentes y se mezclaron v/v en un portaobjeto con un suero policlonal en conejo originado contra fimbria K99 purificada a partir de la cepa de E. coli salvaje y diluido hasta 10-2. Después de mezclar el cultivo celular con el suero se cuantificó el tiempo en que comenzaron a observarse los agregados celulares y se tomó como mayor actividad el menor tiempo de agregación (17). La fimbria se purificó a partir de volúmenes de cultivo igual a 20/Abs530, lo cual es equivalente a 1-2 g de biomasa. Las células después de colectadas por centrifugación a 10 000 rpm se disolvieron en 1 mL de tampón fosfato 50 mM pH 5,5 (Na2HPO4 1,44g/L; KH2PO4 0,24 g/L, NaCl 9 g/L) y la mezcla se incubó a 60 ºC por 30 min. A continuación se centrifugó a 10 000 rpm y el sobrenadante se utilizó sin concentración previa en el ensayo inmunoenzimático (18) para determinar la cantidad de fimbria y se determinó la proteína total según el método descrito por Lowry et al. (19). Para cuantificar los niveles de expresión en cada experimento se utilizó un ensayo inmunoenzimático tipo ELISA basado en un anticuerpo monoclonal (AcM) contra la fimbria natural. Las placas de 96 pocillos se mantuvieron toda la noche a 4 ºC en 100 µL de una solución tampón carbonato/bicarbonato 0,l M (pH 9,6) que contenía 10 µg/mL del AcM CRC K99-2. Los pocillos se bloquearon adicionando leche descremada (0,05 %) en un tampón salino (PBS 1X ) (pH 7,2) y se incubó 1 h a 37 ºC. Luego la placa se lavó cuatro veces con Tween 20 (0,05 %), se añadieron 100 µL de diferentes diluciones del antígeno (fimbria) y se incubó durante 3 h a 37 ºC. Luego se lavó cuatro veces con Tween 20 (0,05 %), se añadieron 100 µL del anticuerpo K99-2 (1/10 000) conjugado con peroxidasa y se incubó 1 h a 37 ºC. Posteriormente se lavó cuatro veces con Tween 20 (0,05 %) y se reveló por adición de 100 µL de una solución de tampón citrato/fosfato (50 mM), 0,05 % de o-fenilendiamina y 0,05 % de peróxido de hidrógeno. La reacción se detuvo al añadir 50 µL de ácido sulfúrico 2N después de 30 min de reacción y se leyó en un multiscan (ScaJet IIp) a 492 nm (18). Resultados y Discusión Efecto de la expresión de la construcción pK098 sobre la frecuencia de transformación En este trabajo se estudió el efecto sobre la E. coli K12 de la expresión del operón fan clonado bajo su propia regulación en un plásmido de alto número de copias. Debido a que el operón fan es uno de los operones del regulón LRP evaluamos si el genotipo relativo al operón Leu influye en el rendimiento final de biomasa fimbriada. Se utilizaron 14 cepas de E. coli que comúnmente se emplean en la expresión de proteínas foráneas (Tabla 1). Tabla 1. Eficiencia de transformación de 14 cepas de E. coli K12 con la construcción genética pK098.
a Todas las cepas fueron transformadas de forma simultánea por el método rápido como se describe en Materiales y Métodos. Las frecuencias correspondientes a las cepas MC1061, AD494 y DHB-4 están referidas a las 16 h de incubación a 37 ºC. El resto de los transformantes crecieron después de 24 h de incubación. La frecuencia de transformación se calculó como el número de transformantes/mL/ug de ADN. bSe transformó una cantidad similar de pUC19 sin inserto (nd no determinado). cLa viabilidad corresponde a las colonias crecidas en placas de LB.Al transformar las diferentes cepas con la construcción pK098 se encontraron las mayores frecuencias en las cepas MC1061, AD494 y DHB-4; estas cepas presentan un genotipo D (ara-leu). En la cepa JM109 y JA221 con un genotipo leuB6, al igual que en las cepas con fenotipo Leu+, se observaron bajas frecuencias de transformación. Como control de este experimento se transformaron todas las cepas con el plásmido pUC19 sin el inserto, y se obtuvieron frecuencias entre 104-105 transformantes/m g de ADN. Las cepas transformadas se sembraron en placas de medio LBA, los transformantes de cepas con genotipo D (ara-leu) se obtuvieron a las 16 h de incubación a 37 ºC. Sin embargo, para el resto de las cepas los transformantes se observaron después de las 24 h de incubación. No se esperaba encontrar diferencias en el crecimiento en medio LBA. Una posible explicación de este fenómeno era que en estas cepas no se reprimía la expresión de la fimbria al crecer la células en medio rico. Se verificó esta hipótesis utilizando una dispersión de colonias obtenidas directamente de las placas de la transformación en un ensayo de aglutinación y se observó que todos los transformantes obtenidos a las 24 h fueron positivos. En las cepas JM101 y JM109, donde no se observaron transformantes, se supone que las células que adquieren el plásmido no son capaces de tener un número suficiente de duplicaciones como para que se observen colonias. Efecto de la expresión de la construcción pK098 sobre el crecimiento y la expresión de los transformantes El operón fan esta positivamente controlado por el regulador LRP, la leucina y la alanina inhiben la unión de esta proteína al promotor (5). Este regulador transcripcional media la respuesta de las bacterias en crecimiento a los cambios en la composición de nutrientes del medio. En las cepas naturales, la máxima expresión de LRP ocurre en medio mínimo. El medio LBA contiene altas cantidades de estos aminoácidos (0,99 mg/mL de leucina y 3,17 mg/mL de alanina) por lo que la síntesis de LRP se encuentra reprimida. El medio M9A se suplementó con los aminoácidos de auxotrofía de cada cepa a la concentración de 25 mg/mL, y el medio salino (49,5 mg/mL de leucina y 158,5 mg/mL de alanina) contiene 20 veces menos cantidad de aminoácidos que el LBA. La Tabla 2 muestra una comparación entre todas las cepas crecidas en los medios LBA, M9A y salino A en cuanto al crecimiento celular. Se observaron diferencias en el crecimiento de las cepas en estos medios. Los valores de crecimiento en medio mínimo o medio salino fueron muy bajos a diferencia del medio LBA. Se apreció en muchos casos lisis celular lo cual dificultó el ensayo de aglutinación y la purificación de la fimbria al volumen de trabajo empleado. Tabla 2. Comparación de las cepas de E. coli K12 transformadas con la construcción pK098.
