 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnologia Aplicada 1999; Vol. 16 No. 3, pp 177-193 REPORTS Code Number: BA99031 La oncoproteína E7 del virus del papiloma humano tipo 16 (VPH 16) es capaz de inducir resistencia al efecto citostático del IFN-a 2b Desarrollo de una nueva metodología para la caracterización estructural de la N-glicosilación de proteínas naturales o recombinantes Identificación y caracterización molecular de geminivirus bipartitos en Cuba Aportes quimiotaxonómicos al estudio del Mycobacterium habana, especie cubana candidato vacunal Factores tóxico-nutricionales y neuropatía epidémica. Programa de investigaciones "SECUBA" Obtención de plantas transgénicas de caña de azúcar (Saccharum officinarum L) mediada por Agrobacterium tumefaciens: una nueva metodología para la transformación genética de esta gramínea Desarrollo de las aplicaciones de los marcadores moleculares en el mejoramiento de la caña de azúcar en Cuba Desarrollo de la crioconservación de germoplasma vegetal en Cuba Humanización de epitopos T en las regiones variables murinas de un anticuerpo que reconoce al receptor del factor de crecimiento epidérmico: obtención de inmunoglobulinas de menor inmunogenicidad Biotecnologia Aplicada 1999; Vol. 16 No. 3, pp 177-178 La oncoproteína E7 del virus del papiloma humano tipo 16 (VPH 16) es capaz de inducir resistencia al efecto citostático del IFN- a 2bAlejandro Moro,1 Andrea Calixto,1 Eduardo Suárez,2 Manuel de J Araña,1 Silvio E Perea1 1Centro de Ingeniería Genética y
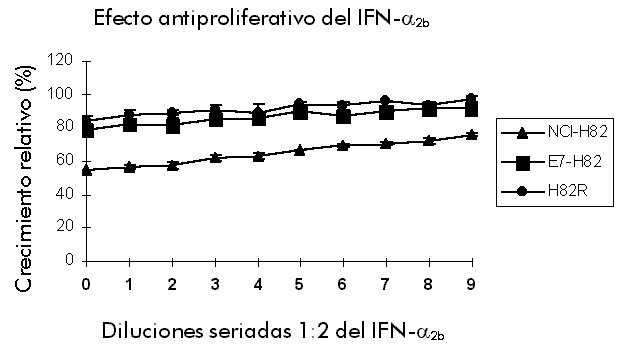
Biotecnología. AP 6162, La Habana 10600, Cuba. Introducción Los interferones (IFN) son importantes citoquinas con efectos antiproliferativo, antiviral y de aumento de la respuesta inmune. Debido a estas propiedades, los IFN son utilizados en la terapia de enfermedades virales y no virales. Sin embargo, algunos estudios clínicos demuestran que puede haber recurrencia o ausencia de respuesta con frecuencia. Los mecanismos de esta resistencia no se conocen aún y se especula que la presencia del virus del papiloma humano (VPH) y su interacción con moléculas de señalización inducidas por los IFN, pudieran formar parte de estos mecanismos de resistencia. Aunque se conoce bien la vía de señalización desde los receptores de los IFN hasta el núcleo, aún queda por estudiar la relación entre los factores de transcripción activados, los genes reguladores del ciclo celular y las oncoproteínas. En este estudio [1] se comparan los efectos del IFN-a 2b sobre los niveles de ARN mensajero (ARNm) de los genes de la p27Kip1 y la 2'-5' Oligoadenilato-sintetasa (2'-5' OAS) en una línea celular sensible (NCI-H82) y otra resistente (H82R). Además, se introdujo la oncoproteína E7 en la línea NCI-H82 y se evaluó la respuesta al IFN-a 2b y la inducibilidad de los genes de p27Kip1 y 2'-5' OAS. Se demostró por primera vez mediante un ensayo antiproliferativo in vitro que la oncoproteína E7 es capaz de inducir resistencia al efecto citostático del IFN-a 2b. Además, se obtuvo la expresión diferencial del gen regulador del ciclo celular p27Kip1, cuya expresión fue correlacionada con el fenotipo de respuesta al IFN. Resultados y Discusión Efecto del IFN-a 2b sobre p27Kip1 y 2'-5' OAS en las líneas celulares NCI-H82 y H82R Dos líneas celulares con diferente respuesta al IFN-a 2b fueron comparadas para determinar si esta citoquina era capaz de alterar la abundancia del ARNm del gen p27Kip1. Se llevó a cabo un experimento cinético con muestras tomadas a las 0, 8, 24, 48 y 72 horas posterior a la adición del IFN-a 2b. Se aisló el