 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Comparación de la quimiosina bovina secretada al medio de cultivo por cepas recombinantes de las levaduras Pichia pastoris y Kluyveromyces lactis Ángela E Sosa Espinosa,1 Jorge Sotolongo
Peña,2 Vladimir Martínez Santana,3 1Subdirección de Regulaciones, 2División de
Formulación y Envase, 3División de Desarrollo,
4División de Bioindustria, 5División de Vacunas. Recibido en mayo de 1998. Aprobado en diciembre de 1998. Code Number: BA99027
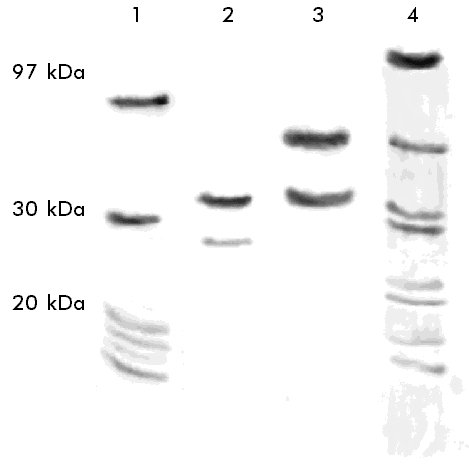
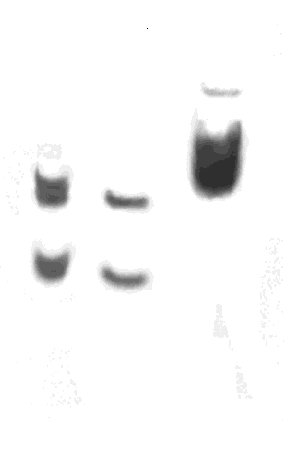
ABSTRACT Bovine chymosin, an aspartic protease that is used for the industrial production of cheese, is extracted from the stomachs of calves. To be obtained from another source, chymosin has been cloned in different microorganisms. However, the post-translational modifications of the recombinant enzyme that frequently occur in the host cells may change its biochemical properties. In this work, the properties of a raw material which contains the bovine chymosin produced in recombinant strains of the yeasts Pichia pastoris or Kluyveromyces lactis are studied. Recombinant chymosin glycosilation was analyzed by the periodic acid-Schiff reaction on polyacrylamide slab gels. Besides, the effect of several parameters (the molar concentration of Ca2+ ions, pH, temperature) on the chymosin coagulant activity and the proteolytic activity and the thermal stability of the enzyme in solution, were studied. In comparison with the yeast K. lactis, it was observed that P. pastoris can be a better host for the expression of chymosin, since the properties of the P. pastoris-derived enzyme seemed to be closer to those of the natural enzyme. Keywords: Kluyveromyces lactis, Pichia pastoris, recombinant chymosin RESUMEN La quimiosina bovina, una proteasa aspártico-dependiente que se utiliza para la producción industrial de quesos, se extrae de los estómagos de terneros lactantes. Para obtenerla de otra fuente, la quimiosina ha sido clonada en varios microorganismos. Sin embargo, las modificaciones postraduccionales de la enzima recombinante que ocurren con frecuencia en la célula hospedera, pueden cambiar sus propiedades bioquímicas. En este trabajo se estudian las propiedades de un crudo que contiene la quimiosina bovina producida en cepas recombinantes de las levaduras Pichia pastoris y Kluyveromyces lactis. La glicosilación de la quimiosina recombinante se analizó mediante la reacción de ácido peryódico-Schiff en geles laminares de poliacrilamida. Además, se estudió el efecto de varios parámetros (concentración molar de iones Ca2+, pH y temperatura) sobre la actividad coagulante de la quimiosina, la actividad proteolítica y la termoestabilidad de la enzima en solución. En comparación con la levadura K. lactis, se observó que P. pastoris puede ser un hospedero más adecuado para la expresión de la quimiosina, ya que las propiedades de la enzima derivada de esta última fueron más cercanas a las de la enzima natural. Palabras claves: Kluyveromyces lactis, Pichia pastoris, quimiosina recombinante Introducción La quimiosina bovina es una proteasa aspártica que se utiliza en la industria del queso. Las células de la mucosa gástrica de terneros lactantes secretan esta enzima en forma de un zimógeno inactivo de 40 777 Da (365 aminoácidos, aa). La activación del zimógeno ocurre por autoproteólisis a pH ácido, que después de la liberación de un péptido de 