 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 1998;15:227-231 PCC2, A NEW TRANSPOSON-LIKE ELEMENT IN THE WHITE ROT FUNGUS Phanerochaete chrysosporiumBianca García,1 @ Manuel Raíces,1 Julio Delgado,1 Göran Pettersson2 1 Centro de Ingeniería Genética y Biotecnología. Ave. 31 entre 158 y 190, CP 10600, La Habana, Cuba. E-mail: banco@cigb.edu.cu2Department of Biochemistry, University of Uppsala, Uppsala, Sweden.
Code Number: BA98036
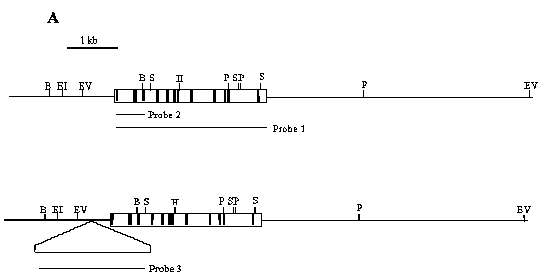
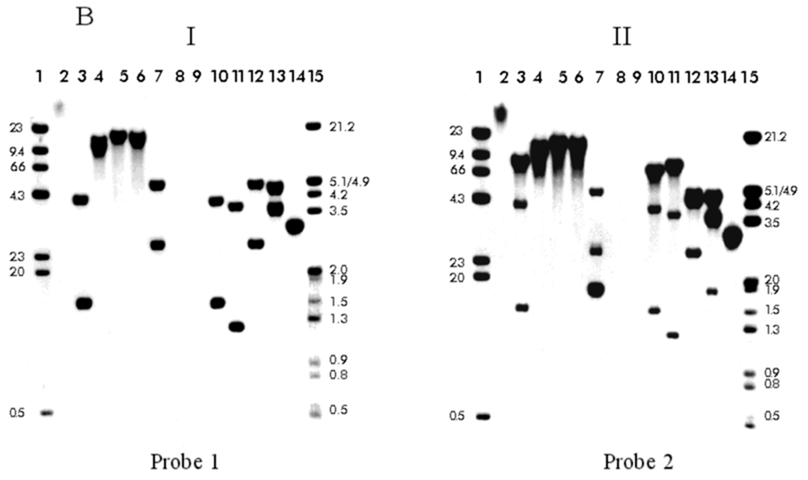
ABSTRACT The gene encoding cellobiose dehydrogenase from Phanerochaete chrysosporium K3 was isolated through PCR and its 3045 bp nucleotide sequence determined. Two distinct 5' flanking sequences were identified by Southern blot hybridization, both having a common 650 bp region, but one of them interrupted by a 2657 bp transposon-like element located at 425 bp upstream from the initial ATG. This element, named Pcc2, carried 14 bp inverted terminal repeats while lacking the conserved transposase motif it is flanked by a 2 bp (GT) target duplication site and is present at a high copy number throughout the P. chrysosporium K3 genome. Keywords: cellobiose dehydrogenase (CDH), gene, Phanerochaete chrysosporium, transposon-like element RESUMEN El gen codificante de la celobiosa deshidrogenasa de Phanerochaete chrysosporium K3 fue aislado mediante PCR y se determinó la secuencia nucleotídica de sus 3045 pb. Mediante hibridación por Southern blot se identificaron dos regiones 5' no codificantes diferentes, ambas contenían una región común de 650 pb, pero una de ellas estaba interrumpida por una secuencia de 2657 pb con características similares a un transposón. Este elemento, que se denominó Pcc2, está localizado a 425 pb del ATG de iniciación y contenía una secuencia de 14 pb repetida e invertida, sin embargo carecía del motivo conservado de las transposasas. El elemento está flanqueado por un sitio blanco de duplicación de 2 pb (GT) y está presente en un alto número de copias en el genoma de P. chrysosporium K3. Palabras claves: celobiosa deshidrogenasa (CDH), gen, Phanerochaete chrysosporium, transposón Introduction The white rot fungus Phanerochaete chrysosporium is one of the most efficient wood degrading fungi yet identified [1, 2]. In the last ten years this microorganism has been extensively characterized for biotechnological approaches based on a selective degradation of lignin and aromatic pollutants [3, 4]. In P. chrysosporium the enzymatic systems required for degradation of wood components, cellulose, hemicellulose and lignin, are complex and seem to act synergistically [5]. The cellobiose dehydrogenase (CDH) enzyme is one interesting element of this degradation system and it is secreted during the induction of the P. chrysosporium cellulolytic system [6]. CDH is a hemoflavoprotein which oxides cellobiose and higher cellodextrins to their corresponding lactones. Molecular oxygen may function as an electron acceptor producing hydrogen peroxide, but other compounds like quinones, ferricyanide and triodide ions are better [7]. The biological function of CDH is unknown, but several suggestions have been presented [8, 9]. Substantial variation has been observed among P. chrysosporium strains [10]. Here, during the isolation of the CDH promoter region, we detected an inserted DNA fragment which seems to be a transposon-like element. Transposable