ARN total y se desarrolló una reacción de transcripción reversa, seguido de una reacción en cadena de la polimerasa (RT-PCR). Los productos de la RT-PCR fueron separados por electroforesis en gel de agarosa y se obtuvo una imagen digital del patrón de separación. En la línea celular sensible (NCI-H82) se obtuvo que el IFN-a 2b induce un aumento en los niveles de ARNm de p27Kip1 a partir de 24 h, con un máximo a las 48 h aproximadamente tres veces superior a los niveles del transcrito en ausencia de IFN-a 2b. En ausencia de IFN-a 2b no hubo variación en los niveles de p27Kip1. Este hecho descarta la posibilidad de que la inducción observada se deba a algún factor inespecífico presente en el suero o en el medio de cultivo. La inducción del gen de la 2'-5' OAS se tomó como control de la unión del IFN-a 2b a su receptor y del desencadenamiento de la vía de señalización. En el caso de la línea NCI-H82, se observó un máximo de inducción 8 h después de la adición del IFN-a 2b, con disminución progresiva, lo que se corresponde con la cinética de inducción temprana reportada para este gen en la literatura. En ausencia de IFN-a 2b (control negativo) no se expresó el gen de la 2'-5' OAS. Resulta interesante que en las células de la línea H82R, no se expresó el gen de la 2'-5' OAS con la misma cinética que en las células sensibles, mientras que los niveles de p27Kip1 no variaron en ningún momento después de la adición del IFN-a 2b. La inducción del gen de la 2'-5' OAS en la línea sensible, descarta la posibilidad de afectación del receptor del IFN-a 2b en estas células. Este constituye el primer reporte en el mundo sobre la expresión diferencial del gen p27Kip1 en células sensibles y en células resistentes al efecto antiproliferativo del IFN-a 2b. Los resultados reflejados aquí tienen gran importancia debido a que el IFN-a 2b ha sido aprobado en Cuba y en el mundo para la terapia de diversas enfermedades por sus efectos antivirales, antiproliferativos y de aumento de la respuesta inmune. Cierto porcentaje de las personas tratadas con IFN-a 2b no responden o muestran recurrencia después de un período de tratamiento determinado. En este trabajo se muestran los resultados de un estudio comparativo realizado en células sensibles y en células resistentes, acerca del efecto del IFN-a 2b sobre un gen regulador del ciclo celular p27Kip1, que ya se ha demostrado que está relacionado con la detención del ciclo celular. El gen p27Kip1 puede que se exprese como parte de la vía de señalización de esta citoquina. Los resultados de este trabajo sugieren que el gen p27Kip1 pudiera ser un mediador clave en el efecto antiproliferativo del IFN-a 2b, y que si no se expresa pueden aparecer células resistentes. Este resultado ayuda en el establecimiento de las bases para una terapia racional, ya que es probable que no respondan aquellos pacientes que tengan el gen p27Kip1 mutado o afectada su inducibilidad por IFN-a 2b. Sería posible entonces identificar, previo al tratamiento, los mejores candidatos para una terapia efectiva. Efecto de la oncoproteína E7 del VPH 16 sobre la respuesta de las células NCI-H82 frente al IFN-a 2b La expresión del oncogén E7 del VPH 16 induce la entrada en la fase S del ciclo celular en presencia de señales antiproliferativas. En este estudio se investigaron los niveles relativos del gen p27Kip1 mediante RT-PCR en células sensibles al IFN-a 2b transfectadas con el gen E7. Los niveles de p27Kip1 no variaron después de la adición del IFN-a 2b. Esto coincide con lo observado para los niveles de ARNm de p27Kip1 en las células resistentes. La