42 aa, rinde la forma activa de la enzima, de 323 aa y un peso molecular de 35 600 Da [1]. La quimiosina bovina que se obtiene de su fuente natural no es modificada, aunque presenta dos sitios potenciales de glicosilación [2, 3]. En la industria del queso esta enzima ha sido utilizada tradicionalmente en la coagulación de la leche, lo que ha hecho difícil su sustitución sin que se afecte la calidad de muchas variedades de queso. Como una posible solución para obtener una molécula similar a la quimiosina natural, se propuso la clonación del gen que codifica la quimiosina B mediante las técnicas de ADN recombinante. Sin embargo, las diferencias de los hospederos en relación con las células de la mucosa gástrica, condicionan diversos cambios bioquímicos en la enzima recombinante que hacen que difiera de la natural. En el caso de Escherichia coli [4] y la levadura Saccharomyces cerevisiae [5], no se obtiene un preparado industrialmente deseable. En Kluyveromyces lactis, 90% de la enzima obtenida es activable y no se han informado diferencias con la enzima que se obtiene de la fuente natural, sin embargo, no es utilizable en todas las variedades de queso [6]. En Pichia pastoris [7], la cepa recombinante que se desarrolló en nuestro laboratorio secreta al medio de cultivo grandes cantidades de quimiosina B, la cual es caracterizada en este trabajo. Hasta el momento, no se conocen las consecuencias de las posibles modificaciones que ocurren en la proteína al ser expresada en esta levadura. Con el objetivo de evaluar la enzima secretada por una cepa recombinante de la levadura P. pastoris, en el presente trabajo se caracteriza un crudo enzimático proveniente del sobrenadante de cultivo de esta levadura, y se compara con el que se obtiene por la misma vía en una cepa recombinante de K. lactis que, hasta el momento, se considera el hospedero más adecuado para la expresión de esta proteína. Los dos crudos enzimáticos se compararon en cuanto a: a) el efecto de varios parámetros (concentración molar de iones Ca2+, pH y temperatura) sobre la actividad coagulante de la quimiosina, y b) la actividad proteolítica y la termoestabilidad de la enzima en solución. Se estudiaron, además, los niveles de glicosilación, con la inclusión de la enzima natural en todos los experimentos como criterio de los parámetros óptimos. Materiales y Métodos Microorganismos y material biológico Cepas MP-36(pRPC-316) de P. pastoris [7] y la cepa CBS683(pKM1-130) de K. lactis [8]. La quimiosina bovina de la casa comercial Sigma Chemical Co. (St. Louis, MO) se utilizó como patrón en todas las comparaciones. Para la obtención de anticuerpos policlonales específicos contra la quimiosina, se utilizaron conejos albinos de Nueva Zelanda. Experimentos en zaranda Las cepas se crecieron en medio YP (extracto de levadura 1%, peptona bacteriológica 2%) con diferentes fuentes de carbono (glucosa 2%, glicerol 2%, lactosa 2%, galactosa 2% o metanol 1%). Los cultivos se inocularon a partir de un precultivo con una absorbancia de 0,05 DO600 y se crecieron a 30 ºC con una agitación de 250 rpm durante 120 h, en una zaranda New Bronswick Scientific (NJ, Estados Unidos). Cada 12 h se tomaron muestras de 1 mL del cultivo y se les midió la absorbancia. Pasado este período, las células se colectaron por centrifugación a 3 000 rpm (Himac Centrifuge, HITACHIâ, Japón), el sobrenadante se separó en alícuotas y se congeló hasta su uso. Fermentación Las levaduras