 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnologia Aplicada 1999; Vol. 16 No. 3, pp. 169-172 Production of an Anti-carcinoembryonic Antigen Single-chain Fv Antibody Fragment as a Cytoplasmic Protein in Escherichia coli Javier E Vázquez,1 Freya Freyre,1 Marta
Ayala,1 Leonardo Canaán-Haden,2 1Division of Immunotechnology and Diagnostics, 2Division of
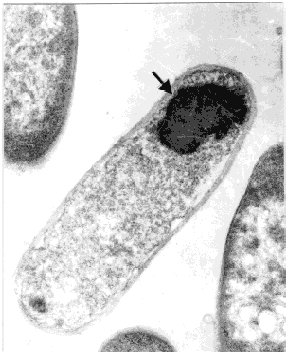
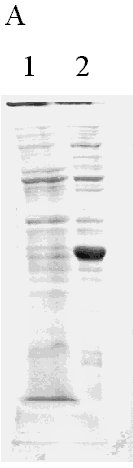
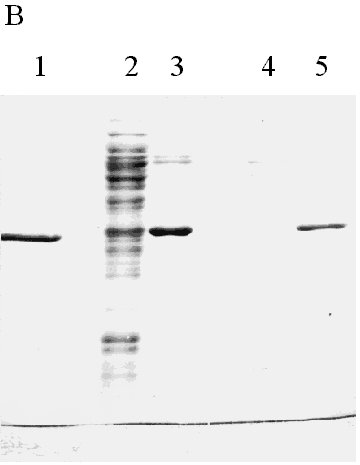
Physical Chemistry. Received in January,1999. Accepted for publication in March, 1999. Code Number: BA99029 ABSTRACT In this paper, a strategy to increase expression in Escherichia coli and simplify the purification of a single-chain Fv (scFv) antibody fragment specific for the carcinoembryonic antigen (CEA), is reported. The anti-CEA scFv fragment was expressed as an insoluble fusion protein in the cytoplasm of E. coli, with an N-terminal domain related in part with the first amino acids of human interleukin 2, and a C-terminal hexahistidine tag. The scFv fragment was extracted by solubilization, purified by immobilized metal ion affinity chromatography and refolded in a slightly alkaline buffer. With this system, the yield of active scFv increased four-fold when compared to a periplasmic variant of the same scFv, previously produced by the authors. Keywords: antibody fragments, carcinoembryonic antigen, CEA, E. coli, scFv RESUMEN En este trabajo se describe una estrategia para incrementar los niveles de expresión en Escherichia coli y simplificar el proceso de purificación, de un fragmento de simple cadena (scFv) de un anticuerpo específico contra el antígeno carcinoembrionario. El fragmento scFv expresado en el citoplasma de E. coli en forma de proteína fusionada e insoluble, posee en su extremo amino un dominio relacionado parcialmente con los primeros aminoácidos de la interleuquina 2 humana, mientras que en su extremo carboxilo incluye un dominio de seis histidinas consecutivas. El fragmento scFv fue extraído por solubilización, purificado mediante cromatografía de enlazamiento metálico y renaturalizado en una solución tampón de pH ligeramente alcalino. Con este sistema se incrementó en cuatro veces el rendimiento final de scFv activo, en comparación con una variante periplásmica previamente obtenida por los autores. Palabras claves: antígeno carcinoembrionario, CEA, E. coli, fragmentos de anticuerpos, scFv Introduction Ior-cea.1 is a mouse monoclonal antibody (MAb) specific for carcinoembryonic antigen (CEA) [1]. Technetium 99m-labelled Ior-cea.1 has successfully finished clinical trials, proving to be useful for in vivo diagnostic radioimaging of colorectal cancer and metastases [2, 3]. Single-chain Fv ( scFv) antibody fragments are small recombinant proteins in which the variable light (VL) and heavy (VH) chain domains of an antibody molecule are connected via an artificial linker to recreate the original binding site [4]. Advantages of scFv over whole antibodies are now explored for radioimmunodetection and in situ radiotherapy of cancer [4-7], due to potentially better tumor penetration and blood clearance, and reduced immunogenicity. Using reverse transcriptase polymerase chain reaction (RT-PCR), and starting with RNA from Ior-cea.1 hybridoma cells, we developed several scFv gene constructions that have been expressed as biologically active antibody fragments in the periplasm of Escherichia coli [8, 9]. A c-myc tagged anti-CEA scFv variant was expressed as a periplasmic protein in E. coli and purified using affinity chromatography with 9E10 MAbs [9], with a final yield of ca. 2 mg of >90% pure protein per liter of culture. With the aim of accumulating enough material for preclinical trials and further using the scFv in humans, this low yield and the need of MAb for purification impose important limitations. While advantageous with respect to the correct folding that can be achieved in this cellular compartment, the production of antibody fragments in the periplasm of E. coli is usually associated to low expression. Exception made of one report [10] where recombinant