respuesta de las células transfectadas al IFN-a 2b fue evaluada mediante un ensayo anti-proliferativo, con la utilización de las líneas NCI-H82 y H82R como controles (Figura). Se obtuvo que las células NCI-H82 son muy sensibles: concentraciones tan bajas como 7,5 UI/mL de IFN-a 2b producen 30% de inhibición. Sin embargo, en el caso de las células transfectadas y H82R hubo muy poca variación, con niveles de crecimiento relativo entre 77 y 97%. El análisis estadístico de estos resultados mostró diferencias significativas entre la línea NCI-H82 y las células H82R y las transfectadas con el gen E7. En teoría se pudiera explicar la ausencia de respuesta de las células transfectadas, si se toma como base un reporte previo [2] que indica que la capacidad de E7 de superar ciertas formas de detención del ciclo celular en G0/G1, se debe en parte a su unión al inhibidor p27Kip1, lo que trae como consecuencia que p27Kip1 no pueda inhibir la actividad quinasa asociada a la ciclina E/Cdk2. Los VPH han sido identificados como agentes causales de diversas lesiones epiteliales proliferativas como el condiloma acuminado y las verrugas comunes. Estos virus han emergido rápidamente como la enfermedad viral de transmisión sexual de mayor prevalencia en la actualidad. Según estimados, uno de cada tres individuos sexualmente activos está infectado con algún tipo de VPH. Aproximadamente un millón de casos nuevos son diagnosticados cada año. También se ha señalado, como factor etiológico del cáncer de cérvix, que aproximadamente 90-95% de los pacientes con cánceres cervicales invasivos contienen ADN de VPH. Por otra parte, los IFN han sido aprobados para la terapia de diversas enfermedades. Sin embargo, estudios clínicos demostraron que con frecuencia hay recurrencia o ausencia de respuesta. Los mecanismos de resistencia no se conocen, aunque la presencia de las proteínas de los VPH y sus interacciones con las moléculas de señalización celular pudieran estar involucrados en esta resistencia. Figura. La transfección del gen E7 del VPH 16 en la línea celular NCI-H82 induce resistencia al efecto antiproliferativo del IFN-a 2b. Se utilizó un rango de concentraciones de IFN-a 2b (Heber Biotec, Cuba) de 7,5 UI/mL a 4 000 UI/mL. Se utilizaron diluciones seriadas 1:2 y se determinó la proliferación celular mediante un ensayo colorimétrico. Los datos son presentados como por cientos de células no tratadas. Los puntos representan la media de las mediciones de tres ensayos diferentes y las barras representan la desviación estándar. Los asteriscos indican diferencias significativas (p < 0,01) con respecto a las células NCI-H82. En este trabajo se establecen, por primera vez, las bases moleculares de la resistencia de tipos oncogénicos de los VPH al tratamiento con IFN-a 2b. La posible vinculación de la presencia de oncoproteínas con la resistencia al tratamiento, permitiría establecer una racionalidad en la terapia con el IFN-a 2b. El clínico sabría antes del comienzo del tratamiento y teniendo en cuenta los resultados de las pruebas de detección de VPH, si el paciente responderá o no al mismo. Esto mejoría la calidad de vida del paciente, pues le evitaría someterse a protocolos con poca probabilidad de éxito y obligaría a diseñar nuevos protocolos que aumenten la eficacia del tratamiento, mediante la inclusión de nuevos fármacos cuya actividad combinada resulte más efectiva que el IFN-a 2b solo. De este modo, aumentaría el número de pacientes curados y, por lo tanto, la eficiencia del sistema nacional de salud. Copyright 1999 Elfos Scientiae