P. pastoris y K. lactis se inocularon a partir de muestras conservadas en glicerol a -20 ºC, en erlenmeyers de 2 L con 500 mL de medio de cultivo: (NH4)2SO4 2 g/L, NH4H2PO4 6 g/L, NaCl 0,5 g/L, CaCl2 anhidro 0,1 g/L, KH2PO4 0,1 g/L, y una solución de sales trazas y vitaminas que contiene CuSO4•5H2O 2,1 mg/L, MnSO4 11 m g/L, NaMoO4• 2H2O 11 m g/L, H3BO3 11 mg/L, ZnSO4•7H2O 70 mg/L, FeSO4•7H2O 11 mg/L, KI 42 mg/L, inositol 2,1 mg/L, tiamina 2 mg/L, piridoxina 1,1 mg/L, ácido nicotínico 1,1 mg/L, biotina 2,1 mg/L y pantotenato de calcio 2 mg/L, y se crecieron por 24 h con una agitación de 250 rpm a 30 ºC. Las cepas se cultivaron por lote incrementado en fermentadores de 5 L modelo BE Marubishi MD-300 con 3 L de volumen efectivo. En las fermentaciones de K. lactis se utilizó el mismo medio, excepto que se adicionó (NH4)2SO4 3,2 g/L, sales trazas tres veces más concentradas y extracto de levadura 5 g/L. En el caso de P. pastoris, el medio no contenía calcio y sólo se utilizaron 22 g de (NH4)2SO4 como fuente de nitrógeno. En ambas fermentaciones se utilizó glicerol 2% como fuente de carbono durante las primeras 12 h de cultivo; en el tiempo de fermentación restante, la fuente de carbono inductora de la expresión génica (lactosa en el caso de K. lactis y metanol para P. pastoris) se mantuvo a 1%, por adición a flujo constante. Los cultivos se crecieron con un régimen de agitación de 600 rpm, una aireación de 1 vvm, la temperatura se mantuvo entre 28 y 30 ºC y el pH entre 5 y 5,5 por adición de NH4OH. Durante las primeras 24 h, se extrajeron muestras cada 4 h y después las extracciones se realizaron cada 12 h. Al final de la fermentación (80 h para K. lactis y 120 h para P. pastoris) se centrifugó el cultivo, el sobrenadante se concentró cinco veces por diafiltración (AMICON® 8400 de 350 mL) contra una solución tampón PBS, se dosificó y se congeló a -20 ºC hasta su análisis. Caracterización de las muestras Las muestras se descongelaron a temperatura ambiente y se activaron según el procedimiento de Pedersen y colaboradores [9]. La actividad coagulante se determinó como lo describió Foltmann [1]. Una unidad de actividad se definió como la cantidad de enzima que coaguló 10 mL de una solución que contiene leche en polvo semidescremada 12% y CaCl2 10 mM, a pH 7 y 30 ºC en 100 s. La concentración de proteínas totales en el sobrenadante se midió por el método de Lowry [10]. Se hicieron tres réplicas por cada experimento. La influencia de la temperatura (30, 40 y 50 ºC) sobre la actividad enzimática, se determinó por medición de la actividad coagulante de las enzimas provenientes de cada hospedero. El efecto del pH sobre la actividad específica se determinó por medición de la actividad coagulante de las enzimas provenientes de cada hospedero, a valores de pH de reacción de 6; 6,4 y 6,8 ajustados con HCl 1M. La influencia de la concentración de iones Ca2+ sobre la actividad específica de la quimiosina, se determinó por medición de la actividad coagulante de las enzimas provenientes de cada hospedero a una concentración de 0, 2, 4 y 8 mM de CaCl2 en la mezcla de reacción. La actividad proteolítica de las enzimas provenientes de cada hospedero, se midió según lo describieron Arima y colaboradores [11]. La estabilidad térmica de los crudos provenientes de cada hospedero, se determinó por medición de la actividad coagulante después de incubar nuevamente los crudos enzimáticos a las temperaturas 30, 40 y 50 ºC