 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnologia Aplicada 1998;15:237-241 EFECTO DEL FACTOR DE CRECIMIENTO EPIDÉRMICO SOBRE LA REGENERACIÓN DEL NERVIO CIÁTICO TRANSECTADO EN RATAS @ Pedro A Prats,1 Lázaro O Castañeda,1 Viviana Falcón,2 Rafael Ortega,3 María C de la Rosa,2 Ivón Menéndez,2 Violeta Labarta,4 Raúl Gómez21 Departamento de Estudios Preclínicos, 2Departamento de Microscopía Electrónica,4División de Estudios Clínicos, Centro de Ingeniería Genética y Biotecnología. AP 6162, CP 10600, Ciudad de La Habana, Cuba. Fax: (53-7) 21 8070; E-mail: bioensa@cigb.edu.cu 3Departamento de Cirugía Experimental, Centro de Investigaciones Médico-Quirúrgicas. Calle 216 y 11-B, Siboney, Playa, AP 6096, Habana 6, Cuba.
Code Number: BA98038
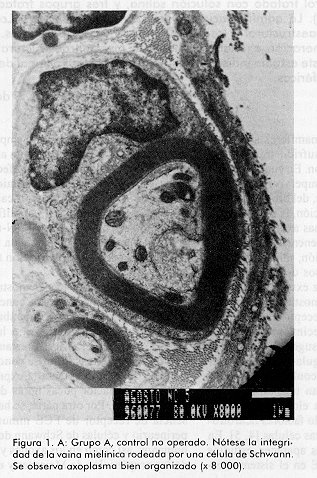
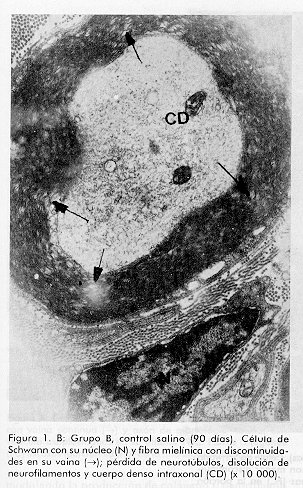
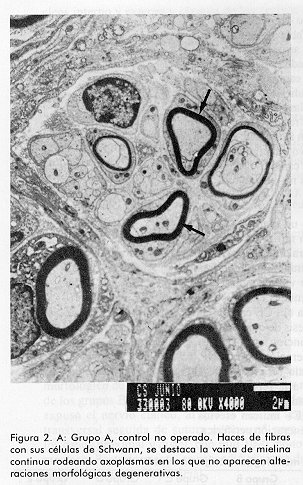
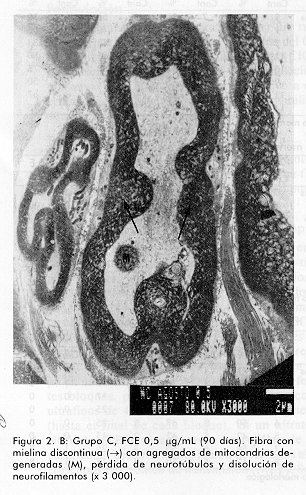
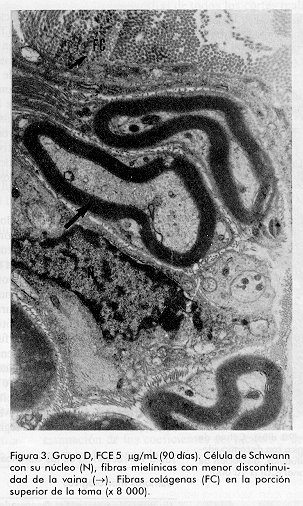
ABSTRACT Epidermal growth factor (EGF) is a well known mitogen involved in the reparation process of a variety of tissues, however, no action on peripheral nerve system has been reported so far. In this study we evaluate the effects of EGF administration on some ultrastructural indicators of sciatic nerve regeneration after transection in Sprague Dawley rats. The design included five groups (non operated control, saline control, and three EGF treated groups: 0.5; 5.0, and 10mg/mL). The administration lasted 20 days. The animals were sacrificed and the samples were ultrastructurally evaluated at days 21, 60 and 90 post-lesion. It was demonstrated a dose-dependent effect of EGF on axonal regeneration levels, more pronounced in the group treated with EGF 10 mg/mL. This result suggests that EGF is a potential candidate for the treatment of peripheral nerve disorders. Keywords: EGF, peripheral nerve injury, sciatic nerve RESUMEN El factor de crecimiento epidérmico (FCE) es un reconocido mitógeno que interviene en los fenómenos de reparación de gran variedad de tejidos, no obstante, su acción sobre el sistema nervioso periférico no ha sido reportada todavía. En este estudio se investigan los efectos de la administración de FCE sobre indicadores ultraestructurales de regeneración del nervio ciático transectado de ratas Sprague Dawley. El diseño incluyó cinco grupos (control no operado, control tratado con solución salina, y tres grupos tratados con FCE a 0,5; 5,0 y 10,0mg/mL por vía intraperitoneal). La administración se realizó durante 20 días. Los animales fueron sacrificados y se realizó la evaluación ultraestructural a los 21, 60 y 90 días postoperatorios. Se constató efecto dosis dependiente del FCE sobre la regeneración axonal y mielínica a nivel ultraestructural, más pronunciado en el grupo tratado con 10,0mg/mL. Este estudio indica que existe la posibilidad racional de emplear el FCE en el tratamiento de trastornos de nervios periféricos. Palabras claves: FCE, daño nervioso periférico, nervio ciático Introducción Los nervios periféricos de los mamíferos son capaces de regenerar cuando han sufrido transección, aplastamiento u otro tipo de lesión. En cualquier caso la regeneración depende de la supervivencia de los cuerpos