 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 1999;16:75-82 Piruvato-carboxilasa de levadura Javier Menéndez División de Biotecnología Industrial. Centro de Ingeniería Genética y Biotecnología. AP 6162, CP 10600. Ciudad de La Habana, Cuba. Fax: (53-7) 33 6008; E-mail: yeastlab@cigb.edu.cu Code Number:BA99013 ABSTRACT Pyruvate carboxylase plays an important role in intermediary metabolism, catalyzing the formation of oxaloacetate from pyruvate and HCO3-. It thus provides oxaloacetate for gluconeogenesis and for replenishing tricarboxylic acid cycle for fatty acid, amino acid and neurotransmitter synthesis. The enzyme is highly conserved and it is found in a great variety of organisms including bacteria, yeasts, fungi and plants, as well as in higher organisms. The yeast Saccharomyces cerevisiae contains two structural genes for pyruvate carboxylase, which are regulated in different ways. On the other hand, Pichia pastoris has only one gene. This protein is a member of a group of biotin-dependent enzymes and the disruption of their coding genes results in the incapacity of the yeast to grow on a minimal medium containing glucose as the sole carbon source. This phenotype led to the isolation of mutants from these two yeasts, which are affected in the glucose repression of a series of genes. Keywords: pyruvate carboxilase, promoters, PYC1, PYC2, yeast RESUMEN La enzima piruvato-carboxilasa desempeña un papel importante en el metabolismo intermediario, al catalizar la síntesis de oxalacetato a partir de piruvato y HCO3-. Esta reacción suministra el oxalacetato necesario para la gluconeogénesis y para alimentar el ciclo de los ácidos tricarboxílicos, ya que algunos intermediarios del mismo se emplean como precursores en la síntesis de ácidos grasos, aminoácidos y neurotransmisores. Esta enzima se encuentra altamente conservada entre diferentes especies y los genes codificantes de la misma han sido aislados de bacterias, levaduras, hongos, plantas, así como de eucariotas superiores. En Saccharomyces cerevisiae existen dos genes codificantes de esta proteína, los cuales son regulados de manera diferencial. Por su parte, en Pichia pastoris se ha identificado solamente un gen. Esta proteína es miembro del grupo de las enzimas dependientes de biotina y la interrupción de la misma provoca un fenotipo de ausencia de crecimiento en medio mínimo cuando la glucosa se emplea como única fuente de carbono. Este fenotipo ha permitido el aislamiento, en estas dos levaduras, de mutantes en los que está afectada la represión por glucosa de una serie de genes. Palabras claves: levadura, piruvato-carboxilasa, promotores, PYC1, PYC2
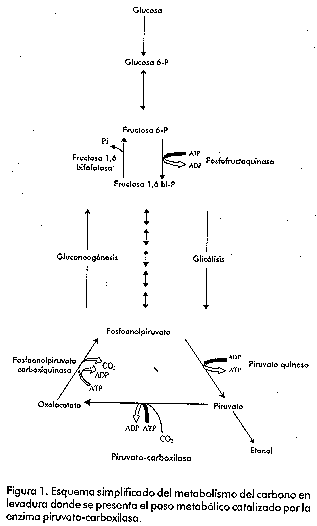
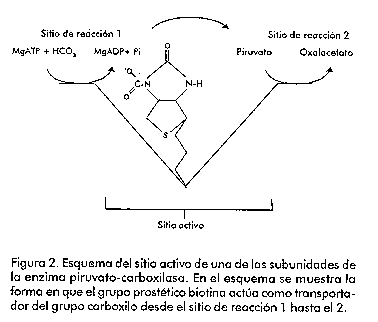
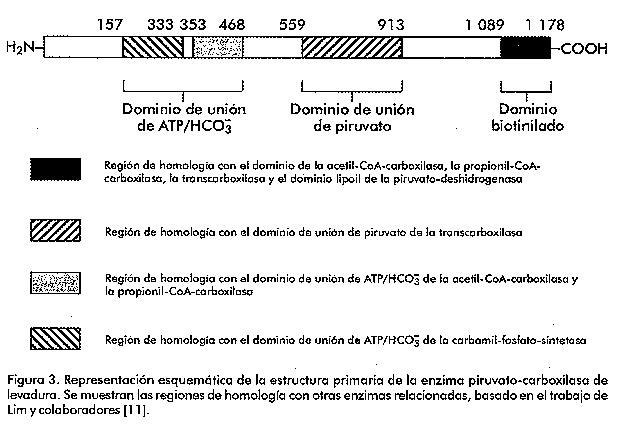
Introducción Las levaduras han sido empleadas por el hombre desde hace varios siglos para la obtención de bebidas alcohólicas y la producción de diferentes alimentos (como el pan). En nuestros días, las levaduras, en particular Saccharomyces cerevisiae, se emplean no sólo en diferentes industrias, sino también como modelos para el estudio de los mecanismos que controlan la expresión génica en los eucariotas. Los principales mecanismos que existen para el control de la transcripción en eucariotas son la activación o la represión de la maquinaria transcripcional [1]. La disponibilidad de nutrientes en el medio de cultivo promueve la generación de señales que van desde la membrana citoplasmática hasta el núcleo, y las levaduras responden a estas señales produciendo cambios en la expresión de diferentes genes [2]. Estos microorganismos pueden usar una gran variedad de compuestos orgánicos como fuente de carbono, sin embargo, emplean preferentemente hexosas como la glucosa y la fructosa. Cuando la levadura dispone de estos compuestos en el medio, los mismos son metabolizados por la ruta glicolítica y van principalmente hacia la fermentación alcohólica [3]. El crecimiento en presencia de otras fuentes de carbono como el glicerol o el lactato, requiere condiciones aeróbicas y es dependiente de la funcionalidad del ciclo de los ácidos tricarboxílicos. Algunos de los componentes del ciclo de los ácidos tricarboxílicos actúan como precursores en la biosíntesis de ácidos grasos, aminoácidos y neurotransmisores. La salida de componentes de este ciclo hacia otras rutas biosintéticas hace que el mismo deje de funcionar si no es alimentado. En las levaduras, existen dos rutas que llevan a cabo estas reacciones anapleróticas del ciclo de los ácidos tricarboxílicos. Una de estas rutas es el ciclo del glioxilato, que da lugar a malato y succinato partiendo de acetil-CoA, y la otra es la producción de oxalacetato a partir del piruvato, reacción catalizada por la enzima piruvato-carboxilasa (Figura 1). Figura 1. Esquema simplificado del metabolismo del carbono en levadura donde se presenta el paso metabólico catalizado por la enzima piruvato-carboxilasa. Hasta hace unos años, la ausencia de información acerca de la estructura de la enzima piruvato-carboxilasa frenó los esfuerzos que se venían realizando con el objetivo de comprender cómo funcionan y se regulan ésta y otras proteínas dependientes de biotina. Sin embargo, en los