a Donde se indica (lisis) se encontró debris celular entre las 3 y 6 h de cultivo; en estos casos no se pudo realizar el ensayo de aglutinación. bDeterminada por aglutinación. Se cuantificó el tiempo en segundos en que se observaron los agregados celulares; (++++) corresponde a una agregación instantánea tomada como la actividad máxima; (+++) la segunda cepa en observarse agregación y así en orden decreciente; (-) corresponde a las cepas donde no se observó agregación después de 30 min de contacto con el anticuerpo. cLas cepas fueron crecidas en medio salino. Los valores en por ciento son referido a la relación de los valores de Elisa (18) y proteínas totales determinadas por el método descrito por Lowry (19).La expresión se determinó en todos los medios por aglutinación y en el medio salino y LBA también fue cuantificada por ELISA. En el medio M9 no se observó expresión por aglutinación en ninguna de las cepas en que fue posible hacer el ensayo y en los medios LBA y salino la expresión obtenida fue similar con valores máximos del 30 % de la proteína obtenida en la extracción. Aunque el crecimiento en medio salino fue menor que en LBA se lograron valores similares de expresión según el ensayo de aglutinación (Tabla 2). En medio M9 no se encontraron células fimbriadas, sin embargo, se detectó la presencia de fimbria en los sobrenadantes de los cultivos de las cepas W3110 y MC1061 ensayadas por ELISA (resultados no mostrados). En la W3110 ocurrió la lisis del cultivo a las 4 h de crecimiento en todos los medios ensayados. Las cepas con fenotipo RecA- como la HB101 crecieron muy pobremente en todos los medios, lo cual no era esperado, ya que esta cepa ha sido utilizada por otros autores en la expresión de fimbria recombinante, aunque con plásmidos derivados de pBR322 el cual es de menor número de copias que el que se utilizó (7). En general, en las cepas con fenotipo salvaje para la síntesis de leucina se observaron bajas frecuencias de transformación y bajos niveles de expresión de fimbria. La expresión de la fimbria a niveles por encima de 5 mg/L de cultivo, la cual es equivalente al 30 % de la proteína total purificada, fue sólo posible en las cepas con genotipo D (ara-leu), Esta deleción se encuentra en el minuto 2 del mapa de ligamiento de E. coli K12 e incluye el operón araBDAC, el gen dadB y el operón leuBDCA, por lo que afecta el consumo de arabinosa y alanina y la síntesis de leucina. Las cepas con fenotipo Leu- no pueden crecer en medios limitados por leucina, por lo que la concentración de leucina intracelular estará siempre determinada por la disponibilidad de este nutriente en el medio de cultivo y no por la síntesis en el interior de la célula. Se supone que en estas cepas los niveles extracelulares de leucina modulan la actividad del regulador LRP, lo cual mantiene en todo momento ciertos niveles de represión y que la expresión depende fundamentalmente de la regulación local. La expresión de fimbria cuando los niveles de LRP fueron altos (medio mínimo) resultaron altamente tóxicos para la célula con la consecuente disminución de la velocidad de crecimiento y en muchos casos la lisis del cultivo. La expresión en medio LBA a los mismos niveles que en medio salino sugiere que la cantidad de transcriptos obtenidos a expensas de la regulación positiva local (FanA y FanB) asociado al alto número de copias del pUC19, fueron suficientes para la obtención de una cantidad de fimbria que no afectó la viabilidad de la célula. Conclusiones Los resultados permiten concluir que es posible obtener niveles aceptables de biomasa fimbriada mediante una selección adecuada de la cepa hospedera, teniendo en cuenta una estrategia de clonación que permita mantener la expresión de fimbria a niveles permisibles no tóxicos para la célula. Nuestros resultados confirman lo planteado por otros autores (7) acerca de la toxicidad que provoca la sobrexpresión de la fimbria en el hospedero. Sin embargo, se encontró que el efecto de toxicidad no se manifiesta igual para todas las cepas de E. coli K12. Las cepas con genotipo D (ara-leu) toleraron mejor una sobredosis de la proteína que las cepas con genotipo leuB6 o fenotipo Leu+. En general, cualquier estrategia a seguir debe considerar el control de los niveles de expresión de fimbria. En este caso ello se logró mediante el uso del control propio del operón en un plásmido de alto número de copias, por el mantenimiento de los cultivos entre ciertos niveles de represión de la regulación global empleando medios más ricos que los utilizados tradicionalmente en la expresión de fimbria y con cepas hospederas de E. coli K12 que presentan la deleción D (ara-leu). Estas observaciones permitieron hacer una selección adecuada del hospedero, paso importante para la expresión de fimbria recombinante a gran escala. Agradecimientos Quisiéramos agradecer al doctor Eugenio Hardy por la revisión crítica de este manuscrito, a Tania Olivera por su asistencia técnica y al trabajo de arbitraje realizado por la Revista. Referencias
Copyright 1998 Elfos Scientiae The following images related to this document are available:Line drawing images[ba98030a.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}