celulares de las neuronas, del brote de axones del cabo proximal y de la elongación, la reenvoltura y la maduración de las nuevas ramas axonales. El proceso reparativo posterior a la degeneración walleriana implica fenómenos de inflamación, migración celular y mitosis, además de procesos de síntesis y expresión de proteínas de la matriz extracelular [1, 2]. Experimentos anteriores han demostrado que tras un daño periférico los niveles de receptores para algunas citoquinas y factores de crecimiento aumentan significativamente; de ahí se ha sugerido que el proceso regenerativo puede estar regulado por estos factores peptídicos. Es un hecho confirmado que las células de Schwann desempeñan una función importante en la promoción de la elongación axonal [3] y aun más, se han identificado factores neurotróficos que son producidos por estas células [4, 5]. En la literatura de los últimos años aparecen estudios que relacionan el papel del FCE en el sistema nervioso central (SNC) [6] por ejemplo, en experimentos con células gliales, induce un alto ritmo de proliferación, se ha demostrado su efecto en el aumento de la supervivencia y del crecimiento de prolongaciones de neuronas subneocorticales de telencéfalo de ratas recién nacidas [7], amplificando los efectos mediados por receptores para N-metil D-aspartato (NMDA), los que participan en la reorganización de los campos cutáneos de recepción tras lesión nerviosa periférica [8]. Estudios recientes han demostrado acciones del FCE en la consolidación de conexiones sinápticas durante el desarrollo [9]. Sin embargo, no se ha encontrado información relativa a la posible función de este factor en el proceso restaurador de lesiones nerviosas periféricas. Sólo se conoce que en las células de Schwann aumenta la expresión de receptores para este factor pocas horas después de ocurrir daño nervioso. Por otra parte, se ha reportado la presencia de receptor de FCE inmunorreactivo en el perineurio y células de Schwann de nervios cutáneos humanos, así como en los axones y células lamelares de los corpúsculos de Meissner y en los axones, núcleos interno y externo y cápsula de los corpúsculos de Paccini [10]. El presente trabajo se diseñó para evaluar, a nivel ultraestructural, los efectos de la administración de FCE sobre el nervio ciático transectado en ratas. Materiales y Métodos Se utilizaron 90 ratas machos, de la línea Sprague Dawley, entre nueve y once semanas de edad y peso promedio entre 200 y 250g. Las ratas se dividieron en cinco grupos de 18 miembros cada uno y fueron anestesiadas con 40mg de pentobarbital sódico intraperitoneal por kilogramo de peso corporal y luego se rasuró el muslo derecho abarcando el área del nervio ciático. Los grupos experimentales fueron los siguientes: A Control normal (no operado) Procedimiento quirúrgico El grupo A sirvió sólo como referencia al aspecto morfológico de los nervios normales. Todas las ratas de los grupos B al E sufrieron el mismo proceder: se expuso el nervio ciático, al que se realizó sección transversal seguida de sutura perineural para fijar los cabos proximal y distal, con hilo 10-0 monofilamentoso. Posteriormente, se procedió al cierre de los planos muscular y superficial. Formulación