elements (TEs) are ancient and ubiquitous components of fungal genomes [10] and make up a substantial proportion of the total DNA in most, if not all, eukaryotic genomes [11]. Some of these elements have been shown to actively affect gene structure and function in several ways: inactivation of gene expression upon insertion, modification of the nucleotide sequence through excision, and probably by inducing extensive chromosomal rearrangements [10]. There are two main classes of TEs: the retrotransposons, that transpose through a process involving reverse transcription, and the transposons that move by an excision-insertion mechanism [11]. Retroelements have been found in a number of fungal species: Foret 1 in Fusarium oxysporum [12], Tad in Neurospora crassa (the first tranposable element reported in fungi) [13], Grasshopper, MGR583 and MAGGY in Magnaporthe grisea [14-16] and Boty in Botrytis cinerea [17]. However, DNA transposons have only recently been found: Fot1 and other DNA elements were identified in F. oxysporum [18-20], Pot2 and MGR586 in M. grisea [21, 22], Ant1 in Aspergillus niger [23] and Pce1 from P. chrysosporium [24]. CDH cDNA, genomic sequence and the deduced protein primary structure have been published [25-27]. Here we report the isolation of the 5' non-coding region of the CDH gene and the identification of a transposon like-element inserted in this region. Experimental procedures Strains and plasmids P. chrysosporium strain K3 was used in this study [28]. Escherichia coli strain DH5a (Bethesda Research Laboratories) was employed as the host for plasmids pBluescript SK (Strategene) and pT7BlueR (Novagen). E. coli was propagated in liquid or on solid Luria-Bertani medium at 37 °C. The plasmid pBluescript KS carrying the CDH cDNA [25] was used as a template in the PCR reaction. Gene isolation For the isolation of chromosomal DNA, 108 spores of P. chrysosporium were inoculated in liquid YPG medium (1% yeast extract, 2% bacto-peptone and 2% glucose, pH 5) at 37 °C for 48 hours. The mycelium was harvested, washed with water and the genomic DNA extracted as described by Raeder and Broda [29]. Primers located 5' (forward, 1F- 5'GCTTC GGTCGAGTTCACGATGCT) and 3' (reverse, 4R- 5'CAGCCCCGAGGACAGTAAT ATAAAG) to the translational start and stop codons, respectively, from the cDNA CDH sequences were used to isolate the genomic gene through PCR, using 100 ng of the genomic DNA as the template. The PCR conditions were 100 mM Tris.HCl (pH 8.3), 500 mM KCl, 1.5 mM MgCl2, 200 mmol of each deoxynucleotide triphosphate, 100 pmol of each primer and 2.5 U of Taq polymerase (Promega) in a final volume of 50 mL. The PCR reaction was carried out using the following program: 3 min at 94 °C, 1 min at 62 °C and 3 min at 72 °C, followed by 30 cycles of 1 min at 94 °C, 1 min at 62 °C and 3 min at 72 °C and a final extension step of 7 min at 72 °C. Finally, the PCR product was gel extracted by using the QIAEX II Agarose Gel Extraction kit (QIAGEN) and ligated to pT7Blue vector (Novagen). DH5a transformants were selected on LBA plus IPTG and Xgal, and a recombinant plasmid, pT7CDH, was chosen for sequencing. DNA methods DNA manipulations, including restriction digests, agarose gel electrophoresis, ligations, transformations of E. coli, and plasmid minipreparations were performed essentially as described by Sambrook, et al. [30], the molecular weight