por un intervalo de 0 a 20 min. El grado de glicosilación de las enzimas provenientes de cada hospedero se determinó semicuantitativamente por el método de Hsi y colaboradores [12]. Se aplicaron 20 m g de albúmina de suero bovino (Sigma, St. Louis, MO), de crudo proveniente de P. pastoris, de crudo proveniente de K. lactis, y de amilasa comercial (Sigma, St. Louis, MO), en una membrana de nitrocelulosa. La membrana se incubó 1 h a 42 ºC en una solución que contenía leche descremada 5% en solución tampón TBS (Tris 50 mM, NaCl 150 mM, pH 8), posteriormente se incubó a temperatura ambiente en una solución que contenía un conjugado concanavalina A-peroxidasa (Sigma, St. Louis, MO). Después de este tiempo, se lavó tres veces con solución tampón TBS, una vez con TBS que contenía Tween 20 0,05% y un último lavado con TBS. Posteriormente, se adicionó una solución que contenía 2 mg de 4-cloro-1-naftol, 600 m L de metanol, 10 mL de TBS y 10 m L de peróxido de hidrógeno 30%, y se incubó a temperatura ambiente hasta la aparición de la señal. Los crudos enzimáticos provenientes de cada hospedero fueron comparados por electroforesis desnaturalizante en un gel de poliacrilamida a 12,5% [13]. Después de la electroforesis, las proteínas se detectaron con el colorante azul Coomassie G-250 (Sigma, St. Louis, MO), con el reactivo de Schiff, o se transfirieron a una membrana de nitrocelulosa (Scheleider & Schuell, Alemania) mediante la técnica de Western blot [14]. La cantidad de quimiosina presente en los sobrenadantes de cultivo de las fermentaciones, se determinó mediante Dot blot: se puntearon 10 m g de proteínas totales de concentración conocida sobre un filtro de nitrocelulosa (Scheleider & Schuell, Alemania), el cual se incubó durante 3 h en 10 mL de una solución de leche descremada 5% preparada en tampón TBS. Después, se añadieron 200 m L de suero de un conejo inmunizado con la quimiosina comercial (Sigma, St. Louis, MO) y se incubó 2 h a temperatura ambiente. El exceso de anticuerpos se eliminó mediante dos lavados en 25 mL de TBS, un lavado en 25 mL de TBS con NP-40 y un lavado en 25 mL de TBS. Después de los lavados, se transfirió el filtro a una solución de TBS con I125, se incubó 1 h, se lavó en 25 mL de TBS y se expuso a una lámina de autorradiografía durante 8 h a -70 ºC. Resultados y Discusión Las levaduras K. lactis y P. pastoris son capaces de generar una gran cantidad de biomasa en medios salinos, y de secretar proteínas extracelulares foráneas cuyos genes han sido integrados a su genoma. Al realizarse la fermentación en un medio salino de composición similar, el máximo de crecimiento de la cepa recombinante de K. lactis que expresaba la quimiosina bovina se obtuvo a las 80 h con un valor de peso húmedo de 250 g/L. Esta cantidad de biomasa en la cepa recombinante de P. pastoris se alcanzó a las 120 h de cultivo. Al final de la fermentación, la proquimiosina en el sobrenadante se activó por disminución del pH. En un estudio cinético (Tabla), se observó que la proquimiosina secretada por P. pastoris se activó totalmente durante la fermentación, lo cual no ocurrió con la secretada por K. lactis. La quimiosina de K. lactis permaneció inactiva durante todo el tiempo de cultivo (80 h), por lo que requirió condiciones más prolongadas de activación a pH 2,5 antes de su utilización. Tabla. Cinética de activación de la quimiosina bovina contenida en los sobrenadantes de cultivos de P. pastoris y K.lactis.
