 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnologia Aplicada 1999; Vol. 16 No. 3, pp. 145-148 Production of Recombinant Treponema pallidum Membrane Protein A in Escherichia coli Using Fed-batch Fermentation Juan M Rivera, María del Carmen Domínguez, Maira Ponce, Ramón E Narciandi División de Inmunotecnología y Diagnóstico.
Centro de Ingeniería Genética y Biotecnología.
Received in February, 1999. Accepted for publication in May, 1999. Code Number: BA99024 ABSTRACT There is special interest on the 42-kDa Treponema pallidum membrane protein A (TmpA) due to its potential use for syphilis serological assay. Different genetic constructions and cloning in bacteria of recombinant DNA containing the synthetic TmpA gene have been reported, but in all cases the expression levels and fermentation productivity have been affected by different factors and, on the other hand, its purification has involved a series of laborious steps. A fed-batch culture of Escherichia coli W3110 capable of achieving high expression levels (36%) and high production yields (1.74 g/L) of recombinant TmpA was developed using the expression vector pR2M6. Different host strains and induction conditions were studied in order to maximize protein expression. In this study, a rapid and simple procedure for the purification of recombinant TmpA to over 85% of purity and about 60% of recovery is reported using immobilized metal ion affinity chromatography. Keywords: Escherichia coli, fed-batch fermentation, recombinant protein, TmpA RESUMEN La proteína de membrana A de 42 kDa de Treponema pallidum (TmpA) es de especial interés debido a sus potencialidades para el diagnóstico serológico de la sífilis. Se han reportado diferentes construcciones genéticas y la clonación en bacterias de fragmentos de ADN que contienen el gen sintético de TmpA. Sin embargo, en todos los casos los niveles de expresión y la productividad de la fermentación han estado afectados por diferentes factores y, por otro lado, la purificación se ha realizado mediante pasos complejos. Para resolver estos inconvenientes, se desarrolló un cultivo incrementado de Escherichia coli W3110 que permite obtener altos niveles de expresión (36%) y un rendimiento elevado (1,74 g/L) de TmpA recombinante mediante la utilización del vector de expresión pR2M6. Se estudiaron diferentes cepas y condiciones de inducción con el objetivo de maximizar los niveles de expresión de la proteína. En este trabajo se reporta un procedimiento rápido y sencillo para la purificación de TmpA recombinante con más de 85% de pureza y alrededor de 60% de recobrado, mediante cromatografía de de afinidad por iones metálicos. Palabras claves: cultivo incrementado, Escherichia coli, proteína recombinante, TmpA Introduction Among the treponemal antigens that have been expressed by recombinant DNA techniques, the 42 kDa Treponema pallidum membrane protein A (TmpA) appears to be a promising candidate for syphilis diagnosis due to the potential sensitivity and specificity that can be achieved in syphilis mass screening. TmpA is also considered a good marker for treatment recovery [1, 2]. Originally, cloning and expression of TmpA were reported using different plasmid constructions in which gene expression was under the control of the l pL promoter [3]. However, the bacteria containing these plasmids were unable to overexpress TmpA. Later, the construction of an expression plasmid carrying two T. pallidum genes encoding the 42-kDa TmpA and the 34-kDa TmpB was described, in which the protein was expressed to an insoluble form associated with the plasma membrane [4]. In that work, an expression level of approximately 15% of total proteins and a fermentation yield of about 6.5 g/L (wet weight) using fed-batch culture, were reported. The purification of this protein involved a series of very complex steps. The authors of the present work have designed and cloned in Escherichia coli a gene construction allowing a soluble variant of a recombinant TmpA to be produced using the pR2M6 vector [5]. Also, the capacity of this system to overproduce recombinant TmpA in a fed-batch fermentation process was analyzed. Different host strains and induction conditions were studied in order to maximize gene expression, and a rapid and simple procedure for the purification of TmpA by a one-step Ni2+-IDA affinity chromatography was used. Materials and Methods Bacterial strains and plasmid E. coli strains W3110, C600, HB101 and GC366 were used as host for the plasmid pR2M6, a derivative of pPF-15 [6], which harbors a DNA sequence for the expression of TmpA gene under the control of the trp promoter and fused to a Hisx6 coding tag in its N-terminus for purification by Ni2+-IDA affinity chromatography. Selection of the host strain The aforementioned E. coli strains transformed with the plasmid pR2M6 were inoculated