 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 1998;15:232-236 HUMAN INTERFERON ALPHA 2B BEHAVES AS A TRIMER AT BASIC pH. A PRELIMINARY X-RAY DIFFRACTION ANALYSIS OF HUMAN ALPHA INTERFERON CRYSTALS GROWN AT BASIC pH @ Aida Díaz-Ruano, 1 Magdalena Segura-Nieto,2 Berenice Cuevas,2 Thierry Prangè31 Centro de Ingeniería Genética y Biotecnología PO Box 6162, Ciudad de La Habana, Cuba.Telf: (53-7) 21 8164, 21 8466; Fax: (53-7) 21 8070; E-mail: model@cigb.edu.cu 2Centro de Investigación y Estudios Avanzados del Instituto Politécnico Nacional de México. Unidad Irapuato, PO Box 629, 36 500, Irapuato. Gto., México. 3Laboratoire pour l Útilisation du Rayonnement Electromagnetique (LURE), Bâtiment 209-D, Centre Universitaire Paris Sud. 91405 Orsay Cedex. France.
Code Number: BA98037
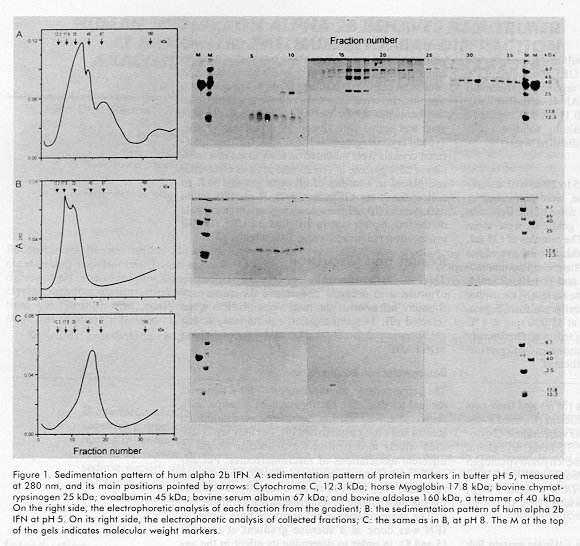
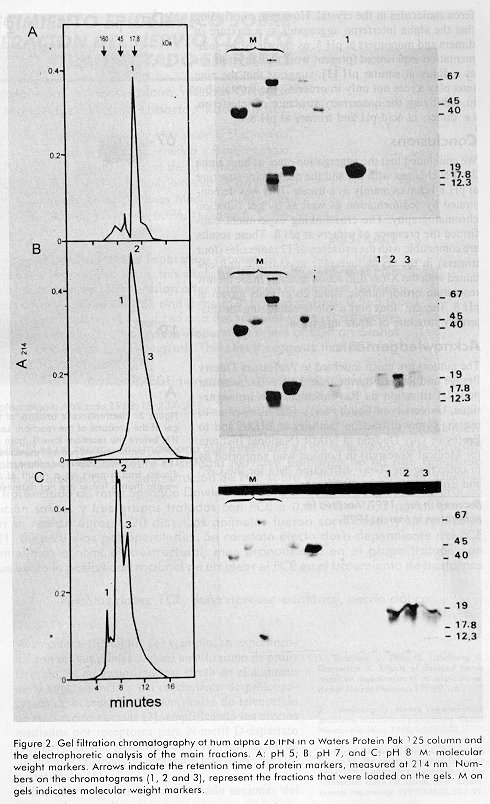
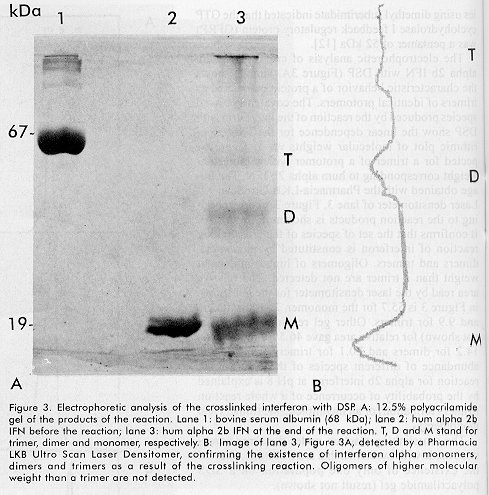
ABSTRACT Recombinant human alpha 2b interferon (hum alpha 2bIFN) was studied by sedimentation, gel filtration and crosslinking experiences. These experiences showed that the quaternary structure of hum alpha 2bIFN changes at different pH. A preliminary X-ray analysis of low resolution orthorhombic crystals of this molecule was done and the crystallographic asymmetric unit contains 12 molecules, possibly four trimers according to the previous sedimentation, gel filtration and crosslinking experiences. The role of the Zn2+ ions in the stabilization of the quaternary structure of alpha interferon is discussed. Keywords: crosslinking reagents, gel filtration, interferon, protein crystals, quaternary structure, sedimentation equilibrium, X-ray diffraction RESUMEN El interferón alfa 2b humano recombinante es estudiado por sedimentación, gel filtración y mediante experiencias con reactivos entrecruzantes. Estas experiencias muestran que la estructura cuaternaria de esta molécula varía con el pH. Se realiza un análisis preliminar por difracción de rayos X a baja resolución en cristales ortorrómbicos de esta molécula. La unidad asimétrica contiene 12 moléculas, posiblemente 4 trímeros, lo cual coincide con los resultados obtenidos por sedimentación, gel filtración y con los experimentos con los reactivos entrecruzantes previamente realizados. Se discute la influencia de los iones Zn2+ en la estabilización de la estructura cuaternaria del interferón alfa. Palabras claves: cristales de proteínas, difracción de rayos X, estructura cuaternaria, gel filtración, interferón, sedimentación reactivos entrecruzantesIntroduction Human alpha 2b interferon (hum alpha 2b IFN) is a 19 256 Da protein of 165 amino acids, belonging to the cytosine family. It is very active in conferring protection against a wide range of viruses [1,2]. After a considerable search for a well ordered crystal, the three-dimensional structure of hum alpha 2b IFN was determined at 2.9 Å resolution [3]. Hum alpha 2b IFN exists in this crystal as a noncovalent dimer, where the interactions between the monomers are mediated by a Zinc ion (Zn2+). These crystals are monoclinic and the crystallographic asymmetric unit contains six molecules. The calculated hydrodynamic volumes of this protein, determined by time resolved fluorescence suppose the existence of a dimer and a tetramer at pH5.5 and 9.4, respectively [4]. We have observed the same behavior during preliminary gel filtration studies. These preliminary results lead to other experiments to try to understand the aggregation-state of alpha interferon and its dependence with the pH. We describe in this report some experiences, at selected pH, confirming that the quaternary structure of the hum alpha 2b IFN changes with pH and it is a