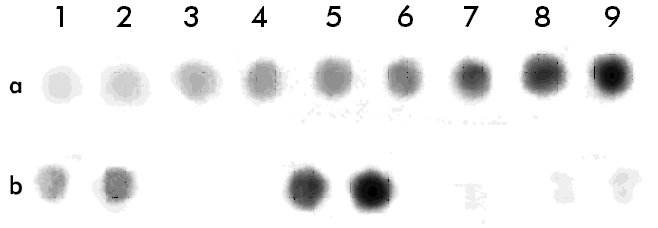
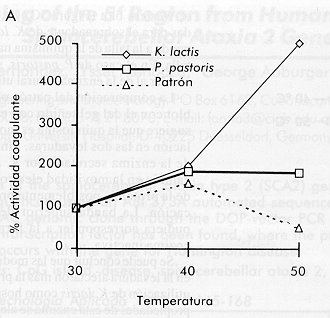
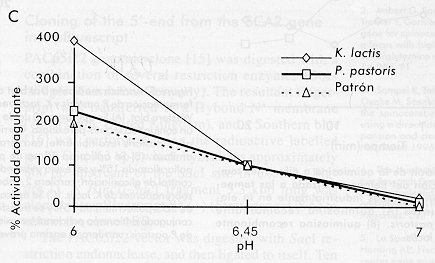
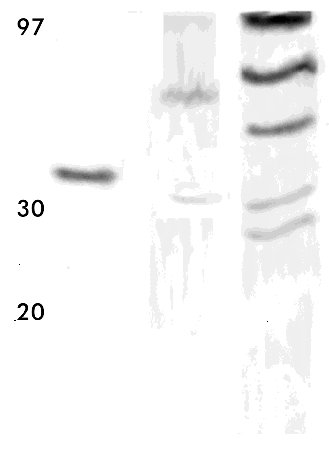
* Los tiempos de activación corresponden al tiempo de incubación a pH 2,5. Al comparar ambos sobrenadantes por electroforesis, también se observó que la pureza de la enzima secretada por K. lactis fue menor que la de la secretada por P. pastoris (Figura 1 carriles 3 y 4). Estos sobrenadantes de fermentación se concentraron y se prepararon crudos enzimáticos con una actividad coagulante similar. Figura 1. Electroforesis de los sobrenadantes de fermentación de las levaduras recombinantes P. pastoris y K. lactis que secretan la quimiosina bovina. Veinte microgramos de proteínas totales fueron aplicados en un gel de SDS-poliacrilamida 15%. Carrilera 1, patrón de peso molecular; carrilera 2, quimiosina natural; carrilera 3, sobrenadante de P. pastoris recombinante; carrilera 4, sobrenadante de K. lactis recombinante. En un ensayo Dot blot, se aplicaron volúmenes equivalentes de ambos crudos en cuanto a la actividad enzimática, y la cantidad de quimiosina se visualizó por revelado utilizando un conjugado anticuerpo policlonal-peroxidasa (Figura 2). Las diferencias en cuanto al reconocimiento de ambos crudos indican que la enzima secretada por P. pastoris es más activa. Al determinarse la actividad final de los crudos, se obtuvo que una unidad de actividad enzimática (UB) es equivalente a 133 m g de quimiosina para la proteína expresada por K. lactis y a 6,6 m g de quimiosina para P. pastoris. Esta última relación es similar a la que se obtuvo con la enzima natural utilizada como patrón en nuestro estudio. Esto representa para la enzima secretada por K. lactis una disminución de su actividad coagulante en 20,15 veces. Figura 2. Dot blot de los sobrenadantes de fermentación de las levaduras recombinantes P. pastoris y K. Lactis que secretan la quimosina bovina. Se midió la actividad de los sobrenadantes después de la activación de la enzima a pH 3,5 y se aplicaron sobre un filtro de nitrocelulosa cantidades que correspondían a la misma actividad enzimática en UB: 1a-9a, curva patrón de la quimiosina natural (20 ng, 50 ng, 100 ng, 200 ng, 500 ng, 1 m g, 2 m g, 5 m g, 10 m g); 1b y 2b, 0,075 UB y 0,15 UB, respectivamente, del sobrenandante de P. pastoris recombinante; 5b y 6b, 0,075 UB y 0,15 UB, respectivamente, del sobrenandante de K. lactis recombinante; 8b y 9b, 20 m g de proteínas del sobrenadante de fermentación de P. pastoris y K. lactis, respectivamente. Para comprobar las propiedades bioquímicas en cada caso, se determinó el efecto de diferentes condiciones de reacción (temperatura, concentración molar de iones Ca2+ y pH) sobre la actividad coagulante de los crudos, utilizando además, como control, la quimiosina bovina natural (Figura 3). En todos los casos se escogieron rangos de trabajo cercanos a los reales utilizados en alguna etapa de la elaboración de quesos a nivel industrial. Figura 3. Dependencia de la actividad coagulante de la
