 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 1999;16:103-108 Effect of Different Adjuvants and Immunomodulators on the Humoral Immune Response of Rabbits and Mice against HIV-1-derived Multi-epitope Polypeptides @ Antonieta M Herrera,1 Dania Vázquez,1 Leonor
Navea,2 Leonor Lobaina,2 1Vaccine Division, AIDS Department. Center for Genetic
Engineering and Biotechnology. PO Box 6162, Havana 10600, Cuba.
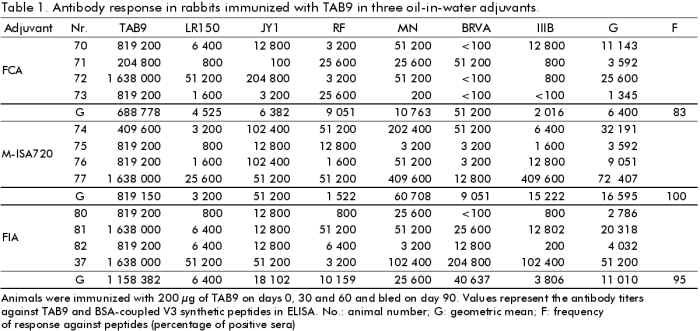
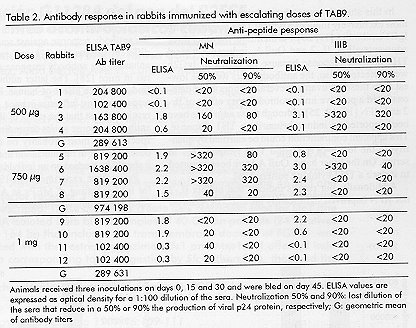
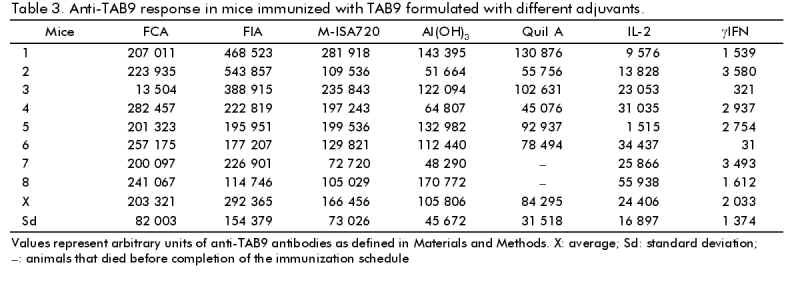
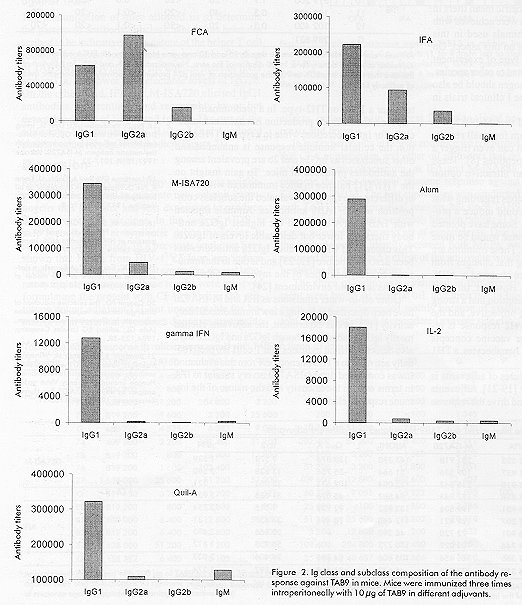
Phone: (53-7) 21 8008, 21 8466. Fax: (53-7) 21 8070, 33 6008 Code Number: BA99018 ABSTRACT The third variable region (V3 loop) of the human immunodeficiency virus (HIV) external glycoprotein gp120 contains the principal neutralizing domain of this protein. Our group has developed multi-epitope polypeptides (MEP), bearing several copies of the V3 loop from different HIV-1 isolates. These chimeric proteins have been able to elicit broadly reactive neutralizing antibodies when administered in Complete Freund's Adjuvant (CFA). For human vaccines, a less reactogenic adjuvant is required. The MEPs TAB9 and TAB13 contain the V3 region from six and eight HIV-1 isolates, respectively, fused to the amino terminus of the Neisseria meningitidis P64K protein. In this paper we describe the effect of several adjuvants and immunomodulators on the antibody response against these MEPs in rabbits and mice. Oil adjuvants proved to be more efficient in promoting the antibody response against MEPs than Alum, Quil A or combinations of Alum with IL-2 and gIFN. The subclass composition of the antibody response was very dependent on the adjuvant employed. CFA induced high levels of IgG2a and IgG2b, while for the rest of the products IgG1 was predominant. We also concluded that the novel oil adjuvant Montanide ISA720 is as efficient as CFA or Incomplete Freund's Adjuvant in stimulating the humoral response in mice and rabbits and therefore, it was selected for further studies in primates. Keywords: adjuvants, HIV, immunomodulators, vaccines, V3 loop RESUMEN La tercera región variable (el lazo V3) de la glicoproteína externa gp120 del virus de la inmunodeficiencia humana (VIH) contiene el dominio principal de neutralización de esta proteína. Nuestro grupo ha desarrollado el concepto de polipéptidos multiepitópicos (PME), proteínas quiméricas que portan varias copias del lazo V3 de diferentes aislamientos del VIH-1. Estas proteínas han sido capaces de inducir una amplia respuesta de anticuerpos neutralizantes cuando se administran en Adyuvante Completo de Freund (ACF). Para vacunas de uso humano, se requiere un adyuvante menos reactogénico. Los PME TAB9 y TAB13 contienen la región V3 de seis y ocho aislamientos de VIH-1, respectivamente, fusionados al extremo amino de la proteína P64K de Neisseria meningitidis. En este artículo se describe el efecto de diferentes adyuvantes e inmunomoduladores en la respuesta de anticuerpos contra estos PME en conejos y ratones. Los adyuvantes oleaginosos estimularon la respuesta inmune humoral contra los PME de forma más eficiente que la alúmina, el Quil A o combinaciones de alúmina con IL-2 e IFNg. La composición de subclases de Ig específica para los PME varió en dependencia del adyuvante empleado. Los mayores niveles de IgG2a e IgG2b se obtuvieron con ACF, mientras que otros adyuvantes indujeron IgG1 predominantemente. También concluimos que el nuevo adyuvante oleaginoso Montanide ISA720 es tan eficiente como el ACF o el Adyuvante Incompleto de Freund en la estimulación de la respuesta humoral en ratones y conejos, y fue seleccionado para estudios posteriores en primates. Palabras claves:adyuvantes, inmunomoduladores, lazo V3, vacunas, VIH Introduction The vast majority of the current available viral vaccines are composed of either attenuated or inactivated