El FCE humano recombinante (FCE-hr) fue suministrado por la Subdirección de Producción del Centro de Ingeniería Genética y Biotecnología (CIGB), La Habana, Cuba. Se elaboraron tres formulaciones utilizando el FCE-hr como principio activo a concentraciones de 0,5; 5 y 10mg de FCE-hr por mililitro de solución salina. Tratamiento A los animales de los grupos B-E se les administró, con frecuencia diaria, 1 mL de solución salina fisiológica o de la dosis correspondiente de FCE por vía intraperitoneal por espacio de veinte días. Toma de muestras y evaluación microscópica Se tomaron muestras del nervio ciático 2mm en dirección distal a la lesión, en cada uno de los tiempos. Los fragmentos de nervio fueron fijados en glutaraldehído al 3,2%, por 1 h a 4°C, lavados en tampón cacodilato 0,1 M, pH 7,4 y posteriormente postfijados en tetróxido de osmio por 1h a 4°C. Se deshidrataron en alcoholes de concentraciones crecientes y se incluyeron posteriormente en resina Spurr. Se colocaron todas las biopsias de cada grupo en un mismo frasco, y se incluyeron en diferentes bloques, posteriormente se realizaron cortes ultrafinos de 400 a 500 Å a todos los fragmentos (hasta el final de cada bloque), en un ultratome NOVA LKB y se colocaron en rejillas de 400 cuadrículas, posteriormente fueron observados al microscopio electrónico de transmisión (MET). Se tomaron microfotografías de todos los cortes realizados y de éstas se seleccionaron aleatoriamente doce vistas por grupo en cada tiempo, en las cuales se evaluó la frecuencia de las variables morfológicas examinadas. Las muestras para el análisis por MET fueron examinadas a ciegas. Indicadores evaluados. Se determinó la frecuencia de aparición de los siguientes cambios morfológicos en cada grupo, en los tres tiempos examinados: · Agregados de mitocondrias degeneradas· Cuerpos laminados intraxonales · Dilatación del retículo endoplasmático liso (vesículas axoplasmáticas) · Cuerpos densos intraxonales · Pérdida de neurotúbulos · Disolución de neurofilamentos · Incremento de fibras colágenas · Discontinuidad de la vaina de mielina Análisis estadístico Para la evaluación de los parámetros morfológicos en cada uno de los tiempos, se aplicó el Test Chi-cuadrado cuando la naturaleza de los datos lo permitió y la prueba exacta de Fisher en los casos en que se detectó diferencias. Con el objetivo de estudiar cuánto más frecuente era la aparición de cambios morfológicos en cada grupo de tratamiento con respecto al control, se realizó un análisis de regresión logística con cada cambio. Se procedió a la estimación de los coeficientes del modelo y de las razones de disparidad (odds ratios). En los casos en que los datos no se explicaban por el modelo se estimaron las diferencias de proporciones y los riesgos relativos, medidas que dan la razón de probabilidad entre las proporciones de cada grupo con respecto al control. Resultados Como se esperaba, el grupo B (control salino) fue el que presentó peor evolución y mayor número e intensidad de cambios morfológicos respecto al grupo A (nervios sanos) (Figura1). A los 21 días no se encontró diferencias entre los grupos en cuanto a la frecuencia de los cambios morfológicos evaluados, sólo en el