F(ab')2 fragments were expressed in the periplasm of E. coli at very high levels (1-2 g/L), the yield of recombinant Fab and scFv antibody fragments in this bacterium under conventional culture conditions is usually low, in the range of 1-10 mg/L [8, 11-15]. Another difficulty related to periplasmic expression is the frequent cell toxicity associated to protein accumulation in the periplasm, which affects scaling up the cultures to fermentors [9]. In this paper, the results in the development of an expression and purification strategy designed to overcome the aforementioned production limitations, are reported. The anti-CEA scFv antibody fragment has now been expressed in the cytoplasm of E. coli fused to the C-terminus of a fragment of 27 amino acids related in part to human interleukin-2 (hIL-2). The scFv fragment was also provided with a hexahistidine C-terminal tag for immobilized metal ion affinity chromatography purification (IMAC) [6]. Materials and Methods Cloning of the anti-CEA scFv gene in the pPACIB.9+ bacterial vector The pPACIB.9+ vector was designed for cytoplasmic expression of scFv antibody fragments in E. coli. This pBR322-based vector bears the tryptophan promoter, a histidine hexapeptide coding sequence at the 3'end of an EcoRI-EcoRV cloning site, and the T4 phage terminator sequence. The protein of interest is produced from pPACIB.9+ as an N-terminal fusion to a sequence of 27 residues related in part to the first amino acids of hIL-2 (M-A-P-T-S-S-S-T-A-Q-T-Q-L-E-L-E-H-L-L-L-D-L-G-I-F-L-E). The scFv gene insert to be cloned into pPACIB.9+ was obtained by PCR amplification [17] of the anti-CEA VH-linker-VL scFv coding sequence contained in the pF3 plasmid [9]. The following synthetic oligonucleotides were used, designed from the exact gene sequence: 5´-gggaattccaggtgaagctcctagagtcg-3´ (sense) The primers also provided the amplified DNA with EcoRI and EcoRV restriction sites, respectively. PCR was carried out using the PanoTaq thermostable polymerase (Panorama Research Inc., USA), according to the following temperature and time profiles: 94 °C for 3 min (1 cycle); 94 °C for 1 min, 50 °C for 1 min, 72 °C for 3 min (27 cycles); 72 °C for 8 min (1 cycle). The amplified scFv gene was gel-purified (Qiaquick Gel Extraction kit, Qiagen, Germany), EcoRI/EcoRV-digested (Promega, USA), and cloned into the pPACIB.9+ vector. XL-1 blue subcloning grade competent cells (Stratagene, USA) were transformed with the construction, grown in antibiotic medium (see below), and recombinant clones identified by restriction analysis of plasmid DNA from mini-preps ( Qiagen, Germany), followed by DNA sequencing [18]. The selected recombinant plasmid was denominated p1904His6. Bacterial culture media E. coli cells bearing the native and recombinant plasmids were grown in Luria Broth (LB) [18] containing ampicillin (100 µg/mL). Transformation of E. coli with the p1904His6 expression plasmid MM294 [F-, endA1, hsdR17, (rk-mk+), sup44, thi-1, relA1?, spoT1?] competent cells were transformed with the p1904His6 plasmid using standard procedures [18]. Initial selection of positive clones was performed by PCR using the same primers described above. Selected bacterial clones were used for expression studies. Expression analysis Recombinant MM294 cells grown for 12 h at 37 °C in LB medium with ampicillin. Dilutions (1:100) of overnight cultures in fresh medium were taken as starting material for the scFv fragment expression experiments. Cells were grown for 2 h at 37 °C and induced for 6 h at 37 °C with 30 µg/mL of 3-beta-indoleacrylic acid in ethanol. Cells were then washed, lysed by sonication, and centrifuged. The soluble and insoluble cellular fractions were analyzed by 15% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) under reducing conditions [19], and by Western blot. Protein blotting to nitrocellulose was performed using a Trans-Blot SD apparatus (Bio-Rad, USA). The nitrocellulose membrane was incubated with specific rabbit anti-scFv IgG, generated with purified periplasmic anti-CEA scFv preparations obtained using the pF3 plasmid [9], followed by incubation