whole viruses. Due to their high intrinsic immunogenicity, these can induce a protective immune response in the absence of immunological adjuvants, or simply by adsorption to Alum. On the other hand, the only subunit vaccine available, the anti-hepatitis B vaccine, forms a "virus-like" particle that resembles the natural pathogen and is also strongly immunogenic. However, there is an increasing number of viral diseases in which the aforementioned vaccination approaches cannot be implemented. The use of small peptides and recombinant subunits has been extensively investigated in the last years as components for new generation and extremely safe vaccines, but in most of these cases, the immunostimulatory capacity of Alum is limited. This fact has prompted the evaluation of novel, more potent immunological adjuvants intended for human applications. Since the Acquired Immune Deficiency Syndrome (AIDS) was first described [1] and the global implications of this pandemic were recognized, several research groups have been focused on the prevention of an infection with HIV. AIDS is one of these examples where the use of conventional vaccines is clearly not recommendable. Moreover, given the extreme genetic divergence of HIV, a conventional vaccine using a specific viral subtype would probably not be able to confer protection against the variants existing worldwide [2]. Other strategies based on recombinant and synthetic subunit vaccines designed to generate a broadly reactive antibody response might be more efficient to accomplish this goal [3, 4]. Our group has reported the generation of multi-epitope polypeptides (MEP) carrying in tandem the V3 regions of the gp120 protein from several HIV-1 isolates as a novel alternative to generate a wide antibody response against V3 loop [5, 6]. The aim of this study was to evaluate the capacity of several adjuvants to enhance and modulate the humoral immune response in rabbits and mice, against two of these MEPs, denominated TAB9 and TAB13, and to select the more attractive among them for further development of pre-clinical and clinical studies. Six adjuvants and two immunomodulators were selected for this study on the basis of their availability and their documented properties. Besides Alum, which is the only adjuvant currently approved for human vaccines, we evaluated Complete Freund's Adjuvant (CFA) and Incomplete Freund's Ajuvant (IFA), which have been traditionally used in animal experiments, but their application in humans has been limited by the extensive reactogenicity they produce [7]. Another oil adjuvant, Montanide ISA 888 (M-ISA888), has been successfully used in veterinary vaccines, but its composition is rather similar to IFA. We also included the adjuvant Montanide ISA 720 (M-ISA720), which is based in metabolizable animal oil and is thought to be less reactogenic in humans [8]. Quil A is a saponin fraction from Quilaria saponaria that has been widely used in animal experiments. Its more purified derivative, QS21, has been recently employed in several clinical trials [9]. Finally, IL-2 and gIFN were employed due to their documented immunomodulatory effect on antibody response [10, 11]. Materials and Methods Animals Female Balb/c mice, of six weeks of age (20 g of weight), were obtained from CENPALAB, Havana, Cuba. New Zealand white female rabbits between 2.0 and 2.5 kg were also supplied by CENPALAB. HIV-1 multi-epitope polypeptides TAB9 is a MEP containing six copies of the central region of the V3 loop from isolates LR150, JY1, RF, MN, BRVA and IIIB in this order, fused to the amino terminal 47 amino acids from P64K protein of Neisseria meningitidis. TAB13 contains two additional V3 regions from consensus sequences C8 and C6 according to the phenetic classification system proposed by Myers et al. [12, 13]. Protein purification Cultures of E. coli W3110 trpA905 cells transformed with