caso de los cuerpos densos intraxonales se pudo verificar mejoría palpable en el grupo E (10mg/mL) en relación con el B (control salino) (Tabla1). En la evaluación intermedia (60 días) se encontraron diferencias significativas según el Test Chi-cuadrado para las variables cuerpos densos intraxonales (X2=9,058; p=0,028) y discontinuidad de la vaina de mielina (X2=12,420; p=6,075E-03) en relación con el control B. Con la aplicación de la prueba exacta de Fisher se encontraron diferencias significativas para varios cambios morfológicos al comparar cada grupo de tratamiento con el control. Entre los grupos B (control salino) y C (0,5mg/mL) no se obtuvieron diferencias estadísticamente significativas para las variables analizadas (Tabla2). Figura 1. A: Grupo A, control no operado. Nótese la integridad de la vaina mielínica rodeada por una célula de Schwann. Se observa axoplasma bien organizado (x 8 000). Figura 1. B: Grupo B, control salino (90 días). Célula de Schwann con su núcleo (N) y fibra mielínica con discontinuidades en su vaina (®); pérdida de neurotúbulos, disolución de neurofilamentos y cuerpo denso intraxonal (CD) (x 10 000). Figura 2. A: Grupo A, control no operado. Haces de fibras con sus células de Schwann, se destaca la vaina de mielina continua rodeando axoplasmas en los que no aparecen alteraciones morfológicas degenerativas. Figura 2. B: Grupo C, FCE 0,5mg/mL (90 días). Fibra con mielina discontinua (®) con agregados de mitocondrias degeneradas (M), pérdida de neurotúbulos y disolución de neurofilamentos (x 3 000). Figura 3. Grupo D, FCE 5mg/mL (90 días). Célula de Schwann con su núcleo (N), fibras mielínicas con menor discontinuidad de la vaina (®). Fibras colágenas (FC) en la porción superior de la toma (x 8 000). Figura 4. Grupo E, FCE 10 mg/mL (90 días). Fibra mielínica con vaina continua (®) y axón de morfología normal con vesícula axoplasmática en su interior (VA). Fibras colágenas (FC) escasas (porción inferior de la microfotografía) (x 15 000). A los 90 días se encontraron diferencias significativas entre los grupos tratados y el control B en relación con todos los cambios morfológicos evaluados; de nuevo el grupo E (10mg/mL) desplegó el mejor desenlace seguido por los grupos D (5mg/mL) y C (0,5mg/mL) en ese orden, semejando un efecto dosis dependiente sobre el proceso de regeneración (Tabla3; Figuras1-4). El análisis de regresión logística arrojó siempre una mayor tendencia de los grupos tratados con FCE a presentar menores probabilidades de cambios morfológicos, también de modo dosis dependiente. En el último momento evaluado la mayoría de las variables presentaron valores tan pequeños de odds ratios y de probabilidad de cambio que no se ajustaron al modelo; no obstante, los datos de riesgo relativo favorecieron las probabilidades de no ocurrencia de cambios morfológicos en los animales tratados (datos no mostrados). Tabla 1. Frecuencia de los cambios morfológicos por grupo a los 21 días.
Cant: frecuencia de aparición del cambio morfológico. Tabla 2. Frecuencia de los cambios morfológicos por grupo a los 60 días.
Cant: frecuencia de aparición del cambio morfológico Tabla 3. Frecuencia de los cambios morfológicos por grupo a los 90 días.