últimos años se ha acumulado una gran cantidad de información acerca de la estructura tridimensional y del mecanismo de acción de la enzima [4]. No obstante, el objetivo central de este trabajo versa sobre los genes codificantes de esta proteína en levaduras, el efecto fenotípico de su interrupción y sobre algunos aspectos que controlan su regulación, tanto a nivel catalítico, como de la síntesis de la enzima. La enzima piruvato-carboxilasa La enzima piruvato-carboxilasa ha sido descrita en una amplia variedad de organismos que incluyen bacterias, levaduras, hongos, invertebrados, vertebrados y plantas [5, 6]. Esta enzima pertenece al grupo de las carboxilasas dependientes de biotina, que abarca la acetil-CoA-carboxilasa (EC 6.4.1.2), la 3-metil-crotonil-CoA-carboxilasa (EC 6.4.1.4) y la propionil-CoA-carboxilasa (EC 6.4.1.3). En el caso particular de la piruvato-carboxilasa, el grupo prostético, la biotina, está unido a la enzima mediante un enlace amida con un residuo de lisina [7]. El sitio activo de las carboxilasas dependientes de biotina se cree que consiste en dos subsitios separados espacialmente, donde la biotina actúa como un transportador móvil del grupo carboxilo (Figura 2). La biotina se carboxila en un subsitio y se transfiere al segundo donde la molécula aceptora, en este caso el piruvato, es carboxilada [8]. Mediante el empleo del complejo enzima-carboxibiotina y de análogos del piruvato, se ha visto también que la unión de este último a la enzima induce el movimiento de la carboxibiotina al sitio donde ocurre la segunda etapa de la reacción [9]. Es en este sitio donde la carboxibiotina se desestabiliza. Por lo tanto, la reacción completa se puede representar como la suma de dos etapas: Figura 2. Esquema del sitio activo de una de las subunidades de la enzima piruvato-carboxilasa. En el esquema se muestra la forma en que el grupo prostético biotina actúa como transportador del grupo carboxilo desde el sitio de reacción 1 hasta el 2. En la enzima purificada de diferentes especies incluyendo S. cerevisiae, el sitio catalítico queda dentro de una cadena polipeptídica simple, de un peso molecular aproximado de 113-130 kDa. La forma activa de la enzima está compuesta por cuatro subunidades de este tipo, cada una de las cuales une una molécula de biotina [10]. Aislamiento de los genes que codifican la piruvato-carboxilasa de levadura La primera secuencia completa publicada del gen codificante de la enzima piruvato-carboxilasa, fue la correspondiente al gen de S. cerevisiae [11]. Este gen, al que nombraron PYC1, se aisló utilizando una sonda de ADN radiactiva sintetizada a partir de la secuencia consenso para el dominio de unión de biotina. Este gen se encuentra localizado en el cromosoma VII [12]. La secuencia de la proteína, deducida a partir de la secuencia de ADN, mostró que el mismo codifica una proteína de 1 178 aminoácidos. Estos autores compararon parte de la secuencia de esta proteína con partes de las secuencias de otras enzimas dependientes de biotina cuyas funciones son bien conocidas, y con las de enzimas que tienen funciones y propiedades catalíticas similares, pero que no son dependientes de biotina. Como resultado de esta comparación (Figura 3) vieron que la región entre los aminoácidos 559 y 913 de la enzima de S. cerevisiae tiene considerable homología con la región amino terminal de la subunidad 5S de la transcarboxilasa (EC 2.1.3.1), correspondiente al dominio de unión de piruvato. También, encontraron un alto nivel de homología de la región comprendida entre los aminoácidos 353 y 468 con parte de la subunidad a de la propionil-CoA-carboxilasa humana y también con parte de la acetil-CoA-carboxilasa de pollo. Estas tres proteínas tienen en común la capaci_dad de unir ATP y HCO3-. La región comprendida entre l aminoácidos 157 y 333, mostró una gran similitud con la mitad amino terminal del dominio de unión de ATP de la enzima carbamil-fosfato-sintetasa. Finalmente, la comparación de los últimos 90 aminoácidos del extremo carboxílico de la enzima de levadura, mostró un alto grado de similitud con el dominio biotinilado de la enzima E2 de la piruvato deshidrogenasa de Escherichia coli. Figura 3. Representación esquemática de la estructura primaria de la enzima piruvato-carboxilasa de levadura. Se muestran las regiones de homología con otras enzimas relacionadas, basado en el trabajo de Lim y colaboradores [11]. En 1991, dos grupos de investigadores publicaron independientemente que en S. cerevisiae existe otro gen codificante de la piruvato-carboxilasa [13, 14]. Este segundo gen, al que se le denominó PYC2, se aisló a partir de una genoteca, empleando como sonda radiactiva el gen PYC1. Este gen se encuentra localiza do en el cromosoma II [13]. Ambas isoenzimas tienen 90% de homología a nivel de secuencia de aminoácidos, tienen una localización citoplasmática y no presentan diferencias en los valores de Km para el ATP y el piruvato. Sin embargo, la proteína codificada por el gen PYC2 es más estable que la codificada por el gen PYC1, la cual pierde casi 100% de su actividad después de incubarse durante una noche a 4 °C [13]. Recientemente, se aisló el gen codificante de esta proteína en la levadura Pichia pastoris [15]. Para