markers were labeled with dCTP32 according to standard procedures. Probe synthesis The probes used in the Southern blot hybridization experiment were generated by PCR using the primers 1F and 4R for probe 1 and 1F-R6 (reverse, 5'CTGG CTAGTGACGTCGAT) for probe 2. In both PCR reactions the pBluescript plasmid carrying the CDH cDNA was used as the template. Plasmid pBS3.6 (described bellow) was used as the template and primers Pp6 (forward, 5'CTAGGCACTGCCG ATAG)- Pp7 (reverse, 5'GCACTACTTAC ATTGTG) for the synthesis of probe 3. Ten nanograms of each plasmid were subjected to 30 cycles of amplification by PCR under the same reaction conditions as described above. Probes were generated from gel-purified DNA fragments by random-primer dCTP32 labeling with the Rediprime kit (Amersham Life Science). Isolation of 5' upstream ATG region Ten micrograms of genomic DNA from P. chrysosporium were taken and digested with several restriction enzymes. The digestion products were separated by agarose gel electrophoresis, transferred to a nylon membrane (Biodyne A transfer membrane, Pall Biosupport membranes) and hybridized with probe 1 and with probe 2 in order to identify the zone corresponding to the promoter region. Between the two hybridizations the filter was stripped for the removal of the probe. Hybridization was performed in 6X SCC, 25% formamide, 5X Denhart's, 10% dextran sulphate and 0.1% SDS at 65 °C. The filter was washed twice in 2X SCC, 0.1% SDS and once in 0.2X SSC, 0.1% SDS at room temperature, following a wash with the last solution, for 15 min at 60 °C. DNA fragments of about 1.2 and 3.6 kb size ranges from the BamH I-EcoR V digestion were gel excised and the DNA recovered using the QIAEX II Agarose Gel Extraction kit. The gel-purified fragments were ligated into EcoR V-BamH I pBluescript KS and the ligation products were used to transform the E. coli DH5a strain. Approximately 450 and 530 transformant colonies were screened for plasmids containing the 1.2 or 3.6 kb fragments, respectively, of probe 1 [30]. Two plasmids containing the predicted 1.2 and 3.6 kb inserts were selected and named pBS1.2 and pBS3.6 respectively. In order to identify the insertion occurring in the genome, the same previously used membrane was stripped and re-hybridized with a 2 Kb fragment from pBS3.6 (probe 2), using the hybridization conditions described above. DNA sequence analysis Double-stranded sequencing of the pBS1.2, pBS3.6, pT7CDH inserts were performed by the dideoxynucleotide chain termination method [31] with Sequenase version 2.0 modified T7 DNA polymerase (United States Biochemical). Primers (Pharmacia) were used to walk progressively along both DNA strands. DNA and protein sequences were analyzed and compared with sequences from the GenBank database using the BLAST [32] or FASTA [33] sequence analysis program. The nucleotide 5' upstream region sequence data will appear in the EMBL Nucleotide Sequence Database under the accession number X97832. Results and Discussion Isolation of the CDH gene from P. chrysosporium K3 The CDH gene was isolated through PCR by using the primers located at 5' and 3' region of the CDH cDNA (see Experimental procedures). The gene sequence was similar to the previously published cDNA sequence of this fungus [25], and it corresponds exactly to the cDNA sequence reported by Li, et al. [26], but showing thirteen interruptions of the open