with horseradish peroxidase (HRPO)-conjugated goat anti-rabbit antibodies (Sigma, USA). Recombinant bacteria were also fixed with 3.2% glutaraldehyde, dehydrated in increasing concentrations of ethanol, and included in Spurr [20]. Ultrafine sections were obtained with an LKB ultramicrotome, deposited in copper grids, and observed in a JEOL-JEM 2000 EX transmission electron microscope. Production of the scFv fragment in a laboratory fermentor Recombinant MM294 cells carrying the p1904His6 plasmid were cultured in a 1.5-L fermentor (B.E. Marubishi, Japan) using LB medium. The inoculum (100 mL of an overnight culture) was added at time 0, temperature was adjusted to 37 °C, air pressure to 1 vvm, and agitation to 400 rpm. The inductor (3-beta indoleacrylic acid at 25 µg/mL in absolute ethanol) was added 2 h later. Expression was maintained for 8 h, and E. coli cells were finally collected by a 20-min centrifugation at 4 °C and 2000 xg. Extraction, refolding and purification of the scFv fragment The bacterial cell pellet after induction was suspended at 10 mg/mL in PBS containing 4 M urea and sonicated for 4 cycles (130-150 W, 1 min each with 2-min intervals) in a bench-top sonicator (B. Braum Biotech, Germany). After 30 min on ice, samples were centrifuged for 5 min at 14 000 xg, 4 °C. The resulting pellet was incubated for 2 h at 4 °C in PBS supplemented with 8 M urea, pH 9.0. After centrifugation, the soluble fraction (with the scFv fragment) was collected and extensively dialyzed at 4 °C in a renaturation buffer (10 mM NaHCO3, 150 mM NaCl, pH 8.0). After dialysis, the final sample was clarified by centrifugation, and loaded onto a Sepharose-IDA-Ni2+ column (Pharmacia, Sweden), equilibrated with 50 mM phosphate buffer, pH 8.0. The column was washed with the same buffer at pH 6.3, 5.5 and 5.0, and the scFv eluted at pH 4.0. The fractions were neutralized and analyzed by SDS-PAGE and Western blot. Protein concentration estimates Protein concentration was determined as suggested by Bradford [21], using bovine serum albumin as standard. Anti-CEA ELISA The biological activity of scFv from bacteria was studied with a specific direct ELISA. Briefly, 96-well microtiter plates (Costar) were coated overnight at 4 °C with CEA (Instituto Nacional de Oncología y Radiobiología, Havana, Cuba) to 0.1 µg per well, using carbonate-bicarbonate buffer pH 9.6. After washing with PBS, the samples to be studied were applied at 10 µg/mL and incubated for 1 h at 37 °C. Anti-scFv specific rabbit IgG, followed by HRPO-conjugated goat anti-rabbit antibodies, were used to develop the reactions with o-phenylenediamine (OPD) in citrate buffer containing 0.005% hydrogen peroxide. The periplasmic anti-CEA scFv fragment [9] was used as positive control. An irrelevant anti-HBsAg scFv fragment was used as negative control [11]. Results and Discussion Increased expression of heterologous proteins in bacteria via genetic fusion with well-characterized sequences, due to the formation of insoluble inclusion bodies, or augmented stability of messenger RNA, has been well documented [22, 23]. In this sense, it has been shown that high cytoplasmic expression of viral and bacterial antigens can be achieved with a specific fusion strategy in which the genes are cloned into vectors under the control of the tryptophan promoter, downstream from the first 60 amino acids of hIL-2 [24-26]. To apply the same strategy to antibody fragment production, the pPACIB9+ vector was designed, in which the hIL-2 related domain was reduced to 27 amino acids with several amino acidic changes, and the cloning sites have been made compatible with antibody variable regions. Since antibody fragments produced in the bacterial cytoplasm are usually biologically inactive, several in vitro refolding procedures to restore antigen recognition were developed [8, 27-32]. The anti-CEA scFv gene was modified by PCR to allow cloning into pPACIB.9+, and several bacterial strains were transformed with the resulting recombinant plasmid p1904His6. The best expression results were reported in MM294, where approximately 20% of the total bacterial