Cant: frecuencia de aparición del cambio morfológico Discusión En este estudio hemos empleado un modelo in vivo de lesión nerviosa periférica, con miras a evaluar la recuperación a nivel ultraestructural de las fibras nerviosas transectadas, sometidas a tratamiento con tres dosis de FCE-hr administrado por vía sistémica. Partiendo de los hallazgos experimentales en el SNC y de la presencia de receptores para el FCE en células de Schwann, se concibió la hipótesis de que este factor podría desempeñar alguna función en los fenómenos que conllevan a la regeneración en el SNP. Nuestros resultados apuntan hacia un efecto dosis dependiente del FCE en cuanto a la reparación de las fibras nerviosas, evidenciado en el análisis ultraestructural. Los datos de MET indican efectos beneficiosos del FCE sobre la regeneración periférica, previniendo la formación de neuromas. Resulta razonable plantear que el tratamiento con FCE protegió a los cuerpos neuronales de los severos cambios de la cromatolisis y posibilitó la deposición de una matriz extracelular mejor organizada. El colágeno en el compartimiento endoneural normal está altamente organizado y forma dos capas distintas sobre las fibras nerviosas mielínicas y no mielínicas. La orientación longitudinal de las fibras de colágeno en la capa endoneural externa junto con la membrana basal de la célula de Schwann juegan un rol importante en la orientación de los axones que regeneran luego de lesión periférica. En la medida que las células de Schwann proliferan forman columnas (bandas de Büngner) rodeadas por la membrana basal [11]. Puesto que estas células expresan receptores para el FCE, cabría pensar en una acción aceleradora del péptido sobre la proliferación de las células de Schwann, y de esta manera acelerar la preparación del camino por donde habrán de crecer los axones regenerantes. En tejidos como el epitelio corneal se ha demostrado inducción de la síntesis de fibronectina por el FCE [12, 13], esta glicoproteína junto a la laminina y el colágeno IV son componentes de la matriz extracelular de capital importancia en los eventos de regeneración de los nervios periféricos [14, 15]. Una de las acciones del FCE podría consistir en el incremento de la producción de este u otros elementos que contribuyan directamente a la restauración axonal. Es plausible, incluso, la idea de que el FCE actúe sobre los fibroblastos endoneurales modulando la producción de colágeno y controlando su organización en la etapa de remodelación, a juzgar por la disminución en las fibras colágenas observada en el grupo E (10µg/mL), lo que supone inducción de proteasas de la matriz. Puesto que la disminución en la aparición de cambios degenerativos como dilatación del retículo endoplasmático liso, pérdida de neurotúbulos, disolución de neurofilamentos y agregados de mitocondrias degeneradas es el principal efecto observado del FCE, y puesto que todos son componentes axoplamáticos, cuyo número y función son regulados por el cuerpo neuronal, es posible una acción del péptido a nivel central, controlando el transporte de elementos moleculares desde el cuerpo al axoplasma. Existen reportes que demuestran la existencia de transporte retrógrado de factores de crecimiento localmente producidos o administrados por vía exógena hacia el cuerpo neuronal, donde preservan a la célula de los cambios post-lesionales periféricos e influyen sobre la regeneración axonal [16, 17]. Bien documentado está el hecho de que la regeneración se afecta por trastornos en el transporte axonal, que es dependiente de la integridad del citoesqueleto y de la síntesis de proteínas como la actina y la tubulina [18]. Los estudios anteriores demuestran que la administración sistémica del factor neurotrófico ciliar, atenúa el déficit neuromuscular en modelos animales de degeneración de neuronas motoras. Además, el daño a nervios periféricos por traumatismos, agentes tóxicos o neuropatía diabética puede incrementar el transporte retrógrado y así la eficacia de factores neurotróficos exógenos [19]. Aunque este estudio no arroja evidencias en este sentido esto no niega la posibilidad de que así ocurra con el FCE. Pueden existir limitaciones de este modelo al aplicarlo a humanos por las diferencias en las características de las lesiones iniciales, las dimensiones del nervio, la complejidad de la