el aislamiento del mismo, se analizó la homología existente en varias regiones de esta proteína entre diferentes especies. Al gen aislado se le denominó PpPYC1 y a diferencia de lo que ocurre en S. cerevisiae, en la levadura P. pastoris, se demostró la existencia de un solo gen codificante de esta proteína. No está claro por qué unas especies presentan varias isoenzimas para una determinada función, mientras que otras sólo muestran una sola actividad. Se sabe que S. cerevisiae es un paradigma de genoma que se ha multiplicado parcialmente a partir de un genoma primitivo. Así, la secuenciación del genoma completo ha permitido mostrar que existen regiones enteras de un cromosoma que se hallan duplicadas en otro y que han sufrido alguna modificación en el curso de la evolución [16]. El análisis de la secuencia nucleotídica del gen aislado de la levadura P. pastoris, mostró que éste presenta un marco abierto de lectura de 1 189 aminoácidos y, por lo tanto, tiene 12 y 5 aminoácidos más que la codificada por los genes PYC1 y PYC2 de S. cerevisiae, respectivamente. La comparación de la secuencia de aminoácidos deducida a partir de la secuencia de ADN del gen clonado, con las regiones codificantes de los genes PYC1 y PYC2 de S. cerevisiae, reveló que existe una homología de 84% con estas proteínas. A su vez, la identidad fue de 72% en el dominio de unión de ATP (dominio A, aminoácidos del 157 al 333; dominio B, aminoácidos del 353 al 488), mientras que la región menos conservada es aquella donde se une la biotina (aminoácidos del 1 089 al 1 178), que muestra 58% de identidad (Figura 4). Figura 4. Comparación entre las secuencias aminoacídicas deducidas para la enzima piruvato-carboxilasa de S. cerevisiae y P. pastoris. Regulación alostérica de la enzima Uno de los aspectos de la función de la piruvato-carboxilasa que ha recibido mayor atención, es su regulación por acetil-CoA. En general, la actividad de la enzima de vertebrados es dependiente de acetil-CoA. La actividad de la enzima de procariotas muestra grados diferentes de dependencia de acetil-CoA; así, se ha observado que la enzima de Pseudomonas citronellolis no depende de éste [17], mientras que la Bacillus stearothermophilus muestra una dependencia total [18]. En le caso de la enzima de higado del pollo [19], asi como en la de Aspergillus nidulans [20] se ha visto mediante microscopía electrónica, que el acetil-CoA tiene un efecto estabilizador sobre la estructura de la enzima. Por su parte, la enzima de levadura es activa en ausencia de acetil-CoA, pues su actividad basal no depende de éste, sino de la presencia de iones potasio o de otros cationes monovalentes [21]. Sin embargo, el acetil-CoA tiene un efecto positivo sobre la actividad de la enzima. A concentraciones saturantes de acetil-CoA (Ka = 6,6 mM), la actividad máxima que se obtiene es tres o cuatro veces mayor que cuando está ausente. El efecto activador del acetil-CoA, combustible del ciclo de los ácidos tricarboxílicos, está en concordancia con la función anaplerótica de la enzima. Esta activación por acetil-CoA es inhibida competitivamente por el NADH, el cual, sin embargo, no afecta la actividad basal. Además del acetil-CoA, los acil-CoA de cadena larga son también potentes activadores de la enzima; de hecho, la Ka para el palmitoil-CoA es dos órdenes de magnitud menor que la del acetil-CoA [21]. En S. cerevisiae, la máxima actividad de la enzima depende, además, de la concentración relativa de los nucleótidos de adenina [22]. El aspartato, por otra parte, es un inhibidor no competitivo de la enzima (Ki = 1,9 mM), mientras que el oxalacetato muestra una inhibición competitiva con respecto al piruvato (Ki = 0,22 mM). La inhibición por aspartato parece ser un mecanismo de control específico, ya que otros ácidos dicarboxílicos no producen una inhibición sustancial [22]. Regulación transcripcional de los genes codificantes de la enzima piruvato-carboxilasa de S. cerevisiae Según se pudo apreciar en el acápite anterior, la enzima piruvato-carboxilasa es regulada alostéricamente y su síntesis parece ser constitutiva. Esto se ha visto al determinar la actividad en cultivos aeróbicos, tanto periódicos, como continuos, mediante el empleo de diferentes sustratos, en los que se observa que la actividad aumenta alrededor de dos veces [23, 24]. Por otra parte, en cultivos continuos crecidos en anaerobiosis y en condiciones en que la glucosa es limitante, la actividad es dos veces superior a la obtenida en condiciones aeróbicas [25, 26]. Por último, la adición de aspartato al medio de crecimiento reduce la actividad piruvato-carboxilasa en 50% en extractos de células dializados. En S. cerevisiae, existen dos genes codificantes de la enzima piruvato-carboxilasa [13, 14] que presentan una homología de 92% a nivel de secuencia de aminoácidos, pero sin alguna característica común en las regiones 5' no codificantes (Tabla 1). Este hecho parece indicar que existen diferencias en la regulación de la expresión de estos genes. Tabla 1. Secuencias en las regiones 5´no codificantes de los genes PYC1 y PYC2 que tienen homología con sitios potenciales de unión de proteínas reguladoras reportadas en S. cerevisiae. Tomado de Stucka y colaboradores [13].