protein content corresponded to a new recombinant protein. TEM indicated that it was mainly produced as insoluble cytoplasmic inclusion bodies (Figure 1), while SDS-PAGE shows an expected band of approximately 30-32 kDa (Figure 2A), and Western blot analysis corroborated that the protein was the scFv fragment. Figure 1. Transmission electron micrograph of recombinant MM294 cells, transformed with the p1904His6 plasmid. A large cytoplasmic inclusion body is marked with an arrow. MM294 cells transformed with the p1904His6 plasmid were then cultured and induced in a 1.5-L fermentor. In comparison with data reported for an anti-CEA periplasmic scFv counterpart [9], it was found that expression and optical density increased three-fold (15% vs. 5%), and four-fold (17 vs. 4 OD at 620 nm), respectively, with close to 10 times more final scFv per fermentation for the cytoplasmic variant. In the extraction process of the insoluble cytoplasmic scFv 4 M urea was first used, which washed most of the contaminant E. coli proteins, and then 8 M urea, which completely solubilized the antibody fragment (Figure 2B). The scFv fragment was then refolded in an alkaline buffer and loaded onto a Sepharose-IDA-Ni2+ matrix. Elution of the scFv fragment was performed at pH 4.0, with a final purity of the antibody fragment of approximately 95%, as estimated by SDS-PAGE and densitometry (Figure 2B). The final yield of scFv after refolding and purification was of approximately 17 mg per liter of culture. Figure 2. Expression and purification analysis of the
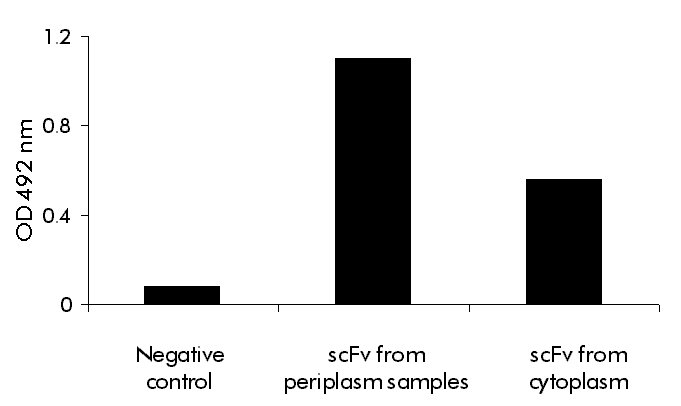
bacterial anti-CEA scFv fragment. While this value represents an 8.5-fold increase in production yield with respect to the periplasmic version of the scFv fragment [9], it has to be considered that refolding efficiency is not 100%. Then, the 9E10 affinity-purified anti-CEA scFv fragment and the refolded cytoplasmic scFv fragment, at similar concentrations, were compared by ELISA. The periplasmic CEA was taken as a control of the "maximum" amount of correctly folded molecules, per mass of protein. As expected, the refolded scFv fragment has approximately 50% of the specific activity of the periplasmic scFv fragment, if optical densities are compared (Figure 3). Still, even adjusting for this parameter, the production yield of active molecules for the refolded scFv fragment is over four times that of the periplasmic antibody fragment. Figure 3. ELISA to detect biological activity after refolding the fused scFv fragment. Polystyrene plates were coated with natural CEA, and cytoplasmic refolded and purified scFv, or 9E10 affinity-purified scFv from periplasm were added at 10 µg/mL and incubated for 1 h at 37 °C. An anti-HBsAg scFv at the same concentration was used as negative control. The specific binding was detected by adding rabbit IgG raised against either the anti-CEA periplasm scFv or the anti-HBsAg scFv. The reaction was developed with an anti-rabbit IgG HRPO-conjugate, followed by substrate solution. Optical densitiy (OD) was determined at 492 nm. In conclusion, an expression strategy that increases scFv yield and substitutes MAbs-affinity chromatography by IMAC with similar end purity, has been designed. Another potential advantage of expression to insoluble form was derived from the possibility of developing culture procedures for high level production [25]. As aforementioned, periplasmic expression usually brings about severe cell toxicity, and using a similar expression system for another scFv it has been seen that the optical density of the cultures can rise approximately four times, when expression is switched from periplasm to cytoplasm [32]. References 1. Tormo B, Gavilondo JV, Domínguez C, Freyre FM,
Rodríguez T, Biberfeld P. CEA in colonic adenocarcinomas and
precancerous lesions. An immunohistochemical study with a novel
monoclonal antibody. APMIS 1989;97:1073-80. Copyright 1999 Elfos Scientiae The following images related to this document are available:Photo images[ba99029a.jpg] [ba99029d.jpg] [ba99029b.jpg] [ba99029c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}