reading frame by short introns. In order to isolate the 5´non-coding region, the genomic DNA was digested with several restriction enzymes and hybridized in a Southern blot (Figure 1B). We used two specific CDH probes: probe 1 covering all the CDH cDNA and probe 2 which spans 670 bp in the 5' region of the CDH cDNA (Figure 1A). The BamH I site is located at 614 bp in the genomic clone, within the fourth intron, therefore two fragments or a single fragment longer than 614 bp should be expected when the total DNA is digested to completion with BamH I and hybridized to probe 1 or 2, respectively, provided that only a single copy of the gene exists in this organism. However, in the Southern blot experiment three fragments of 1.5, 4.1 and 8.0 kb were observed when hybridization was carried out with probe 1 (Figure 1 BI, lane 3) of which, only the 1.5 and 4.1 kb fragments were detected with probe 2 (Figure 1 BII, lane 3). Two fragments of similar sizes were observed, using EcoR I (lane 4), EcoR V (lane 5) and Xba I (lane 6) restriction enzymes and both probes in the hybridization. Since the gene does not have internal cut sites for these enzymes, it seems probable that these fragments represent two copies of the CDH gene in P. chrysosporium K3. As shown by digestion with Sal I, which cuts the gene three times, four different size fragments (1.8, 2.5, 2.6 and 4.8 kb) were detected using probe 1 (Figure 1BI, lane 7), and an expected smaller fragment of 290 bp was observed after 48 hours of exposure. The 1.8 kb fragment appeared at double intensity, which together with the 2.5 and 4.8 kb fragments were detected after hybridizing with probe 2 (Figure 1BII, lane 7), suggesting that the two copies show no differences from the central to the 3' terminal part of the gene. Genomic DNA was subjected to double digestion with several pairs of restriction enzymes, and results of hybridization of the resulting fragments with both probes are shown in Figure 1B, lanes 8-11. This analysis clearly established that there were no differences between the two copies in the coding and 3' flanking regions, and it shows that all differences occur within the 5' flanking region. A difference of about 2.6 kb was observed using all the restriction digestions (see for example Figure 1BII, lanes 3, 7, 10, 11 and 12), except with EcoR I-Hind III digestion (lane 13). On the basis of these results, a restriction map of the CDH gene and the flanking regions was constructed (Figure 1A). In order to isolate the two 5' upstream regions the 1.2 and 3.6 kb fragments obtained from the BamH I-EcoR V digestions were gel extracted and subcloned in the pBluescript plasmid, the resulting plasmids were named pBS1.2 and pBS3.6, respectively. The sequence analysis shows that there were no differences in 650 bp of the 5' region of the coding region and in the first 425 bp upstream the ATG. However, an inserted sequence was detected only in the pBS3.6 plasmid, followed by the same nucleotide sequence in both plasmids. Figure 1A: . schematic diagram of the CDH gene (open box) with 5' and 3' flanking regions (line), introns (solid box) and restriction sites BamH I (B), EcoR I (EI), EcoR V (EV), Xba I (X), Pst I (P), Sal I (S) and Hind III (H). The triangle represents a 2.6 kb insertion. Labeled horizontal bars beneath the maps identify the regions that correspond to probes used in Southern blot hybridization.