La expresión de los genes PYC1 y PYC2 en S. cerevisiae se estudió a lo largo del crecimiento de la levadura en medio mínimo y en medio rico, utilizando la glucosa o el etanol como fuentes de carbono [27]. Estos autores analizaron la expresión de la enzima, tanto en cepas que portan los dos genes, como en cepas que portan uno de ellos interrumpido. En sus condiciones experimentales, los autores observaron que ambos genes PYC se transcriben en cultivos crecidos en glucosa y en etanol, aunque los niveles de ARN mensajero son dependientes de la fuente de carbono empleada. En el caso específico de los cultivos crecidos en glucosa, se observó que el gen PYC1 tenía una expresión constante, mientras que el gen PYC2 se expresaba mayoritariamente en los estadios iniciales del crecimiento. En el caso de los cultivos crecidos en etanol, la expresión del gen PYC1 fue superior a la del gen PYC2. Curiosamente, la expresión de PYC1 en los cultivos crecidos en etanol también fue superior a la expresión obtenida en los cultivos crecidos en glucosa. Este resultado es sorprendente, ya que se ha visto que el ciclo del glioxilato es el encargado de aportar el oxalacetato necesario durante el crecimiento en presencia de compuestos de dos átomos de carbono [28]. Una explicación posible para este resultado pudiera ser que los niveles de ARN mensajero determinados en estos experimentos, son relativos a los niveles de ARN total. Estas diferencias observadas pudieran deberse, en parte, a diferencias en el contenido de ARN ribosomal en las células crecidas en presencia de glucosa o etanol [29]. Con el objetivo de corroborar los resultados descritos anteriormente [27], se aislaron las regiones 5' no codificantes de los genes PYC1 y PYC2 mediante reacción en cadena de la polimerasa y se fusionaron al gen reportero lacZ [30]. La funcionalidad del sistema de fusión al gen lacZ se comprobó midiendo la actividad ß-galactosidasa en presencia de diferentes fuentes de carbono. En las condiciones experimentales empleadas, la actividad obtenida a partir de la expresión bajo el control del promotor del gen PYC1, fue dependiente de la fuente de carbono. Se obtuvieron los mayores niveles de actividad ß-galactosidasa en el medio con etanol, mientras que la obtenida a partir de la expresión bajo el control del promotor del gen PYC2 no muestra variaciones significativas en las diferentes fuentes de carbono. Resulta curioso el patrón de expresión obtenido con el promotor del gen PYC1, el cual coincide con el resultado del grupo de Brewster [27]. Como se mencionó anteriormente, durante el crecimiento en presencia de compuestos de dos átomos de carbono, el ciclo del glioxilato parece ser el encargado de suministrar el oxalacetato necesario para el funcionamiento continuo del ciclo de Krebs, y la piruvato-carboxilasa aparece como innecesaria. En este sentido, se pueden mencionar los resultados de un estudio en el que un doble mutante D(pyc1, pyc2) es capaz de crecer en presencia de piruvato, ya que el ciclo del glioxilato está desreprimido [31]. No obstante, los resultados indican claramente que el ciclo del glioxilato, así como la reacción catalizada por la piruvato-carboxilasa, son esenciales para el suministro del oxalacetato necesario durante la gluconeogénesis. Sería interesante estudiar si la expresión de los genes que codifican las enzimas del ciclo del glioxilato influye de alguna manera en la del gen PYC1. Esto podría realizarse, por ejemplo, mediante la medición de los niveles de actividad piruvato-carboxilasa en cepas icl1-. Para identificar dentro de estos promotores las regiones responsables de su regulación, se realizaron deleciones seriadas de los mismos en el sentido 5'. Las deleciones obtenidas en el caso del promotor del gen PYC1, permitieron identificar dos regiones cuya eliminación tiene un efecto marcado sobre la transcripción del gen lacZ [30]. La primera deleción actúa negativamente y fue identificada entre las posiciones -483 y -331 con respecto al ATG de iniciación. Un análisis de esta región del promotor, mostró que en la misma aparecen tres secuencias homólogas a la región represora URS1 del gen CAR1, el cual codifica la enzima arginasa [32]. Estas secuencias incluyen un núcleo central conservado de la secuencia 5´-CCGCC-3´ y en particular, la comprendida entre -432 y -424 (5´-AGCCGCCCA-3´), es idéntica a una secuencia que aparece en los promotores de los genes CHA1 [33] y HOP1 [34]. Este URS ha sido identificado como regulador negativo de la transcripción de varios genes no relacionados entre sí (Tabla 2) y es capaz de unir una o varias proteínas en experimentos de retraso en gel. Bien pudiera ser que estas proteínas estuviesen implicadas en una forma de regulación negativa de la expresión del gen PYC1. Tabla 2. Homologías entre las regiones URS presentes en diferentes genes, con respecto a la región URS1 del gen CAR1.