plasmids pTAB9 or pTAB13 bearing sequences encoding proteins TAB9 and TAB13 respectively, were harvested and suspended in Tris 10 mM, EDTA 1 mM, pH 8.0 (TE) at 0.1 g of biomass/mL. Lysozyme was added to a final concentration of 25 mg/mL, and the cells were incubated for 1 h with gentle, occasional mixing. After sonication at 4 °C and centrifugates, the pellet was washed with TE + 4M urea and then extracted in TE + 8M urea. DNA was precipitated with 1% streptomycin sulfate before the chromatographic step. This material was centrifuged and the supernatant was loaded onto a 10 x 250 mm C4 reverse-phase column for high-performance liquid chromatography (HPLC). A continuous gradient from 10 to 80% acetonitrile in 0.1% trifluoroacetic acid was used for the elution. A flow rate of 1.5 mL/min was employed and the absorbance at 280 nm was monitored. The purity of the final preparation was assessed by SDS-PAGE [14]. Preparation of adjuvants Aluminum hydroxide (Alum) . Aluminum hydroxide (Alhydrogel, Superfos Biosector, Vedbaek, Denmark) was mixed with protein at a 70:1 (Alum:protein) ratio. The mixture was incubated with gentle agitation for 1 h at 37 °C. The preparation was centrifuged for 15 min at 10 000 rpm (Himac centrifuge, Hitachi®, Japan) and protein concentration was determined by the method of Bradford [15]. Interleukin-2 (IL-2) and gamma interferon (gIFN). Human recombinant IL-2 or gIFN (Heber Biotec, Havana, Cuba) were added to the protein solution just before injection. Ten thousand units of IL-2 and 1 mg of gIFN were used per mouse, per inoculation. Quil A. Quil A Saponin (Superfos Biosector, Vedbaek, Denmark) was suspended in sterile water and added to the protein solution. A dose of 25 mg per animal was employed. CFA and IFA. The protein was mixed with an equivalent volume of either CFA or IFA (Sigma, St. Louis, Miss) and emulsified by strong agitation during 15 min in a homogenizer (Ultraturrax). The quality of the emulsion was tested by sampling a drop into ice-cold water. M-ISA720 and M-ISA888. The emulsions composed by either M-ISA720 or M-ISA888 (Seppic, Paris, France) and TAB9 or TAB13 were prepared following a similar procedure to that described for Freund's adjuvants. These preparations contained 30% of aqueous phase and 70% of oil phase as suggested by the manufacturers. Immunization procedures Comparison of adjuvants in rabbits. Groups of four New Zealand white rabbits were immunized three times with 200 µg of TAB9 or TAB13 emulsified in either CFA, IFA, M-ISA888, M-ISA720, Quil A, Alum, Alum + IL-2 and Alum + gIFN on days 0, 30, and 60. The immunogens were administered subcutaneously. Animals were bled 30 days after the last inoculation and the sera titrated against TAB9 and V3 peptides in ELISA (see below). Escalating doses of TAB9. Three groups of New Zealand white rabbits received three inoculations on days 0, 15 and 30 and were bled on day 45. The doses were 500, 750 and 1 000 µg of TAB9. ELISA titers and neutralizing antibodies against strains MN and IIIB were also measured. Comparative evaluation of different adjuvants in mice. Nine groups of eight mice each were immunized with 10 µg of TAB9, three times at two weeks intervals. The immunogens were administered intraperitoneally in 0.1 mL for each animal. Two weeks after the third inoculation, blood was extracted from the plexus and animals were sacrificed. Serum was obtained and stored until use. Conjugation of peptides to bovine seroalbumin (BSA) Peptides were coupled to BSA using the heterobifunctional agent maleimidobenzoyl-N-hydroxysuccinimide ester (MBS) (Boehringer Mannheim, Germany). Five milligrams of carrier protein were dissolved in the coupling solution (0.01 M phosphate buffer, pH 6.0) at 10 mg/mL and then activated with MBS solution (10 mg/mL in dimethyl sulfoxide) to a final concentration of 1 mg/mL. The mixture was stirred gently for 30 min at room