Figure 1. B: Southern blot analysis of genomic DNA from P. chysosporium K3. Genomic DNA was digested with BamH I (lane 3), EcoR I (lane 4), EcoR V (lane 5), Xba I (lane 6), Sal I (lane 7), EcoR I-BamH I (lane 10), EcoR V-BamH I (lane 11), EcoR I-Hind III (lane 12), EcoR I-Pst I (lane13). In lane 1 is the undigested genomic DNA, in lane 14, 1 ng of the genomic gene isolated with primers 1F-4R (probe 1) as the positive control. The DNA was electrophoretically separated on an 0.8% agarose gel, transferred to a nylon membrane and hybridized to probe 1(I) or 2 (II). Lanes 1 and 15 show the dCTP32 labeled-lDNA digested with Hind III and Hind III-EcoR I, respectively. The same molecular weight markers were run in lanes 8 and 9 and their sizes are shown in kb on both sides. Recently, Li, et al. reported that cellobiose dehydrogenase from P. chrysosporium OGC101 is encoded by two allelic variants, named cdh-1 and cdh-2 [27]. The cDNA and genomic sequence reported by us coincided with the sequence of cdh-2, as well as the previous cDNA sequence reported by Li, et al., except the inserted sequence found in the 5' upstream region of the CDH gene in strain K3 [26]. Sequence analysis of the insertion region Sequence analysis of the two isolated 5' flanking regions of the CDH gene revealed a 2.6 kb insertion at a GT site in one of them, 425 bp upstream of the ATG codon (Figure 1A). The complete sequence of the insertion was determined (Figure 2). An element was identified that contains perfect inverted terminal repeats (ITRs) of 14 bp.
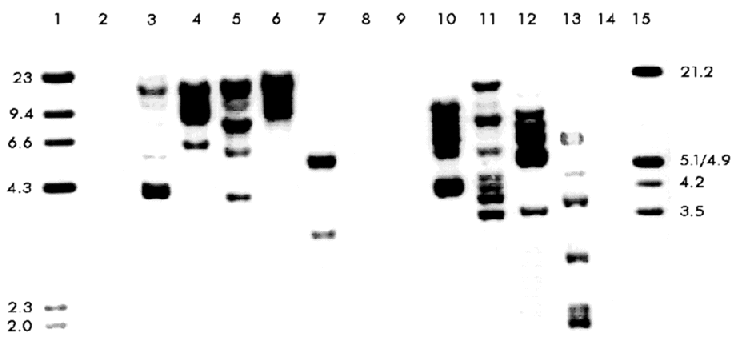
Figure 2. Nucleotide sequence of the insertion element (Pcc2) within the 5' upstream region of the plasmid pBS3.6. The underlined sequences are the 14 bp inverted terminal repeats (ITRs). The numbered sequence corresponds to the 5' flanking region of the CDH gene from the pBS1.2 plasmid. The nucleotide sequence has been deposited in the EMBL Nucleotide Sequence Database under the accession number X98835. The element was flanked on both sides by a GT dinucleotide, only one copy of which occurs at the corresponding site in the 5' upstream ATG region of the gene without the insertion, suggesting a duplication of the target site formed during the insertion. Certain features of the insertion sequence, such as the presence of perfect terminal repeats and the duplication of the target site associated with the insertion are common for many prokaryotic and eukaryotic transposons [11, 34]. The insertion therefore is likely to be a transposable element (which we named Pcc2), although this element does not contain a typical dinucleotide TA target duplication found in most of the fungal transposons related to the Caenorhabditis elegans Tc1 element [35]. Transposons have been found in a number of fungal species and classified in two main classes: class I elements which transposes by reverse transcription of an RNA copy of the element and class II which transposes directly through DNA copies. Class I includes elements with long terminal repeats (LTR) as Foret and skippy in F. oxysporum [12, 36], MAGGY in M. grisea [16], Boty in B. cinerea [17] and Mars4 in Ascobolus immersus [37]. Also several non-LTR