La segunda región identificada dentro del promotor PYC1 se encuentra entre -330 y -298. Esta región actúa como un elemento activador de la transcripción (UAS), el cual parece ser necesario para la expresión del gen, tanto durante el crecimiento en presencia de glucosa, como durante el crecimiento en presencia de etanol. Además, este fragmento fue capaz de formar un complejo específico ADN-proteína cuando se empleó como sonda en experimentos de retraso en gel, en los que se utilizaron extractos nucleares preparados a partir de cultivos crecidos en condiciones de represión y desrepresión [30]. Sin embargo, este complejo desaparece cuando en estos experimentos se emplean extractos preparados a partir de cepas que presentan mutados los genes rtg1 y rtg2. Estos genes participan en la regulación transcripcional de la enzima citrato sintasa no mitocondrial [35]. Curiosamente, también se ha demostrado que estas mutaciones disminuyen en 50% los niveles de actividad piruvato-carboxilasa [36]. Es interesante señalar que cepas de levaduras que tienen interrumpido uno de los dos genes RTG1 y RTG2 o ambos, no crecen en medio mínimo con glucosa o acetato como únicas fuentes de carbono, y son auxotróficas para aspartato o glutamato [35]. Este fenotipo es típico de células que presentan afectado el ciclo de los ácidos tricaboxílicos o el ciclo del glioxilato. A su vez, ese mismo fenotipo es el que presentan las cepas en las que se ha eliminado la función de la piruvato-carboxilasa [13]. Por lo tanto, esas proteínas, bien directa o indirectamente, podrían estar implicadas en la regulación de la transcripción de al menos el gen PYC1. En el caso de los elementos responsables de la transcripción del gen PYC2, los experimentos realizados permitieron identificar al menos una de las regiones necesarias para su regulación. En este caso, se vio que la región comprendida entre -417 y -291 es necesaria para la transcripción de este gen cuando la glucosa se emplea como fuente de carbono. No obstante, con esta región (entre -417 y -291), no se pudo obtener la formación de complejos específicos ADN-proteína [30]. El análisis de la secuencia de ADN de esta región del promotor PYC2, mostró que contiene dos secuencias repetidas del pentanucleótido 5´-CTTCC-3´, al que se le denomina bloque CT. Este pentanucleótido ha sido descrito en el promotor del gen PGK [37], el cual contiene una región activadora compuesta de varios elementos entre los que se encuentran tres secuencias repetidas del mismo. Por último, a pesar de que en la secuencia del promotor del gen PYC2 existen varios sitios potenciales de unión de proteínas reguladoras (Tabla 1), la eliminación de estos sitios no provoca ninguna variación significativa de la actividad ß-galactosidasa [30]. Defecto metabólico causado por la interrupción de los genes PYC El efecto de la interrupción de los genes PYC en la cepa portadora de varias auxotrofías, W303, ha sido publicado previamente [13], y se ha observado que los mutantes que expresan sólo uno de los dos genes no mostraron ningún fenotipo evidente. Sin embargo, estos mismos autores reportaron que la interrupción de los dos genes produjo una cepa incapaz de crecer en medio mínimo que contiene glucosa como única fuente de carbono. El crecimiento de esta cepa deficiente en la enzima piruvato-carboxilasa, empleando glucosa como única fuente de carbono, fue posible solamente cuando se empleó aspartato como fuente de nitrógeno. Un fenotipo similar se ha reportado en un doble mutante D(pyc1, pyc2) obtenido en la cepa auxotrófica DBY746 [27]. Estos autores observaron un requerimiento parcial de aspartato en la cepa que tiene interrumpido solamente el gen PYC1, lo que sugiere que la contribución relativa de las isoenzimas a la actividad total es dependiente de la cepa. En el caso de P. pastoris, se pudo observar que la interrupción del único gen de la piruvato-carboxilasa presente en esta levadura, produjo un fenotipo similar al encontrado en S. cerevisiae cuando se interrumpen los dos genes PYC [15]. En este caso, la adición de aspartato o glutamato permitió el crecimiento de la cepa que tiene el gen interrumpido, a diferencia de lo que ocurre en S. cerevisiae, donde el glutamato no permite el crecimiento cuando está presente la doble interrupción [13]. No está claro si esto es una particularidad de la cepa usada, o si está relacionado con alguna peculiaridad del transporte de aminoácidos en esa especie. Aislamiento de mutaciones que suprimen el fenotipo en levaduras La incapacidad que muestran las cepas de S. cerevisiae y P. pastoris deficientes en