arquitectura fascicular y el tiempo requerido para la regeneración. De cualquier modo ello es una aproximación válida a importantes enfermedades debilitantes y la demostración de un efecto hasta hoy no reportado del FCE, lo que abre insospechadas posibilidades a la investigación básica y clínica de esta molécula. Quedan aún algunas cuestiones que dilucidar como la formulación para la administración parenteral del péptido, el tiempo óptimo de evaluación, la dosis adecuada para lograr los efectos biológicos esperados, así como las poblaciones neuronales sensibles y las posibles interacciones con otros factores neurotróficos de efecto probado. Este estudio indica que existe el potencial para el empleo del FCE en el tratamiento de desórdenes de nervios periféricos. Referencias 1. Tadahisa U, Zhao Q, Lundborg G, Danielsen N. Effects of delayed nerve repair on regeneration of rat sciatic nerve. Restor Neurol Neurosci 1995;9:1-5. 2. Lundborg G. Nerve regeneration problems in a clinical perspective. Restor Neurol Neurosci 1990;1:297-302. 3. Guest JD, Rao A, Olson L, Bunge MB, Bunge RP. The ability of human Schwann cell grafts to promote regeneration in the transected nude rat spinal cord. Experimental Neurology 1997;148(2):502-22. 4. Nishi R. Neurotrophic factors: two are better than one. Science 1994; 265(5175):1052-3. 5. Ortíz-Hidalgo C, Weller RO. Peripheral nervous system. In: Stephen S. Stenberg, editor. Histology for pathologists, New York: Raven Press, Ltd, 1992:169-93. 6. Carpenter MK, Winkler C, Fricker R, Emerich DF, Wong SC, Greco C, et al. Generation and transplantation of FCE-responsive neural stem cells derived from GFAP-hNGF transgenic mice. Experimental Neurology 1997;148(1):187-204. 7. Morrison RS, Harley IK, Frances ML, Bradshaw RA. Trophic stimulation of culture neurons from neonatal rat brain by epidermal growth factor. Science 1987;238(4823):72-5. 8. Lewin GR, McKintosh E, McMahon. NMDA receptors and activity-dependent tuning of the receptive fields of spinal cord neurons. Nature 1994; 369(6480):482-5. 9. Patterson PH. Cytokines and the function of the mature nervous system. CR Acad Sci Paris, Sciences de la vie/Life sciences 1993;316:1150-7. 10. Vega JA, Vázquez E, Naves FJ, Calzada B, del Valle ME, Repressa JJ. Expression of epidermal growth factor receptor (EGFr) immunoreactivity in human cutaneous nerves and sensory corpuscles. Anat Rec 1994;240(1):125-30. 11. Tona A, Perides G, Rahemtulla F, Dahl D. Extracellular matrix in regenerating rat sciatic nerve. A comparative study on the localization of laminin, hyaluronic acid, and chondroitin sulfate proteoglycans, includin versican. J Histochem Cytochem 1993;41(4):593-9. 12. Nishida T. Fibronectin synthesis by the rabbit cornea effects of mouse epidermal growth factor and cyclic AMP analogs. Jpn J Ophtalmol 1984;28:196-202. 13. Tripathy RC. Prospects for epidermal growth factor in the management of corneal disorders. Surv Ophthalmol 1990;34:457-62. 14. Labrador RO, Buti M, Navarro X. Influence of collagen and laminin gels concentration on nerve regeneration after resection and tube repair. Experimental Neurology 1998;149(1):243-52. 15. Utley DS, Lewin SL, Cheng ET, Verity AN, Sierra D, Terris DJ. Brain-derived neurotrophic factor and collagen tubulization enhance funtional recovery after peripheral nerve transection and repair. Arch Otolaryngol Head Neck Surg 1996;122(4):407-13. 16. Curtis R, Adryan KM, Zhu Y, Harkness PJ, Lindsay RM, DiStefano PS. Retrograde axonal transport of ciliary neurotrophic factor is increased by peripheral nerve injury. Nature 1993;365(6443):253-5. 17. Curtis R, Scherer SS, Somogyi R, Adryan KM, Ip NY, Zhu Y, et al. Retrograde axonal transport of LIF is increased by peripheral nerve injury: correlation with increased LIF expression in distal nerve. Neuron 1994;12(1):191-204. 18. Mclean WG. The role of the axonal cytosqueleton in diabetic neuropathy. Neurochem Res 1997;22(8):951-6. 19. Senger DL, Campenot RB. Rapid retrograde tyrosine phosphorylation of trkA and other proteins in rat sympathetic neurons in compartmented cultures. J Cell Biol 1997;138(2):411-21. Copyright 1998 Elfos Scientiae The following images related to this document are available:Photo images[ba98038c.jpg] [ba98038a.jpg] [ba98038b.jpg] [ba98038d.jpg] [ba98038e.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}