temperature. The MBS-activated BSA was dialyzed against phosphate buffered saline (PBS) pH 7.5. Five milligrams of each peptide were dissolved in PBS + 3M guanidine hydrochloride at 5 mg/mL. The MBS-BSA solution was mixed with the peptide solution for 3 h at room temperature with gentle stirring. The conjugate was dialyzed against PBS pH 7.5 for 12 h and stored at 4 ºC. Enzyme-linked immunosorbent assay (ELISA) for titration of rabbit and mice sera All steps were carried out in a volume of 100 µL if it is not specified otherwise. Polystyrene microtiter plates (High Binding, Costar, MA, USA) were coated for 3 h at 37 °C with the recombinant protein or BSA-coupled peptides diluted to 2 mg/mL in 0.05 M carbonate-bicarbonate buffer pH 9.6. The wells were washed twice with 0.05% Tween 20 in distilled water and blocked with PBS containing 0.5% BSA (Boehringher Mannheim, Germany) (blocking solution) for 1 h at 37 °C. Serum samples were serially diluted in blocking solution containing 0.5% Tween 20 and 7.5% of goat serum (dilution buffer), added to the plates, and incubated for 1 h at 37 °C. Plates were washed again three times and anti-mouse or anti-rabbit Ig-peroxidase conjugates were added at the appropriate dilutions. For subtyping of mouse antibodies, biotinylated anti-mouse Ig of the appropriate subclass (Amersham, United Kingdom) was added at a 1/5 000 dilution, followed by a streptavidin-biotinylated horseradish peroxidase conjugated (Amersham, United Kingdom) diluted 1/16 000, and incubated for 30 min at 37 °C. Positive reactions were visualized with o-phenylenediamine in 0.1 M citric acid, 0.2 M NaH2PO4 pH 5.0 and 0.015% H2O2 as sustrate; the reaction was stopped with 50 µL of 2.5 M H2SO4. Measurement of optical densities (OD)at 492 nm was made in a MicroELISA plate reader. Interpretation of the results For TAB9 antibodies in mice, a standard curve consisting of dilutions of a pool of anti-TAB9 mouse sera was set in every ELISA plate. A value of 100 000 arbitrary units/mL was assigned to this standard serum. The values of arbitrary units in each serum were calculated from the standard curve according to the linear regression equation. For titration of rabbit and mouse sera with anti-subclass antibodies, the average OD of the corresponding dilution of the negative control sera was taken as the zero-standard. The cut-off point was set as twice the background readings. All sera were assayed twice. The maximun dilution of the serum giving OD values above the cut-off point was considered as the serum titer. Antibody titers were logarithmically transformed and the means were compared using the ANOVA test. The Spearman correlation test was employed to study the association between OD and neutralization titers. The statistical program StatWin was used. HIV-1 neutralization assay The neutralization assay was carried out as described [6]. Briefly, several dilutions of rabbit sera in RPMI 1640 medium with 10% fetal calf serum (FCS) were incubated in triplicate together with 100 TCID50 of either HIV-1/IIIB or HIV-1/MN for 1 h at 37 °C in 96-well flat-bottom microplates (Costar, Cambridge, MA, USA). A total of 1 x 105 MT4 cells were added to each well and incubated at 37 °C. After 24 h, 150 µL of culture medium were replaced with fresh medium. At the third day in culture the cells were suspended by gently pipetting, and 100 µL of the suspension were replaced by fresh medium. Viral production was estimated on day 7 by a p24 ELISA (DAVIHLAB, San José, La Habana, Cuba). Experiments were repeated three times. Results Oil-in-water emulsions induced the highest anti-TAB13 titers in rabbits In a first experiment we evaluated the adjuvant effect of several compounds on the humoral response of rabbits against TAB13. Antibody titers were clearly higher when oil adjuvants were employed. The adjuvant effect of M-ISA888 was comparable to that of other classical