la actividad piruvato-carboxilasa, de crecer en medio mínimo con glucosa como única fuente de carbono, se debe a que el ciclo del glioxilato no puede suplir el papel que realiza la piruvato-carboxilasa en esas condiciones. No obstante, se han aislado mutaciones supresoras, tanto en S. cerevisiae [31, 38], como en P. pastoris [15], que permiten el crecimiento en medio mínimo con glucosa como única fuente de carbono. Las mutaciones identificadas en S. cerevisiae, BPC1-1 [31] y DGT1-1 [38] son dominantes y alélicas entre sí. En el caso específico de la mutación DGT1-1, se produce una disminución de la entrada de glucosa a la célula. Tal disminución del transporte de glucosa hacia el interior de la célula, al parecer disminuye la represión catabólica que ésta ejerce sobre una gran variedad de genes. Teniendo en cuenta que en levadura la única vía conocida que puede rellenar el ciclo de los ácidos tricarboxílicos en ausencia de la enzima piruvato-carboxilasa es el ciclo del glioxilato [28], las bases moleculares de esta supresión parecen estar relacionadas con una afectación de la represión catabólica que se ejerce sobre el gen que codifica la enzima isocitrato liasa. Esta afectación en la represión permite la expresión del gen codificante de la isocitrato-liasa, en condiciones en las que normalmente está reprimido. Estos autores determinaron la actividad iso-citrato-liasa en las cepas portadoras de la mutación supresora, y tanto en S. cerevisiae [31, 38], como en P. pastoris [15], observaron que la enzima estaba desreprimida durante el crecimiento en presencia de glucosa. En todos los casos, estas mutaciones supresoras no fueron específicas sólo del ciclo del glioxilato, ya que ambas mutaciones producen la desrepresión de varios genes normalmente reprimidos por glucosa. No obstante, existen diferencias entre el fenotipo causado por las mutaciones BPC1-1/DGT1-1 y por el supresor aislado en P. pastoris. Por una parte, no se alcanza el mismo grado de desrepresión para todas las enzimas analizadas en las cepas portadoras de las mutaciones BPC1-1/DGT1-1, mientras que en P. pastoris, la mutación supresora causa una desrepresión total de todas las enzimas. Además, en P. pastoris la mutación supresora produce un incremento en la actividad de la enzima glutamato deshidrogenasa (dependiente de NAD+) por encima de los valores obtenidos en la cepa salvaje, en todas las condiciones analizadas. Este incremento fue similar al observado en mutantes del gen ure2 de S. cerevisiae, para la actividad de la glutamato deshidrogenasa y de otras enzimas relacionadas con el metabolismo del nitrógeno cuando el glutamato o la glutamina son utilizados como fuente de este elemento. Sin embargo, hasta ahora no hay evidencias de que el gen ure2 esté relacionado con la represión catabólica por la fuente de carbono [39]. Según se mencionó en el caso específico de la mutación DGT1-1, el principal defecto que aparece es una reducción drástica de la entrada de glucosa a la célula. Esta mutación perturba la expresión de algunos de los transportadores de este azúcar [38], eliminando la expresión de los genes HXT1 y HXT3, los cuales se expresan a altos niveles durante el crecimiento de la cepa salvaje en presencia de glucosa. Resultados no publicados del laboratorio de Gancedo (Universidad Autónoma de Madrid, España) indican que todas esas mutaciones son alélicas del gen salvaje MTH1 [40] y corresponden a proteínas defectuosas, ya que una deleción total del gen no produce el fenotipo descrito (Gancedo, 1997). En el caso de P. pastoris, no se ha determinado en qué grado está afectado el transporte de glucosa en la cepa que porta la mutación supresora. No obstante, teniendo en cuenta que esta mutación fue aislada utilizando un sistema de selección similar al empleado en el aislamiento de la mutación BPC1-1, es muy probable que en esta cepa también esté afectado el transporte de glucosa. Conclusiones La enzima piruvato-carboxilasa desempeña una función anaplerótica importante al suministrar el oxalacetato necesario para el crecimiento de la levadura en presencia de diferentes fuentes de carbono. En S. cerevisiae, a diferencia de lo que ocurre en P. pastoris, existen dos isoenzimas que contribuyen a la actividad total de la enzima piruvato-carboxilasa. El papel de los dos genes PYC aún no ha podido ser dilucidado, debido a las diferencias obtenidas en los experimentos realizados. No obstante, sí ha quedado claro que en S. cerevisiae ambos genes se regulan de manera diferencial. El empleo de mutantes que no presentan actividad piruvato-carboxilasa, ha permitido