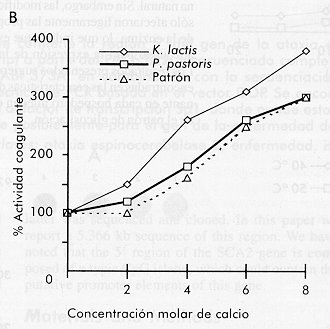
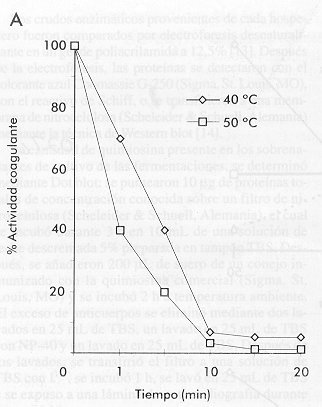
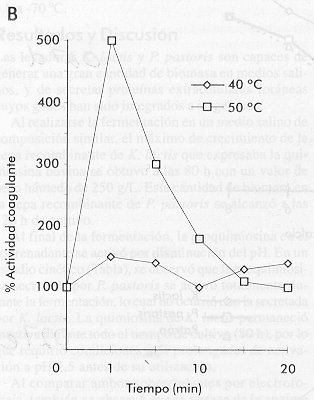
quimiosina bovina de: La Figura 3 muestra el resultado de estos experimentos. Es importante destacar que el comportamiento cinético de la quimiosina secretada por P. pastoris se acerca más al comportamiento de la enzima natural en todos los ensayos. En el estudio de la influencia de la temperatura de reacción sobre la actividad coagulante, la quimiosina secretada por K. lactis mostró una tendencia opuesta a la quimiosina patrón, aumentando su actividad con el aumento de la temperatura (Figura 3). Un estudio cinético de inactivación por calor del crudo enzimático a las temperaturas de escaldamiento o cocción utilizadas en la elaboración de quesos (40-50 ºC), mostró que la enzima secretada por K. lactis es mucho más resistente a la inactivación por calor que la secretada por P. pastoris (Figura 4). En este experimento se observó un incremento de la actividad coagulante por encima de 100% al incrementar la temperatura en el crudo proveniente de K. lactis. Este comportamiento puede deberse al hecho de trabajar con un crudo y no con la enzima purificada. Figura 4. Inactivación de la quimiosina a
diferentes temperaturas en función del tiempo, realizada a
las temperaturas de cocción utilizadas industrialmente en la
elaboración de quesos. En los quesos de maduración larga (> 30 días), una proteólisis excesiva provoca sabores amargos que afectan la calidad del queso. La relación entre la actividad proteolítica y la actividad coagulante, se estudió como criterio de actividad inespecífica en ambos crudos. Al comparar con la enzima natural (100%), la quimiosina proveniente de K. lactis mostró valores 2,5 veces mayor, mientras que la proveniente de P. pastoris mostró un incremento de 1,5 veces en la actividad proteolítica. Estos resultados sugieren que la enzima proveniente de K. lactis puede tener alguna modificación en su estructura que la diferencia de la enzima de P. pastoris. En cuanto a la disminución de la actividad coagulante y al incremento de la actividad proteolítica en el caso de la enzima secretada por K. lactis, en nuestro estudio se encontró un comportamiento similar al descrito por Aikawa y colaboradores [15] para la proteasa aspártica del hongo Mucor pusillus con actividad coagulante de la leche. Esta proteína es uno de los sustitutos comerciales de la quimiosina de ternero más generalizados. Estos autores encontraron que, al expresar esta proteasa en una cepa recombinante de la levadura S. cerevisiae, su actividad coagulante disminuyó a la mitad y su actividad proteolítica aumentó en la misma proporción. Cuando analizaron las modificaciones ocurridas, encontraron