into 5 mL of M9 medium (6 g/L Na2HPO4, 3 g/L KH2PO4, 1 g/L NH4Cl, 0.5 g/L NaCl, 2 mM MgSO4, 10mM CaCl2) supplemented with casamino acids (10 g/L), glucose (10g/L), tryptophan (10 m g/mL), and ampicillin (50 m g/mL), and incubated at 37 ºC for 2 h. After this time, 3b -indoleacrylic acid (b IA) (40 m g/mL) was added and the cultures were incubated for 8 h. The bacterial pellets were tested for protein expression by densitometric analysis of 12.5% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) [7]. Shake flask experiments W3110 cells were grown for 8 h at 37 ºC in 50 mL of Luria Bertani (LB) medium (10 g/L tryptone, 5 g/L yeast extract, 10 g/L NaCl) suplemented with ampicillin (50 m g/L) and tryptophan (100 m g/mL). For each experiment, 1 mL of culture was inoculated into 50 mL of M9 medium supplemented with casamino acids (10 g/L), glucose (10 g/L) and ampicillin (50 m g/mL), and incubated with shaking at 37 ºC for 12 h. The expression of the TmpA gene was studied using an initial tryptophan concentration in the medium ranging from 0 to 100 m g/mL in the absence of b IA. The effect of the inductor concentration was determined by adding 20 g/mL of tryptophan to the initial culture medium and 0, 20, 40, 60 or 100 mg/mL of b IA when the culture optical density (OD530) reached 1. Samples were taken every 4 h of cultivation. Fed-batch media composition and culture conditions Precultures were grown overnight in shake flasks containing LB medium (50 mL) supplemented with tryptophan (100 m g/mL) and ampicillin (50 m g/mL) at 37 ºC. Fed-batch cultures were performed in a 2-L jar fermentor (B.E. Marubishi, Japan). The initial culture M9 medium (1.3 L) contained glucose (5 g/L), yeast extract (1 g/L), tryptone (5 g/L), tryptophan (100 m g/L), and ampicillin (50 m g/mL). Fermentation was carried out at pH 7, with an air flow of 1 vvm and a stirring speed of 580 rpm, at 37 ºC. The addition of the feeding solution (125 mL), composed of M9 medium, glucose (20 g/L) and yeast extract (15 g/L), was started after 2 h of fermentation. TmpA gene expression was induced by depletion of tryptophan in the medium. Analytical methods Cell growth was followed by cell dry weight determination. Expression levels were determined by 12.5% SDS-PAGE of the samples taken from the cultures followed by quantitation in an LKB Bromma 2202 Ultrascan laser densitometer (LKB, Sweden). From this analysis, the percentage of recombinant protein produced from total cellular protein was calculated. The total cellular protein was determined using the method reported by Lowry et al. [8], using bovine serum albumin as standard. For Western blot analysis, the proteins were transferred from 12.5% polyacrylamide gels to 0.45 mm nitrocellulose membranes in a semi-dry transfer system (Bio-Rad, USA). The membrane was blocked with 2% powdered milk in phosphate-buffered saline (PBS) for 1 h at 37 ºC, and incubated for 1 h at the same temperature with a 1:20 dilution of anti-T. pallidum serum from a syphilis patient (pre-adsorbed to E. coli proteins). After three washings with PBS containing 0.05% Tween 20, the membrane was incubated with a 1:1000 rabbit anti-mouse IgG antibody conjugated with horseradish peroxidase (HRPO) (Sigma, USA) for 30 min at 37 °C. The reaction was developed with 0.5 mg/mL of 3,3-diaminebenzidine (Sigma, USA) and 0.015% of hydrogen peroxide. Protein purification One gram of harvested cells was lysed by sonication in 10 mL of phosphate buffer pH 8 containing 8 M urea. Cell debris was removed by centrifugation at 4 ºC and the supernatant was loaded onto a Fractogel EMD Chelate 650(S) column (Merck, Germany) equilibrated in the same buffer. The matrix was washed with phosphate buffer using a descending pH gradient (8 to 6). The adsorbed proteins were eluted in phosphate buffer pH 3.5. Results and Discussion Influence of the host strain on protein expression levels A strong host strain dependence of protein expression and degradation was observed. The highest expression levels were achieved using W3110 (30%). For C600 (15%), GC366 (12%) and HB101 (5%) a very high proteolytic activity was found when the samples were analyzed by Western blot, which is the fundamental cause of the difference in the expression levels achieved for each of the strains. Protein degradation was enhanced in HB101 and very slight in W3110 strain. In all cases, TmpA was produced as an intracellular soluble protein. In order to continue the shake flask experiments as well as the development of fed-batch cultivation, W3110 strain was selected. Optimization of the induction conditions Different concentrations of tryptophan and b IA were used in order to optimize induction conditions. Tables 1 and 2 summarize the results obtained. Table 1. Effect of tryptophan concentration on cell growth, expression level and production of TmpA.