la identificación de mutaciones que suprimen este defecto. Las consecuencias de estas mutaciones, tanto en S. cerevisiae, como en P. pastoris, han mostrado ser no específicas solamente del ciclo del glioxilato, afectando también otras enzimas cuyos genes son normalmente reprimidos por glucosa. Hasta la fecha, ésta es la primera mutación aislada en la levadura P. pastoris que afecta la represión por glucosa de una serie de genes. Creemos, por lo tanto, que la caracterización detallada de esta mutación puede contribuir significativamente a la comprensión de los mecanismos de regulación de la expresión génica en P. pastoris, y proporcionar herramientas que puedan redundar en el mejoramiento de las aplicaciones biotecnológicas de esta levadura. Agradecimientos Quisiera agradecer a Bianca García, Liliana Basabe y al doctor José de la Fuente por la lectura crítica del manuscrito y las sugerencias brindadas. Este trabajo fue financiado, en parte, por el proyecto PB94-0091-CO2-01 de la DGICYT de España, por la Acción Especial Bio95-2051 y por el proyecto BIO4-CT95-0132 de la Unión Europea. Es parte también del proyecto From Gene to Products in Yeast: A Quantitative Approach, financiado por la Comunidad Europea (DGXII Framework IV Program Cell Factories). Referencias 1. Gancedo JM. Carbon catabolite repression in yeast. Eur J Biochem 1992; 206: 297-313. 2. Johnston M, Carlson M. Regulation of carbon and phosphate utilization. In: Jones EW, Pringle JR, Broach JR, editors. The molecular and cellular biology of the yeast Saccharomyces: Gene expression. Vol 2. New York: Cold Spring Harbor Laboratory Press; 1992. p.193-281. 3. Gancedo C, Serrano R. Energy-yielding metabolism. In: Rose AH, Harrison JS, editors. The Yeasts. Vol 3. 2nd ed. London: Academic Press; 1989. p.205-59. 4. Attwood PV. The structure and the mechanism of action of pyruvate carboxylase. Int J Biochem Cell Biol 1995;25:231-49. 5. Wallace JC. Distribution and biological function of pyruvate carboxylase in nature. In: Keech DB, Wallace JC, editors. Pyruvate carboxylase. Boca Raton: CRC Press; 1985. p.5-63. 6. Wuterle ES, Nikolau BJ. Plants contain multiple biotin enzymes: discovery of 3 methylcrotonyl-CoA carboxylase, propionyl-CoA carboxylase and pyruvate carboxylase in the plant kingdom. Arch Biochem Biophys 1990;278:179-86. 7. Rylatt DB, Keech DB, Wallace JC. Pyruvate carboxylase: isolation of the biotin-containing tryptic peptide and determination of its primary sequence. Arh Biochem Biophys 1977; 183:113-22. 8. Keech DB, Attwood PV. The reaction mechanism. In: Keech DB, Wallace JC, editors. Pyruvate carboxylase. Boca Ratón: CRC Press; 1985. p.109-40 9. Goodall GJ, Baldwin GS, Wallace JC, Keech DB. Factor that influence the translocation of the N-carboxybiotin moiety between the two sub-sites of pyruvate carboxylase. Biochem J 1981;199: 603-9. 10. Wallace JC, Easterbrook-Smith SE. The structure of pyruvate carboxylase. In: Keech DB, Wallace JC, editors. Pyruvate carboxylase. Boca Ratón: CRC Press; 1985. p.65-108. 11. Lim F, Morris CP, Occhiodoro F, Wallace JC. Sequence and domain structure of yeast pyruvate carboxylase. Biol Chem 1988; 263:11493-7. 12. Morris SC, Lim F, Wallace JC. Yeast pyruvate carboxylase: gene isolation. Biochem Biophys Res Comm 1987;145:390-6. 13. Stucka R, Dequin S, Salmo JM, Gancedo C. DNA sequences in chromosomes II and VII code for pyruvate carboxylase isoenzymes in Saccharomyces cerevisiae: analysis of pyruvate carboxylase-deficient strains. Mol Gen Genet 1991;229:307-15. 14. Walker ME, Vale DL, Rohde M, Devenish RJ, Wallace JC. Yeast pyruvate carboxylase: Identification of two genes encoding isoenzymes. Biochem Biophys Res Comm 1991; 176:1210-7. 15. Menéndez J, Delgado J, Gancedo C. Isolation of the Pichia pastoris PYC1 gene encoding pyruvate carboxylase and identification of a suppresor of the pyc phenotype. Yeast 1998;14:647-54. 16. Wolfe KH, Shields DC. Molecular evidence for an ancient duplication of the entire yeast genome. Nature 1997;387:708-13. 17. Taylor BL, Routman S, Utter MF. The control of synthesis of pyruvate carboxylase in Pseudomonas citronellolis. Evidence from double labeling studies. J Biol Chem 1975; 250:2376-82. 18. Sundaram TK, Cazzulo JJ, Kornberg HL. Anaplerotic CO2 fixation in mesophilic and thermophilic bacilli. Biochem Biophys 1969; 192:355-7. 19. Mayer F, Wallace JC, Keech DB. Further electron microscopic studies on pyruvate carboxylase. Eur J Biochem 1980;112:265-72. 20. Osmani SA, Mayer F, Martson FAO, Selmes IP, Scrutton MC. Pyruvate carboxylase from Aspergillus nidulans: effects of regulatory modifiers on the structure of the enzyme. Eur J Biochem 1984;139:509-18. 