oil-in-water emulsions such as IFA (Figure 1). Figure 1. Antibody response against TAB13 in rabbits immunized with different adjuvants and immunomodulators. C1-C4: rabbits (1 to 4 from left to right) immunized three times with 200 µg of TAB13 in each adjuvant formulation; GMT: geometric mean of the antibody titers in each group. The bars absent for rabbit 4 in M-ISA888 and Quil A correspond to death due to toxicity of the adyuvant. The adjuvant effect of M-ISA720 on the antibody response against TAB9 in rabbits was equivalent to that of CFA and IFA Based on the results of the previous experiment and following the manufacturer suggestions, we next evaluated the effect of M-ISA720 (adjuvant formulation designed for human vaccines) on the immune response against TAB9. The titers of anti-TAB9 antibodies elicited by this adjuvant were statistically similar to those obtained with CFA or IFA (P > 0.05). Moreover, the sera were reactive against all V3 peptides included in the protein (Table 1). Table 1: Antibody responses in rabbits immunised with TAB9 in three oil-in-water adjuvants Effect of escalating doses of TAB9 in M-ISA720 Since the levels of antibodies required for protection against HIV (if any) are largely unknown, it is desirable for the vaccine candidates to be evaluated to induce optimal levels of antibodies. In the following experiment we evaluated the antibody response of rabbits immunized with increasing doses of TAB9 in M-ISA720 (Table 2). The highest antibody titers were found for the group of animals immunized with 750 µg/mL of TAB9 (300 µg/kg body weight), but no significant differences among doses could be documented (P > 0.05). However, this group also displayed the highest neutralization titers for strains MN and IIIB. A positive correlation between anti-peptide antibodies and neutralization titers was found for isolates MN (r = 0.723, P = 0.007) and IIIB (r = 0.689, P = 0.016). Table 2: Antibody response in rabbits immunized with escalating doses of TAB9 Comparative immunogenicity of TAB9 in different adjuvant preparations in mice The effect of the adjuvants on the antibody response against TAB9 was also evaluated in mice. Table 3 shows the values obtained for each experimental group. In this model, the best responses were also found for the oil adjuvants CFA, IFA and M-ISA720. Mice immunized with TAB9 in Alum or Quil A developed intermediate levels of antibodies, while they were remarkably lower in animals receiving TAB9 together with gIFN or IL-2. Table 3: Anti-TAB9 response in mice immunized with TAB9 formulated with different adjuvants Subtyping the antibody response against TAB9 The immunization of mice allowed us to determine the pattern of antibody subclass, which has been described to be indicative of the nature of the helper T cell response triggered by the immunogen. Administration of TAB9 with CFA induced the highest titers of IgG1 and IgG2a, and lower but detectable levels of IgG2b. On the other hand, IFA and M-ISA720 elicited IgG1 antibodies preferentially and remarkably lower but detectable levels of IgG2a and IgG2b. The rest of the adjuvants, Alhydrogel, gIFN, IL-2 and Quil A induced almost exclusively IgG1 antibodies (Figure 2). Figure 2. Ig class and subclass composition of the antibody response against TAB9 in mice. Mice were immunized three times intraperitoneally with 10 µg of TAB9 in different adjuvants. Discussion Recombinant DNA technology and peptide synthesis offer the potential to generate new subunit immunogens that are safer and less expensive to produce than conventional vaccines. In the case of AIDS, a safe recombinant or synthetic subunit vaccine is strongly recommended. In a previous work, we have shown that MEPs from HIV-1 V3 loop can induce broadly reactive antibodies that neutralize several laboratory isolates. However, these results were achieved formulating the protein with