trimer at pH 8. We also report the preliminary x-ray diffraction analysis of low-resolution orthorhombic crystals of hum alpha 2b IFN. The results obtained for hum alpha 2b IFN by sedimentation equilibrium, gel filtration and crosslinking discussed in this paper help us to explain the x-ray diffraction results. Materials and Methods Characteristics of the human alpha interferon used in the present work The hum alpha 2b IFN was obtained by recombinant DNA technology and it was characterized by mass spectrometry [5,6]. The last purification step was done in an analytical reverse phase high performance liquid chromatography (rp-HPLC) C4 wide pore column (JT Baker). The solutions used were trifluoroacetic acid (TFA) (sequanal grade, Pierce) 0.1% in water (solution A), and TFA 0.05% in acetonitrile (HPLC grade Merck, Germany) (solution B). The hum alpha 2b IFN leaves the column at 56% solution B in a 60 min linear gradient from 15% solutionA to 60% solution B. The hum alpha 2b IFN from the rp-HPLC C4 column was diluted in water twice and dialyzed to ammonium acetate 50mM pH5 to eliminate the acetonitrile and to fold the interferon at the same time. SDS-PAGE analysis Analysis by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) containing 12% acrylamide was performed as reported by Laemmli [7]. Gels were run in PROTEAN II cells (BioRad), and stained in a solution of 0.1% w/w Coomassie blue R-250 (BioRad) in destaining solution (10% methanol-10% acetic acid in distilled water). Sedimentation equilibrium Linear sucrose gradients from 5 to 20%, with a cushion of 500mL of 60% sucrose, were performed by gravity in tubes in an SW40Ti rotor of the Beckman Ultracentrifuge L8-70M. The gradients were incubated 12h, at 4ºC before use. The samples of 125mL of protein at 4mg/mL were added at the top of the gradients. The buffer solutions were ammonium acetate-acetic acid 50mM, pH5 and Tris-HCl 25mM, pH8. The sucrose, ammonium acetate, Tris, acetic acid and the HCl were analytical grade from Sigma. Centrifugation was performed at 35 000 rpm at 4ºC for 50 h. Gradients were collected in approximately 35 fractions of 350 mL each, at the end of centrifugation. Each gradient was duplicated. Molecular weight markers The molecular weight markers (MW) and their corresponding sedimentation coefficient (S), used in the SDS-PAGE and in the gradients were: rabbit C cytochrome (MW 12.3kDa, 1.71 S), horse myoglobin (MW 17.8 kDa, 2.0 S), bovine chymotrypsinogen (MW 25 kDa, 2.54 S), ovoalbumin (MW 45 kDa, 3.45S), bovine serum albumin (MW 67kDa, 4.06S) and bovine aldolase (MW 160 kDa, 7.35 S). Gel filtration analysis Gel filtration was performed in a Waters protein Pak 125 column, in a Waters HPLC. The gel filtration runs were at pH 5, 7 and 8. Molecular weight markers and interferon samples were in three pH eluents: 25-mM ammonium acetate, pH 5; 25 mM Tris, pH 7 and 25mM Tris, pH 8.0. Crosslinking experiences The crosslinking reagent was dithio bis succinyl propionate (DSP) from Pierce. It is a homobifunctional N-hydroxisuccinimide (NHS) ester with a spacer arm of 12 Å. The reaction of the hum alpha 2b IFN with DSP took place in 50 mM triethanolamine buffer pH 8, 100 mM NaCl for 60 min at room temperature. DSP was previously dissolved in dimethylsulfoxide (DMSO). The molar amount of DSP to the total amount of lysine was 100 (hum alpha IFN has 10 Lys). The concentration of hum alpha 2b IFN was 1mg/mL at the beginning of the reaction. The retention time of the electrophoretic bands corresponding to the products of the crosslinking reaction was determined with a Pharmacia-LKB Ultro-Scan XL Laser densitometer. X-ray diffraction analysis Orthorhombic hum alpha 2b IFN crystals