21. Myers DE, Tolbert B, Utter M F. Activation of yeast pyruvate carboxylase: interaction between acyl-Coenzyme A compounds, aspartate and substrates of reaction. Biochem 1983;22:5090-6. 22. Miller AL, Atkinson DE. Response of yeast pyruvate carboxylase to the adenylate energy charge and other regulatory parameters. Arch Biochem Biophys 1972;152:531-8. 22. Palacián E, de Torrontegui G, Losada M. Inhibition of yeast pyruvate carboxylase by L-aspartate and oxaloacetate. Biochem Biophys Res Comm 1966;24:644-9. 23. Divjak S, Mor JR. On the activity of carbon dioxide fixation in growing yeasts. Arch Microbiol 1973;94:191-99. 24. Haarasilta S, Taskinen L. Location of three key enzymes of gluconeogenesis in baker's yeast. Arch Microbiol 1977;113:159-161. 25. Haarasilta S, Oura E. Effect of aeration on the activity of gluconeogenic enzymes in Saccharomyces cerevisiae growing under glucose limitation. Arch Microbiol 1975;106: 271-3. 26. Oura E, Haarasilta S, Londesborough J. Carbon dioxide fixation by baker´s yeast in a variety of growth conditions. J Gen Microbiol 1980;118:51-8. 27. Brewster NK, Val NK, Walker ME, Wallace JC. Regulation of pyruvate carboxylase isozyme (PYC1, PYC2) gene expression in Saccharomyces cerevisiae during fermentative and non-fermentative growth. Arch Biochem Biophys 1994;311:62-71. 28. Schöler A, Schüller HJ. Structure and regulation of the isocitrate lyase gene ICL1 from the yeast Saccharomyces cerevisiae. Curr Genet 1993;23:375-81. 29. Pronk JT, Steensma HY, van Dijken JP. Pyruvate metabolism in Saccharomyces cerevisiae. Yeast 1996;12:601-10. 30. Menéndez J, Gancedo C. Regulatory regions in the promoters of Saccharomyces cerevisiae PYC1 and PYC2 genes encoding isoenzymes of pyruvate carboxylase. FEMS Microbiol lett 1998;2:345-352. 31. Blázquez MA, Gamo FJ, Gancedo C. A mutation affecting carbon catabolite repression suppresses growth defect in pyruvate carboxylase mutants from Saccharomyces cerevisiae. FEBS lett 1995;377:197-200. 32. Luche RM, Sumrada R, Cooper TG. A cis-acting element present in multiple genes serves as a repressor protein-binding site for the yeast CAR1 gene. Mol Cell Biol 1990; 10:3884-95. 33. Bornaes C, Ignjatovic MW, Schjerling P, Kielland-Brandt MC, Holmberg S. A regulatory element in the CHA1 promoter which confers inducibility by serine and threonine on Saccharomyces cerevisiae genes. Mol Cell Biol 1993;13:7604-11. 34. Vershon AK, Hollingsworth NM, Johnson AD. Meiotic Induction of the yeast HOP1 gene is controlled by positive and negative regulatory sites. Mol Cell Biol 1992;12:3706-14. 35. Liao X, Butow R. RTG1 and RTG2: Two genes required for a novel path of communication from mitochondria to the nucleus. Cell 1993;72:61-71. 36. Small WC, Brouder RD, Sandor A, Fedorova N, Li G, Butow RA, et al. Enzymatic and metabolic studies on retrograde regulation mutants of yeast. Biochem 1995;34: 5569-76. 37. Chambers A, Stanway C, Kingsman AJ, Kingsman S. The UAS of the yeast PGK gene is composed of multiple functional elements. Nucl Acids Res 1988;16:8245-60. 38. Gamo FJ, Lafuente MJ, Gancedo C. The mutation DGT1-1 decreases glucose transport and alleviates carbon catabolite repression in Saccharomyces cerevisiae. J Bacteriol 1994;176:7423-29. 39. Minehart PL, Magasanik B. Sequence and expression of GLN3, a positive nitrogen regulatory gene of Saccharomyces cerevisiae encoding a protein with a putative zinc finger DNA-binding domain. Mol Cell Biol 1991;11: 6216-28. 40. Hubbard EJA, Jiang R, Carlson M. Dosage dependent modulation of glucose-repression by MSN3 (STD1) in Saccharomyces cerevisiae. Mol Cell Biol 1994;14:1972-8. 41. Bishop DK, Park D, Xu L, Kleckner N. DMC1: a meiosis-specific yeast homologous of E. coli recA gene required for recombination, synaptonemal complex formation, and cell cycle progression. Cell 1992;69:439-56. 42. Wang HT, Frackman S, Kowalisyn J, Esposito RE, Elder RT. Developmental regulation of SPO13, a gene required for separation of homologous chromosomes at meiosis I. Mol Cell Biol 1987;7:1425-35. 43. Engebrecht J, Roeder GS. MER1, a yeast gene required for chromosome pairing and genetic recombination, is induced in meiosis. Mol Cell Biol 1990;10:2379-89. Recibido en julio de 1998. Aprobado en octubre de 1998. The following images related to this document are available:Line drawing images[ba99013d.gif] [ba99013c.gif] [ba99013b.gif] [ba99013a.gif] [ba99013e.gif] |
| |||||||||

{kind=link}
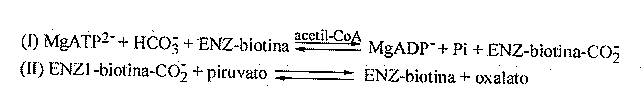{kind=link}
{kind=link}
{kind=link}
{kind=link}