Table 2. Effect of b IA concentration on cell growth, expression level and production of TmpA.
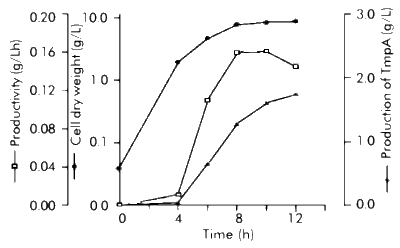
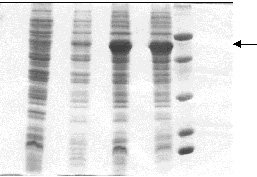
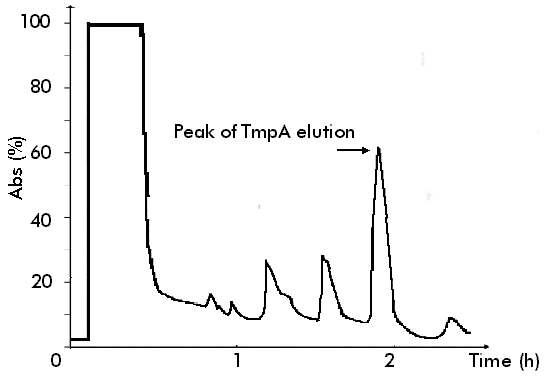
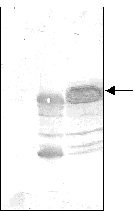
It was found that the amount of synthesized TmpA decreased, while tryptophan concentration increased. High protein expression levels were observed even without b IA addition when low tryptophan concentrations (0-40 m g/mL) were used, and gene expression could only be maintained at basal levels using high tryptophan concentrations (60-100 m g/mL). In spite of being one of the most commonly employed methods to obtain an adequate induction of the trp promoter [9], the addition of b IA was unnecessary to achieve protein expression because the best results were obtained by trp promoter derepression. On the contrary, a considerable reduction of protein production and cellular growth was observed while b IA concentration increased, due to the fact that b IA induces tryptophan starvation. Different groups have previously reported similar results after studying the expression of recombinant proteins using the trp promoter [10-12]. Awareness of these details is important when fed-batch culture is developed because timing control of recombinant gene expression is often advantageous, since overproduction of recombinant proteins is deleterious or even fatal for cell growth and overall protein synthesis in the host [13]. Fed-batch cultivation Fed-batch culture is an efficient method for culturing microorganisms to achieve a high biomass production and large amounts of heterologous protein [10]. However, it has been previously demonstrated [14, 15] that optimal expression and product stabilty resulting from the combination of effects (i.e. medium composition, induction strategy) may not be predicted, because it also depends upon host strain and plasmid construction and may be achieved using widely different fermentation conditions. The results obtained from shake flask experiments were applied in the development of a single-stage fed-batch culture in which glucose was added at a feeding rate of 3.36 g/Lh. The authors previously reported [16] that this feeding flow permits the production of large amounts of recombinant protein because glucose starvation is avoided, which allows, at the same time, to maintain low concentrations of glucose during the fermentation process (this is very beneficial for cell growth). Figure 1 shows the fermentation kinetics of a fed-batch culture producing TmpA recombinant protein. Two well-defined phases can be observed. In the first (until 4 h of cultivation), the cells grow exponentially at a specific rate of 0.98 h-1 and gene expression is almost repressed. In the second (within 4-12 h of cultivation), the cells begin to produce the recombinant protein due to tryptophan depletion in the culture medium and the specific growth rate drastically decreases. Higher cellular density and TmpA production was achieved after 12 h of cultivation, with the obtention of 8.8 g of cell dry weight/L and 1.74 g/L of recombinant protein. Figure 1. Fermentation kinetics of E. coli W3110 producing recombinant TmpA. Fed-batch cultures were supplemented in M9 medium supplemented with glucose (5 g/L), yeast extract (1 g/L), triptone (5 g/L), tryptophan (100 m g/mL) and ampicillin (50 m g/mL). Fermentation was carried out at 37 ºC, pH 7, 1 vvm and 580 rpm. The addition of the feeding solution (125 mL) was started after 2 h of fermentation. TmpA gene expression was induced by depletion of the initial tryptophan concentration present in the medium. Schouls et al. [4] reported the use of fed-batch culture for TmpA production under the control of the l pL promoter, in which glucose was added during cultivation at a rate sufficient to maintain 1 g/L. They obtained about 6.5 g/L of cell wet weight (approximately 1.6 g/L of cell dry weight) and a production of 120-150 mg/L of recombinant protein. In terms of growth and production, and from the comparison of