were grown at 20ºC using hanging drops equilibrated by vapor diffusion. Typically, 2.5 mL of the protein at a concentration of 25mg/mL in 25 mM Tris pH 8 buffer was combined on a siliconized cover slip with an equal volume of a reservoir solution containing 2M potassium phosphate buffered at pH8. The cover slip was then inverted over 1 mL of the reservoir solution contained in the well of a Limbro cell culture plate and sealed with silicon oil or vacuum grease. Typical crystals were of 0.5X0.25X0.1mm. Interferon crystals were mounted on fiber loops in a solution of 25% glucose in Tris 25 mM pH8 as cryobuffer and placed in a stream of nitrogen gas at 100K to collect the X-ray diffraction data. X-ray diffraction data were collected from randomly oriented crystals on the image plate system installed at LURE (DW32 synchrotron radiation station, DCI wiggler [8]). Results and Discussion Three methods were used to monitor the association of proteins in order to understand the quaternary structure behavior of the hum alpha 2b IFN at the selected pH: 1- sedimentation equilibrium, 2- fast gel filtration chromatography, and 3- crosslinking SDS-PAGE. Sedimentation equilibrium The sedimentation equilibrium provides a powerful method for detecting heterogeneity of a sample with respect to self-association. Sedimentation equilibrium studies were long established [9], but they are is still used widely to determine the molecular weight of different associations of macromolecules. For example, sedimentation analyses of R67 dihidrofolate reductase [10], have been done recently to monitor the pH dependence of its oligomerization state. The sedimentation equilibrium of hum alpha 2b IFN was done in a sucrose gradient at two pH (5and8), in order to determine its effect in the aggregation-state of the protein. The sucrose gradients were simultaneously calibrated with marker proteins of known molecular weights and sedimentation coefficient at pH 5 and 8. There was almost no difference in the sedimentation behavior of the markers at both pH (data not shown). In Figure 1A, the sedimentation pattern of the marker proteins at pH 5 is shown and the corresponding electrophoretic analysis of each fraction is also given at the right side of this figure. Figure 1. Sedimentation pattern of hum alpha 2b IFN. A: sedimentation pattern of protein markers in buffer pH 5, measured at 280 nm, and its main positions pointed by arrows: Cytochrome C, 12.3 kDa; horse Myoglobin 17.8 kDa; bovine chymotrypsinogen 25 kDa; ovoalbumin 45 kDa; bovine serum albumin 67 kDa; and bovine aldolase 160 kDa, a tetramer of 40kDa. On the right side, the electrophoretic analysis of each fraction from the gradient; B: the sedimentation pattern of hum alpha 2b IFN at pH 5. On its right side, the electrophoretic analysis of collected fractions; C: the same as in B, at pH 8. The M at the top of the gels indicates molecular weight markers. The sedimentation pattern of hum alpha 2b IFN made at pH5 and the electrophoretic analysis of each fraction (Figure 1B), reveal that the interferon spread from fraction 5 to 11. This is explained because there is a mixture corresponding to molecules of heterogeneous size, of monomers and dimers, with a maximum in the monomer zone. On the other hand, the results of the sedimentation pattern of hum alpha 2b IFN ob-tained at pH8 (Figure 1C) show that interferon appears mainly on fraction 14 and 15 corresponding to a molecular weight of a trimer. There was a clear difference in the sedimentation pattern of hum alpha 2b IFN in the two pH, suggesting differences in self-association. At pH5 there is a predominance of monomers and dimers (Figure 1B), while at pH 8 there