retrotranposons with structural features of long interspersed nuclear elements have been characterized: Tad in N. crassa [13], MGR583 in M. grisea [15], Mars1 in A. immersus [37] and others, but short interspersed nuclear like elements have been recognized [10]. In class II some of the TEs are representative of families described in other organisms: impala [20] and Ant1 in A. niger [23]. Others are members of a new family described so far only in fungi (reviewed in 10), this is the case for Fot1 in F. oxysporum [18], Pot2 and MGR586 in M. grisea [21, 22]. Other less-characterized TEs are known that may represent new families: Hop in F. oxysporum [19] and Pce1 from P. chrysosporium [24]. Pcc2 has several features which allow to classify it as a transposon-like element, however with the data we now have, it is not possible to find a homology with either class 1 or class 2 of the described TEs families. We isolated a 2 kb fragment (probe 3) by PCR using specific primer oligonucleotides which correspond to the sequence of this element. With this probe and Southern hybridization, we found that this insertion appeared in high copy numbers and randomly dispersed within the genome of P. chrysosporium K3 (Figure 3), which is also a typical feature of a transposon-like element. Figure 3. Distribution of the Pcc2 element in the P. chrysosporium K3 strain. Southern blot is identical to that shown in Figure 1 B, but hybridized here with the Pcc2 probe (probe 3). A database examination and comparison of the Pcc2 sequence revealed no extended open reading frame and as in the first transposon-like element described in P. chrysosporium (Pce1) shows no significant homology with any known nucleotide or protein sequences. Other transposons contain large open reading frames which show similarity at the amino acid level to the transposase motif conserved in the Tc1 superfamily [10], but many others such as the Fot1 from F. oxysporum [18] and P. chrysosporium Pce1 [24] do not. The Pce1 element was shown to harbor a 1747 bp insertion within the lignin peroxidase allele of P. chrysosporium BKM-F-1767 and has several features of a transposon-like element, including inverted terminal repeats and a dinucleotide TA target duplication. Insertion of a transposable element within or adjacent to a chromosomal gene can directly alter its expression. In most cases the element blocks expression, but sometimes the effect is subtler. Transposable elements can increase the number of copies of sequences within the genome, either because they are, by chance, recognized as intermediates for replicative transposition events or because they are included in duplications produced by recombination between elements, generating sequence variation within genes that may be evolutionarily advantageous [10]. The presence of this element in the promoter region may affect the expression of the CDH gene and might play a role in genetic variation and evolutionary divergence of this gene in the P. chrysosporium genome. Further studies are underway to examine the possible role of Pcc2 on the inheritance and transcription of the CDH gene. Acknowledgement This work has been supported by the Swedish