que el efecto se debía a la glicosilación de dos de los tres posibles sitios, los cuales no están modificados en la enzima natural. Para comprobar una posible modificación por glicosilación de la quimiosina proveniente de cada levadura objeto de estudio, se aplicaron 20 m g de los crudos enzimáticos sobre nitrocelulosa y se revelaron con un conjugado concanavalina A-peroxidasa. Este ensayo mostró una señal más intensa en la quimiosina proveniente de K. lactis que en la de P. pastoris (Figura 5A). Cantidades equivalentes a 20 m g de proteínas totales de los sobrenadantes de fermentación, se analizaron por electroforesis desnaturalizante estándar en geles laminares de poliacrilamida, seguido de tinción específica con base de Schiff (Figura 5B). Esta tinción mostró que ambos sobrenadantes contenían proteínas glicosiladas. Al analizar las mismas muestras mediante Western blot (Figura 5C), se observaron dos bandas para el sobrenadante de K. lactis, una que corresponde a la talla de la quimisina natural y otra de mayor talla. En el caso de P. pastoris, se observó un patrón similar al de la enzima natural utilizada como control. Figura 5. Análisis mediante Dot blot de los crudos
enzimáticos provenientes de los sobrenadantes de
fermentación de P. pastoris y K. lactis recombinantes,
electroforesis seguida de tinción con base de Schiff y
Western blot. La comparación del patrón obtenido por Western blot con el del gel teñido con el reactivo de Schiff, sugiere que la quimiosina es modificada por glicosilación en las dos levaduras. Sin embargo, en el caso de la enzima secretada por K. lactis se observa un cambio en la movilidad electroforética y en la talla de la proteína, posiblemente producto de la modificación. La banda superior de 46 kDa observada pudiera corresponder a la forma glicosilada de la enzima inactiva. Se puede concluir que las modificaciones efectuadas en la levadura afectaron más la proteína en el caso de la utilización de K. lactis como hospedero, por lo que las propiedades de esta enzima se alejan de las de la proteína natural. Sin embargo, las modificaciones en P. pastoris sólo afectaron ligeramente las propiedades bioquímicas de la enzima, lo que indica que este hospedero es más adecuado para la expresión de la quimiosina bovina. Los resultados presentados sugieren que las diferencias encontradas en las características de la enzima recombinante en cada hospedero pueden deberse a diferencias en el patrón de glicosilación. Referencias 1. Foltmann B. Prochymosin and chymosin (Prorennin and Rennin).
Meth Enzymol 1970;19:421-36. Copyright 1999 Elfos Scientiae The following images related to this document are available:Photo images[ba99027e.jpg] [ba99027d.jpg] [ba99027a.jpg] [ba99027h.jpg] [ba99027i.jpg] [ba99027j.jpg] [ba99027c.jpg] [ba99027f.jpg] [ba99027b.jpg] [ba99027g.jpg] | ||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}