was a main component in the region of trimers (Figure 1C). Gel filtration analysis Fast gel-permeation chromatography also allows a fast direct measurement of the size of pure proteins, according to its state of association. The gel filtration chromatographic profiles of hum alpha 2b IFN were different according to pH. At pH5, hum alpha 2b IFN elutes later in the region of a monomer (Figure 2A). The electrophoretic analysis of the top fraction of peak 1, in lane 1 (right side of panel 2A), contains most of the protein loaded onto the column. Hum alpha 2b IFN elutes at the retention time of a dimer (i.e. 38.5 kDa) at neutral pH (Figure2B). At pH7, the peak is wider, and probably contains reversible self-association of monomers and dimers, with the predominance of dimers as compared with the retention time of the molecular weight markers. Hum alpha 2b IFN elutes mainly as a trimer (i.e. 57.8kDa) at pH 8 (peak 2, Figure 2C). This result confirms the first observation with the sedimentation experiment (Figure 1C), that at basic pH, hum alpha 2b IFN behaves mainly as a trimer. Peak 1 should correspond to oligomers of a size greater than a trimer. It represents the exclusion volume that is 80kDa for globular proteins. Peak 3 represents the dimer population (38.5 kDa), according to its retention time. The electrophoretic analysis of fractions 1 to 3 from the chromatogram is shown in Figure 2C. Lane2 is loaded with the main component, peak 2. This peak has two components. The first one corresponds to the trimer (57.8 kDa) and the second one is probably the equilibrium mixture of trimers and dimers. Peak 3 was loaded on lane 3. The fractions of hum alpha 2b IFN in buffers with pH7 and 8 were dialyzed after gel filtration, lyophilized and applied into the gel. This manipulation could be the reason why the proteins show some degradation in the gels (panel 2B and 2C, lanes 1-3, both gels, at the right side of Figure 2). Figure 2. Gel filtration chromatography of hum alpha 2b IFN in a Waters Protein Pak 125 column and the electrophoretic analysis of the main fractions. A: pH 5; B: pH 7, and C: pH8. M: molecular weight markers. Arrows indicate the retention time of protein markers, measured at 214 nm. Numbers on the chromatograms (1, 2 and 3), represent the fractions that were loaded on the gels. M on gels indicates molecular weight markers. Crosslinking analysis Chemical crosslinking analysis with bifunctional reagents allows a "snap-shot" fixation of reassociating oligomers. The early studies with dimetyl suberimidate (DMS) [11] showed that the products of the crosslinking reaction are constituted by a set of species whose molecular weights are equal to an integral number of the protomer molecular weight. For oligomers composed of identical protomers the number of products is equivalent to the number of protomers in the oligomer. As an example, recent cross-linking studies using dimethyl suberimidate indicated that the GTP cyclohydrolase I feedback regulatory protein (GFRP) was a pentamer of 52 kDa [12]. The electrophoretic analysis of crosslinked hum alpha 2b IFN with DSP (Figure 3A, lane 3) shows the characteristic behavior of a protein associated as trimers of identical protomers. The covalently linked species produced by the reaction of the interferon with DSP show the linear dependence for the semi-logarithmic plot of molecular weights vs. distance expected for a trimer of a protomer with a molecular weight corresponding to hum alpha 2b IFN. The image obtained with the Pharmacia-LKB UtroScan XL Laser densitometer of lane 3, Figure 3A corresponding to the reaction products is shown on Figure3B. It confirms that the set of species of the