both results, the process developed by our group represents increases in cell weight and recombinant protein production of at least 5.5 and 11.6 times, respectively. A larger concentration of tryptophan (200 m g/mL and 400 m g/mL) was added to the initial culture medium, with the objective of delaying the start of recombinant protein expression and lengthening the exponential growth phase. However, the results were the same, which could be due to the induction of tryptophanase expression and therefore a rapid depletion of tryptophan by the low glucose concentrations maintained during the fermentation process [17]. SDS-PAGE of samples taken every 4 h during the process is shown in Figure 2. A high expression level of TmpA was observed at 8 h of cultivation, reaching the maximum expression at 12 h (36%). This result was very similar to that obtained in shake flask culture with better induction conditions. In terms of expression level of recombinant TmpA, the fed-batch process developed represents an increase of 2.4 times, compared to that previously published in the literature. Figure 2. SDS-PAGE in 12.5% gels of samples taken during fed-batch fermentation of E. coli producing recombinant TmpA. Lanes 1-4, cultivation for 0, 4, 8 and 12 h; lane 5, molecular weight marker: 67, 41, 25, 18 and 14 kDa from top to bottom. The arrow indicates the band corresponding to recombinant TmpA. TmpA purification The purification procedure for recombinant TmpA reported by other authors [2, 4] consists of a series of complex steps such as extraction from membrane, ion exchange chromatography and electroelution, which makes the process very expensive with an overall yield of 3 mg/g of cell wet weight. In this study, TmpA could be easily purified using a one-step Ni2+-IDA affinity chromatography. Figure 3 shows the characteristic chromatogram of the purification process. About 60% of TmpA was recovered in the elution peak at pH 3.5, with a yield of around 15 mg/g of cell wet weight. Protein purity was higher than 85% if the degradation observed in Western blot is taken into account (Figure 4). Figure 3. Characteristic chromatogram of TmpA separation using Fractogel EMD Chelate 650(S). The cells (1 g) were lysed by sonication in phosphate buffer pH 8 containing 8 M urea (10 mL). The cell debris was removed by centrifugation at 4 oC and the supernatant was loaded onto the column. After different washes using a descending pH gradient (8 to 6), TmpA was eluted at pH 3.5. Figure 4. Western blot assay using serum sample from a patient with early latent syphilis. Lane 1, total protein from E. coli W3110 strain transformed with the pR2M6 plasmid lacking the TmpA gene; lane 2, recombinant TmpA purified by Ni2+-IDA affinity chromatography; lane 3, total protein from E. coli W3110 strain transformed with the pR2M6 plasmid carrying the TmpA gene (sample of 12 h of fed-batch fermentation). The arrow indicates the band corresponding to recombinant TmpA. Conclusions The expression of TmpA is highly dependent on the host strain and the induction conditions. The fed-batch process developed and the purification procedure using Ni2+-IDA affinity chromatography, permit the production of large amounts of TmpA with a high purity. Acknowledgments The authors would like to thank Drs Eduardo Pentón and Lidia I. Novoa for carefully reading the manuscript. They also thank Raudel Sosa and Daniel Rodríguez for technical assistance during stages of this work. References 1. Ijsselmuiden OE, Schouls LM, Stolz E, Aelbers GNM,
Agterberg CM, Top JM, et al. Sensitivity and specificity of
an enzyme-linked immunosorbent assay using the recombinant
DNA-derived Treponema pallidum protein TmpA for
serodiagnosis of syphilis and the potential use of TmpA for
assessing the effect of antibiotic therapy. J Clin Microbiol 1989;
27:152-7. Copyright 1999 Elfos Scientiae The following images related to this document are available:Photo images[ba99024d.jpg] [ba99024b.jpg]Line drawing images[ba99024a.gif] [ba99024c.gif] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}