Agency for Research Cooperation with Developing Countries (SAREC). We thank Professor Ulf Pettersson and his staff at the Department of Medical Genetics of the Biomedical Center in Uppsala for giving BG the possibility to work in his laboratories. We deeply thank Professor Robert Brambl and Dr. David Eaker for valuable suggestions and critical reviews of the manuscript. References 1. Eriksson KE, Blanchette RA, Ander P. Microbial and enzymatic degradation of wood and wood components. Springer-Verlag Berlin Heidelberg, Germany; 1990. 2. Boominathan K, Reddy CA. Fungal degradation of lignin: Biotechnological Applications. Pp 763-822. In Arora, D.K., MuKerjee, K.G. and Elander, RP. (eds) Handbook of Applied Mycology. Vol. 4. Fungal Biotechnology. Marcel Dekker Inc. NewYork; 1992. 3. Kirk TK, Farrel RL. Enzymatic "combustion": the microbial degradation of lignin. Annu Rev Microbiol 1987;41:465-505. 4. Hammel KE. Organopollutant degradation by lignolytic fungi. Enzyme Microb Technol 1989;11:776-7. 5. Ruel K, Barnoud F, Eriksson KE. Ultrastructural aspect of wood degradation by Sporotrichum pulvurulentum. Observations on spruce wood impregnated with glucose. Holzforschung 1984;38:61-8. 6. Uzcategui E, Raices M, Montesino R, Johansson G, Pettersson G, Eriksson KE. Pilot scale production and purification of the cellulolityc enzyme system from the white rot fungus Phanerochaete chrysosporium. Biotechnol Appl Biochem 1991;13:323-34. 7. Morpeth F. Some properties of cellobiose oxidase from the white rot fungus Sporotrichum pulvurulentum. Biochemical J 1985;228:557-64. 8. Bao W. Renganathan V. Cellobiose dehydrogenase of Phanerochaete chrysosporium enhances crystalline cellulose degradation by cellulases. FEBS Lett 1992;302:77-80. 9. Ander P. The cellobiose oxiding enzymes CBQ and CBO as related to lignin and cellulose degradation a review. FEMS Microb Rev 1994;13:297-312. 10.Daboussi MJ. Fungal transposable elements: generators of diversity and genetic tools. J Genet 1996;75:325-39. 11. Finnegan DJ. Transposable elements: How non-LTR retrotransposons do it. Current Biol 1997;7:245-8. 12. Julien J, Poirier-Hamon S, Brygoo Y. Foret1, a reverse transcriptase-like sequence in the filamentous fungus Fusarium oxysporum. Nucleic Acids Res 1992;20:3933-7. 13. Kinsey JA, Helber J. Isolation of a transposable element from Neurospora crassa. Proc Natl Acad Sci USA 1989; 86:19:29-33. 14. Dobinson KF, Harris RE, Hamer JE. Grasshopper, a long terminal repeat (LTR) retroelement in the phytopathogenic fungus Magnapothe grisea. Mol Plant-Microbe Interact 1993;6:114-26. 15. Hamer JE, Farral L, Orbach MJ, Valent B, Chumley FG. Host species-specific conservation of a family of repeated DNA sequences in the genome of a fungal plant pathogen. Proc Natl Acad Sci USA 1989;86:9981-5. 16. Farman ML, Tosa Y, Nitta N, Leong SA. MAGGY, a retrotransposon in the genome of the rice blast fungus Magnapothe grisea. Mol Gen Genet 1996; 251:665-74. 17. Diolez A, Marches F, Fortini D, Brygoo Y. Boty, a long terminal repeat retroelement in the phytopathogenic fungus Botrytis cinerea. Appl Environ Microbiol 1995;61:103-8. 18. Daboussi MJ, Langin T, Brygoo Y. Fot1, a new family of fungal transposable elements. Mol Gen Genet 1992; 232:12-6. 