crosslinking reaction of interferon is constituted by monomers, dimers and trimers. Oligomers of higher molecular weight than a trimer are not detected. The relative area read by the laser densitometer for the gel shown in Figure 3 is 55.7 for the monomer, 25.8 for dimers and 9.9 for trimers. Other gel readings (this gel is not shown) for relative area gave 46.5 for monomers, 14.2 for dimers and 20.1 for trimers. The relative abundance of different species of the crosslinking reaction for alpha 2b interferon at pH8 is explained by the probability of occurrence of a whole reaction (trimers formation) or an incomplete one (dimers and monomers). Figure 3. Electrophoretic analysis of the crosslinked interferon with DSP. A: 12.5% polyacrilamide gel of the products of the reaction. Lane 1: bovine serum albumin (68kDa); lane 2: hum alpha 2b IFN before the reaction; lane 3: hum alpha 2b IFN at the end of the reaction. T, D and M stand for trimer, dimer and monomer, respectively. B:Image of lane 3, Figure 3A, detected by a Pharmacia LKB Ultro Scan Laser Densitomer, confirming the existence of interferon alpha monomers, dimers and trimers as a result of the crosslinking reaction. Oligomers of higher molecular weight than a trimer are not detected. The crosslinking reaction under the same conditions made with Lysozime, that is a monomer, showed the existence of only one population on a SDS polyacrilamide gel (result not shown). X-ray diffraction analysis Crystals of hum alpha 2b IFN were grown at pH 8, using potassium phosphate as the precipitating agent. They were analyzed by x-ray diffraction. Bragg reflections were observed to a resolution not better than 7 Å for these crystals. Cell parameters were determined using the IMREFIX and IDXREF Programs in the MOSFLM package [13]. The space group was determined by analysis of systematic extinction and assigned as orthorhombic P21 21 21. Unit cell parameters of the resulting cell are: a=93.3; b=121.4 and c=224.1 Å. The relation of the volume of the asymmetric unit to the protein molecular mass for this crystal considering 50% of solvent contented, suggests that the crystallographic asymmetric unit contains 12 molecules. This is compatible with the existence of 4 trimers according to the previous results. The knowledge of the existence of interferon trimers in solution at pH 8 and its possibility to associate as a dodecamer in the crystal explains the unexpected large size of the unit cell of this crystal. Comparing the dimension of the asymmetric unit and the symmetry of these crystals with the ones reported by Rradhakrishnan [3], it is obvious that the self-aggregation states of the interferon are different. This result is explained mainly by the different pH used to grow both crystals. Trying to improve the quality of the phosphate interferon alpha crystals, we performed some crystallization trials adding divalent cations [14]. We obtained crystals using pH 8 and phosphate as the precipitating agent but introducing up to 0.5 mM of Cadmium (Cd2+) and Zinc (Zn2+) ions in the mother liquid where the crystals grew. These crystals have the same orthorhombic symmetry and about the same size of the asymmetric unit as the phosphate crystals. We improved the resolution of the orthorhombic crystals up to 4.5 Å with the crystals containing Zn2+ while those without a divalent cation and others with Cd2+, diffracted only up to a 7Å resolution under the same measuring conditions. This result suggests that Zn2+ ions play a role in ordering the interferon molecules in the crystal. However, our finding that the alpha interferon aggregates as a mixture of dimers and monomers at pH5, as shown by the sedimentation