CFA, which is the prototype potent adjuvant [7], but unacceptable for human or even veterinary vaccines due to its severe side effects, including pain, abscess formation, local necrosis and fever. In our experiments in rabbits and mice we confirmed that CFA in particular, and other oil adjuvants that do not contain mycobacterium, as IFA and M-ISA888, promote the best humoral immune response against TAB9 and TAB13. The first experiment was conducted with TAB13 due to practical reasons, but since TAB9 showed to induce better antibody titers in rabbits [13], it was selected for the next set of experiments. Due to the high degree of identity between both MEPs, we assumed that the relative effect of the adjuvants would be similar for both proteins. The three oil adjuvants evaluated in this experiment were based on mineral oil, and their use in humans is not recommended mainly because of granuloma formation, which is considered a severe side effect for healthy individuals. We therefore evaluated M-ISA720, an adjuvant formulation recommended for human trials. M-ISA720 differs from the other oil adjuvants in that it is made of natural metabolizable oil and a highly refined emulsifier from the mannide monooleate family, with a safety profile potentially better than IFA [8]. The results demonstrated that the antibody response against TAB9 in M-ISA720 was comparable to that obtained with CFA and IFA. The antibodies generated were also widely reactive against all V3 peptides included in the protein. Even if a potent adjuvant is going to be used, it is highly desirable, especially in the case of an HIV vaccine candidate, to optimize the dose of immunogen to achieve maximal antibody titers. Four doses of TAB9 in M-ISA720 were thus explored, and although the differences among groups were not statistically significant, the higher values of geometric mean titers in ELISA and neutralizing antibodies were achieved with 750 µg. The small number of animals used in this study does not permit to conclude on this aspect. On the other hand, the results of this type of experiment can not be automatically extrapolated to other species. Therefore, several doses of immunogen should be also evaluated in primates and in phase I clinical trials in humans. Other authors have also found that M-ISA720 is an adjuvant more potent than Alum for small recombinant proteins [16, 17] and that it can even trigger a CTL response against synthetic peptides [8]. These features have made M-ISA720 an attractive option for human vaccines. There have been many speculations regarding what kind of T helper (TH) response could induce a protective response against HIV [18]. Some have pointed out that a TH1-like immune response would be more efficient to prevent HIV infection. These opinions are strongly influenced by the finding that changes from class I to class 2 response have been observed in AIDS patients with disease progression. However, this vaccine candidate is based on the assumption that a strong humoral immune response can be protective, and the achievement of a predominant TH2 response is required. On the contrary, in other vaccine concepts aimed at the induction of cytotoxic lymphocytes, a TH1 response will be clearly desirable. One of the most important features of adjuvants is their immunomodulatory capacity [19-21]. Adjuvants can stimulate the immune system and drive the response to either a TH1- or TH2-type. In a predominant TH2 response, the antibody production is maximized, especially for IgG1 secretion, while in a typical TH1 reaction the cellular immune response is enhanced and other subclasses as IgG2a and 2b are prevalent among the antibodies produced in mice. To gain insight on the TH1/TH2 balance in mice immunized with TAB9 in different adjuvants, we evaluated the subclass composition of anti-TAB9 antibodies. Animals injected with TAB-9/CFA exhibited the highest IgG2a