19. Daboussi MJ, Langin T. Transposable elements in the fungal plant pathogen Fusarium oxysporum. Genetica 1994; 93:49-59. 20. Langin T, Capy P, Daboussi MJ. The transposable element impala, a fungal member of the Tc1-mariner superfamily. Mol Gen Genet 1995;246:19-28. 21. Kachroo P, Leong SA, Chattoo BB. Pot2, an inverted repeat transposon from the rice blast funfus Magnapothe grisea. Mol Gen Genet 1994;245:339-48. 22. Farman ML, Taura S, Leong SA. The Magnapothe grisea DNA fingerprinting probe MGR586 contains the 3' end of an inverted repeat transposon. Mol Gen Genet 1996;251:675-81. 23. Glayzer DC, Roberts IN, Archer DR, Oliver RP. The isolation of Ant1, a transposable element from Aspergillus niger. Mol Gen Genet 1995;249:432-438. 24. Gaskell J, Wymelenberg AV, Cullen D. Structure, inheritance, and transcriptional effects of Pce1, an insertional element within Phanerochaete chrysosporium lignin peroxidase gene lipI. Proc Natl Acad Sci USA 1995;92:7465-9. 25. Raices M, Paifer E, Cremata J, Montesino R, Stahlberg J, Dive C, et al. Cloning and characterization of a cDNA encoding a cellobiose dehydrogenase from the white rot fungus Phanerochaete chrysosporium. FEBS Lett 1995;369:233-8. 26. Li B, Srinivasa R, Nagall A, Renganathan V. Cloning of a cDNA encoding cellobiose dehydrogenase from Phanerochaete chrysosporium. Appl Envirom Microbiol 1996;62:1329-35. 27. Li B, Srinivasa R, Nagall A, Renganathan V. Cellobiose dehydrogenase from Phanerochaete chrysosporium is encoded by two allelic variants. Appl Envirom Microbiol 1997;63:796-9. 28. Johnsrud SC, Eriksson KE. Cross-breeding of selected and mutated homokaryotic strains of Phanerochaete chrysosporium K-3: New cellulase deficient strains with increased ability to degrade lignin. Appl Microbiol Biotechnol 1985;21:320-7. 29. Raeder U, Broda P. Rapid preparation of DNA from filamentous fungi. Lett Appl Microbiol 1985;1:17-20. 30. Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning. A Laboratory Manual. Cold Spring Harbor Laboratory Press, New York; 1989. 31. Sanger F, Nicklen S, Coulson AR. DNA sequencing with chain terminating inhibitors. Proc Natl Acad Sci USA 1977; 74:5463-7. 32. Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol 1990;215:403-10. 33. Pearson WR, Lipman DJ. Improved tools for biological sequence analysis. Proc Natl Acad Sci USA 1988;85:2444-48. 34. Finnegan DJ. Eukaryotic transposable elements and genome evolution. Trends Genet 1989;5:103-7. 35. Prasat SS, Harris LJ, Baillie DL, Rose AM. Evolutionarily conserved regions in Caenorhabditis transposable elements deduced by sequence comparison. Genome 1991;34:6-12. 36. Anaya N, Roncero MIG. skippy, a retrotransposon from fungal plant pathogen Fusarium oxysporum. Mol Gen Genet 1995; 249:637-47. 37. Goyon C, Rossignol JL, Faugeron G. Native DNA repeats and methylation in Ascobolus. Nucl Acids Res 1996;24:3348-56. Copyright 1998 Elfos ScientiaeThe following images related to this document are available:Line drawing images[ba98036c.gif] [ba98036d.gif] [ba98036a.gif] [ba98036b.gif] |
| |||||||||

{kind=link}
{kind=link}
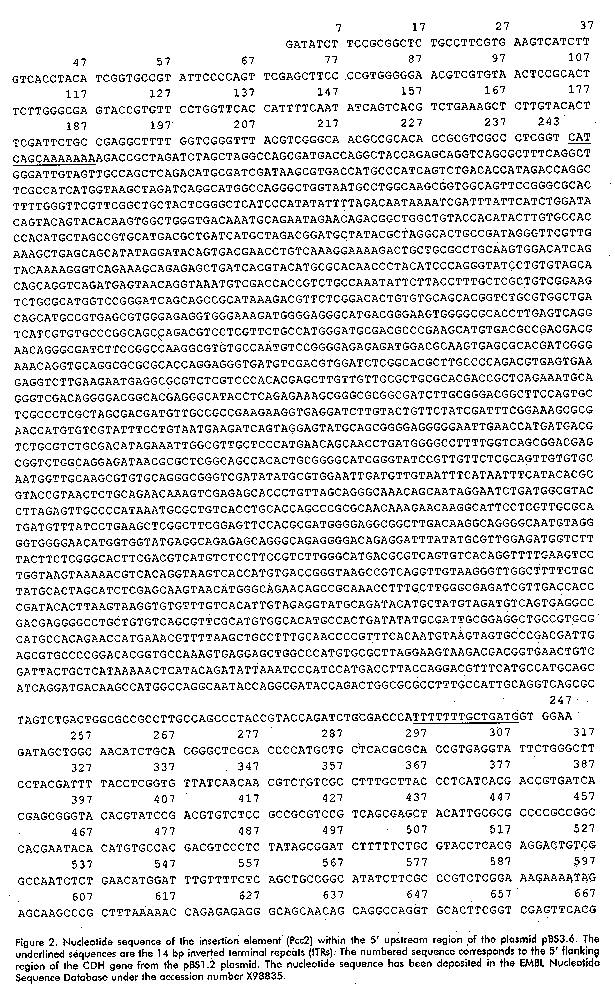{kind=link}
{kind=link}