experiment (present work) but crystallizes as a dimer at similar pH [3], suggests that the zinc ions play a role not only in ordering the crystals but in stabilizing the quaternary structure of interferon, i.e. dimers at acid pH and trimers at pH 8. Conclusions We concluded that the aggregation-state of hum alpha 2b IFN changes with pH and the quaternary structure at pH 8 behaves mainly as a trimer. This was demonstrated by sedimentation as well as by gel filtration chromatography. The crosslinking experiment confirmed the presence of trimers at pH8. These results are compatible with the existence of 12 molecules (four trimers) in the crystallographic asymmetric unit obtained with the X-ray diffraction analysis made on low resolution orthorhombic alpha 2b crystals grown at pH8. The Zn2+ ions play a role in stabilizing the quaternary structure of alpha interferon. Acknowledgements The authors are much indebted to Professors Thierry Prange and Roger Fourme from LURE (Laboratoire pour l Útilisation du Rayonnement Electromagnetique, University of South Paris), who allowed us to use the X-ray diffraction facilities at LURE and to Professor Guy Dodson at NIMR (National Institute for Medical Research) in London who supported us in collecting the X-ray diffraction data for the Cd2+ and Zn2+ interferon crystals. R eferences1. Isaacs A, Lindenman J. Virus interference 1. The interferon. Proc R Soc London Ser B 1957;147:258-67. 2. De Maeyer E, de Maeyer-Grignar J. Interferons and other related cytokines. John Wiley and Sons, New York .1989. 3. Rradhakrishnan R, Walter LJ, Hruza A, Reichert P, Trotta PP, Nagabhushan TL, etal. Zinc mediated dimer of human interferon-µ2b revealed by x-ray crystallography. Structure 1996;4:1453-1463. 4. Vicent M, Li de la Sierra I, Berberán Santos M, Díaz A, Díaz M, Padrón G, etal. Time-resolved flluorescense study of human recombinant interferon µ2. J Eur J Biochem 1992;210:953-961. 5. Quiñones Y, Agraz A, Silva A, Padrón G, Mella C, Díaz R, et al. High purity recombinant human alpha-2 interferon free from oligomeric forms in E. coli. Highlights Mod Biochem 1992;2:1237-1246. 6. Padrón G, Besada V, Agraz A, Quiñones Y, Herrera L. Mass spectrometric analysis of recombinant human µ-2 interferon. Analytica Chimica Acta 1989; 223:361-9. 7. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1971;277:680-5. 8. Fourme R, Dhez P, Benoit JP, Kahn R, Dubuisson JM, Besson P, et al. Bent crystals, bent multilayer optics on a multipole wiggler line for x-ray diffractometer with an image plate detector. Rev Sci Instrum 1992;63:810-3. 9. Fruton JS, Simmonds S. General Biochemistry. E.R. 2nd. ed., La Habana, 1958. 10. Bradrick TD, Shattuck C, Strader MB, Wickers C, Eiseintein C, Howell EE. Redesigning the quaternary structure of R67 dihydrofolate reductase. J Biol Chem 1996;271:28031-7. 11. Davies GE, Stark GR. Use of dimethyl suberimidate: a crosslinking reagent in studying the subunit structure of oligomeric proteins. Proc Natl Acad Sci USA 1970;66:651. 12. Yoneyama T, Brewers JM, Hatakeyama K. GTP cyclohydrolase I feedback regulatory protein is a pentamer of identical subunits. J Biol Chem 1997; 272:9690-6. 13. Leslie AGW. MOSFLM. A program package for processing rotation data collected on film or image plates. Distributed from AGW. Leslie, MRC Laboratory of Molecular Biology. Hills Road, Cambridge, CB2 2QH, U.K. 1992. 14.Trakhanov S, Quiocho FA. Influence of divalent cations in protein crystallization. Protein Science 1995;4:1914-19. Copyright 1998 Elfos Scientiae The following images related to this document are available:Photo images[ba98037b.jpg] [ba98037c.jpg] [ba98037a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}