antibody titers and lower but detectable levels of IgG2b. This capacity of CFA to induce IgG2a antibodies has been described before [22, 23] and is due to the immunomodulatory activities of the mycobacterium components in a lipid environment [24]. Other oil-in-water emulsions as IFA and M-ISA720 have been described as having a low immunomodulatory activity [19]. In this experiment, the adjuvants elicited mostly IgG1 antibodies, although IgG2a and IgG2b were also detected, which indicates that T cells are preferentially activated to a TH2 phenotype. From this evaluation it was concluded that M-ISA720 is very similar to IFA in terms of both the intensity and the nature of the immune response elicited. In this study it was also confirmed that Alum almost exclusively induces a typical TH2 response with IgG1 antibodies [19, 20]. Furthermore, the other three adjuvants gIFN, IL-2 and Quil A, induced a typical TH2 immune response because no IgG2a or IgG2b were detected. Also, the antibody titers were the lowest with these adjuvants. Several authors have demonstrated a positive immunomodulatory effect of IL-2 and gIFN [10, 19, 25], although these adjuvants have been reported to induce primarily a TH1 response. This apparent contradiction could be clarified by quantifying the levels of TH1 and TH2 lymphokines in serum. On the other hand, Quil A have been reported to induce a TH2 cytokine pattern [26]. Additionally, our results indicate that neither IL-2 nor gIFN improved significantly the effect of Alum on the antibody response in rabbits. However, the co-administration of IL-2 has been reported to increase the antibody response against several immunogens in different species [11, 27], and murine gIFN has also enhanced the antibody response against influenza virus in mice [28]. The latter authors were not able to find a positive effect of human IL-2 in the antibody response against influenza in mice. Taken together, these results suggest that the effect of IL-2 and gIFN on the antibody response varies depending on the antigen, the species, and most probably on other factors as the dose and route of immunization used, and consequently, it should be studied on an individual basis. From these experiments M-ISA720 was selected for further testing of MEPs as AIDS vaccine candidates in non-human primates and clinical trials. Refrerences 1. Centers for Disease Control. Pneumocystis pneumonia- Los Angeles. MMWR 1981;30:250-2. 2. Verani P, Buttó S, Taddeo B, Federico M, Rossi GB. HIV variability and perspectives for a vaccine. Vaccine 1993;11(5): 542-5. 3. Alving CR, Detrick B, Richards RL, Lewis MG, Shafferman A, Eddy GA. Novel adjuvant strategies for experimental malaria and AIDS vaccines. Ann N Y Acad Sci 1993;690:265-75. 4. Nardelli B, Lu Y, Shiu DR, Delpierre-Defoort C, Profy AT, Tam JP. A chemically defined synthetic vaccine model for HIV-1. J Immunol 1992;148:914-20. 5. Duarte CA, Montero M, Seralena A, Valdés R, Jiménez V, Benítez J, et al. Multi-epitope polypeptide (MEP) containing epitopes of HIV-1 envelope induces neutralizing monoclonal antibodies against the V3 loop. AIDS Res Hum Retrovirus 1994;10(3):235-43. 6. Montero M, Menéndez A, Domínguez MC, Navea L, Vilarubia OL, Quintana D, et al. Broadly reactive antibodies against a gp120 V3-loop multi-epitope polypeptide neutralize different isolates of Human Immunodeficiency Virus type 1 (HIV-1). Vaccine 1997;15(11):1200-8. 7. Freund J. The mode of action of immunologic adjuvants. Adv Tuberc Res 1956; 7:130. 8. Scalzo AA, Elliott SL, Cox J, Gardner J, Moss DJ, Suhrbier A. Induction of protective cytotoxic T cells to murine cytomegalovirus by using a nonapeptide and a human-compatible adjuvant (Montanide ISA720). J Virol 1995;69:1306-9. 9. Moncef Slaoui. Recent advances with new vaccine adjuvants: from pre-clinical to clinical development. In: New trends in vaccine R and D: adjuvants, delivery systems and antigen formulations. Programme and Abstracts; 1998 Feb 26-28; Paris, France. 10. Heath AW, Playfair JHL. Cytokines as immunological adjuvants. Vaccine 1992; 10:427-34. 11. Numberg JH, Doyle MV, York SM, York CI. Interleukin-2 acts as an adjuvant to increase the potency of inactivated rabies virus vaccine. Proc Natl Acad Sci 1986; 86:4240-3. 12. Myers G, Korber B, Wain-Hobson S, Smith R, Pavlakis GN, editors. Human retroviruses and AIDS. A compilation and analysis of nucleic acid and amino acid sequences. Los Alamos (New Mexico): Los Alamos National Laboratory Press; 1994. 13. Menéndez A, Montero M, Quintana D, Gómez C, Navea L, Vilarubia OL, et al. V3 multi-epitope polypeptides: the number of V3 regions affects the exposition of epitopes and immunogenicity [abstract]. In: Sussel R, Guillet V, Ruedy N, editors. Abstracts 1. XI International Conference on AIDS; 1996 July 7-12; Vancouver, Canada. Vancouver: WordPower Business Services; 1996. p.212. Abstract nr Tu-A.2104. 14. Laemmli UK. Cleavage of structural proteins during the assembly of the head of the bacteriophage T4. Nature 1970; 227:680-5. 15. Bradford MM. A rapid and sensitive method for quantitation of microgram quantities of proteins utilizing the principle of protein-dye binding. Anal Biochem 1976;72:248. 16. Pye D, Vandenberg KL, Dyer SL, Irving DO, Goss NH, Woodrow GC, et al. Selection of and adjuvant for vaccination with the malaria antigen, M-ISA720. Vaccine 1997;15(9):1017-23. 17. Lawrence GW, Saul A, Giddy AJ, Kemp R, Pye D. Phase I trial in humans of an oil based adjuvant SEPPIC Montanide ISA720. Vaccine 1997;15(2):176-8. 18. Clerici M, Sheare GM. Correlates of protection in HIV infection and the progression of HIV infection to AIDS. Immunology Letters 1996;51:69-73. 19. Cox JC, Coulter AR. Adjuvants: a classification and review of their modes of action. Vaccine 1997;15 (3):248-56. 20. Cooper PD. The selective induction of different immune responses by vaccine adjuvants. In: Strategies in vaccine design, (Ada GL, editor) RG Landes Company, 1994,125-58. 21. Bomford R, Stapleton M, Winsor S, MckNight, Andronova T. The control of the antibody isotype response to recombinant Human Immunodeficiency Virus gp120 antigen by adjuvants. AIDS Res Human Retroviruses 1992;8(10):1765-71. 22. Kenney JS, Hughes BW, Masada MPl. Influence of adjuvants on the quantity, affinity isotype and epitope specificity of murine antibodies. J Immunol Meth 1989; 121:157-66. 23. Kensil CR, Patel U, Lennick M, Marciani D. Separation and characterization of saponins with adjuvant activity from Quilaria saponaria Molina cortex. J Immunol 1991;146:431-7. 24. Parant MA, Audibert FM, Chedid LA. Immunostimulatory activities of a lipophilic muramyl dipeptide derivative and of desmuramyl peptidolipid analogs. Infect Immun 1980;27:826-31. 25. Vogel FR, Powell MF. A compendium of vaccine adjuvants and exhipients. In: Powell MF, Newman MJ, editors. Vaccine Design. The Subunit and Adjuvant Approach. New York: Plenum Press; 1995. p.141-228. 26. Victoratos P, Yiangou M, Avramidis N, Hadjipetrou L. Regulation of cytokine gene expression by adjuvants in vivo. Clin Exp Immunol 1997;109:569-78. 27. Hughes HPA, Campos M, van Drunen S, Littel van den Hurk, Zamb T, Sordillo LM, et al. Multiple administration of interleukin-2 potenciates antigen-specific responses to subunit vaccination with bovine herpesvirus-1 glycoprotein IV. Vaccine 1992;10(4):226-30. 28. Cao M, Sasaki O, Yamada A, Imanishi J. Enhancement of the protective effect of inactivated influenza virus vaccine by cytokines. Vaccine 1992;10(4):238-42. Received in September, 1998. Accepted for publication in December, 1998. Copyright 1999 Elfos Scientiae The following images related to this document are available:Photo images[ba99018e.jpg] [ba99018b.jpg] [ba99018a.jpg] [ba99018c.jpg] [ba99018d.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}