Biotecnologia Aplicada 1999; Vol. 16 No. 3, pp 178-180 Desarrollo de una nueva metodología para la caracterización estructural de la N-glicosilación de proteínas naturales o recombinantes José A Cremata, Omar Quintero, Raquel
Montesino, Rossana García Centro de Ingeniería Genética y
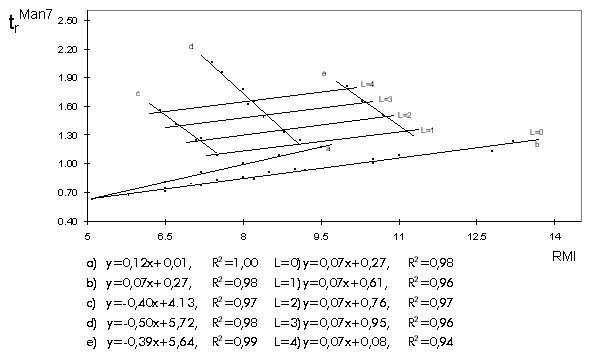
Biotecnología, AP 6162, Ciudad de La Habana, Cuba. Introducción Se describe una metodología nueva, sencilla y sensible para el análisis y caracterización estructural de oligosacáridos N-enlazados de glicoproteínas naturales o recombinantes. Esta metodología está incluida en las llamadas técnicas de perfil de mapeo bidimensional de oligosacáridos neutros y sialilados, mediante el empleo del ácido 8-amino-1,3,6-naftalen trisulfónico (ANTS) como agente derivatizante fluorescente. Como principales virtudes de esta metodología se destacan: 1. Se requieren sólo picomoles de oligosacárido para su caracterización, lo que representa microgramos de la glicoproteína de interés a diferencia de los métodos tradicionales (RMN), que utilizan cantidades superiores en dos ordenes de magnitud. 2. Permite prescindir, en muchos casos, del uso de métodos físico-químicos de análisis muy especializados y caros como la espectrometría de masas y la resonancia magnética nuclear protónica de alta frecuencia. 3. Se pudiera incluir como método de análisis de la glicosilación lote a lote en procesos productivos de glicoproteínas recombinantes. Descripción del resultado e importancia La metodología propuesta incluye los siguientes pasos: 1. Reacción de aminación reductiva con ANTS de los oligosacáridos liberados por hidrólisis enzimática con la endoglicosidasa PNGasa F. 2. Separación simultánea o consecutiva en columna de aminoadsorción (NH2-HPLC) y/o mediante electroforesis en geles de poliacrilamida (FACE). 3. Representación bidimensional de los valores de tiempo de retención relativo con respecto a una referencia interna, Man7GlcNAc2, (HPLC) e índice de migración relativo con respecto a una serie de maltooligosacáridos (FACE). 4. A partir del procedimiento anterior, se construye un mapa bidimensional donde se incluyen compuestos de referencia previamente reportados en la literatura o caracterizados por métodos adecuados, y las muestras problema son caracterizadas por su localización en este mapa (Figura y Tabla). Figura. Representación gráfica de los valores de tiempo de retención relativo (trMan7) e indice de migración relativo (RMI) de los compuestos de referencia. Las estructuras incluidas en este gráfico pertenecen a las familias siguientes: a) oligomanósidos desde Man5GlcNAc2 hasta Man9GlcNAc2; b) complejos asialilados del tipo diantenarios, triantenarios y tetraantenarios; c) diantenas sialiladas; d) triantenas sialiladas y e) tetraantenas sialiladas. Las líneas L representan familias de compuestos de diferente número de antenas pero con similar carga total efectiva dada por la contribución de ácidos siálicos con enlaces a 2,3 o a 2,6. Debajo del gráfico aparecen las ecuaciones que representan a cada recta con sus valores correspondientes de R2. Los valores de coeficientes de correlación son mayores o iguales a 0,97 para las ecuaciones a-e y mayores o iguales a 0,94 para las L=0-4. Las pendientes de las ecuaciones a y b resultaron positivas mientras que las de las ecuaciones c-e fueron negativas debido a la contribución negativa de los ácidos siálicos enlazados a los terminales no-reductores de los oligosacáridos. En caso de análisis de muestras problemas se introducen los datos de trMan7 y RMI, y según su ubicación en el mapa así será la estructura propuesta. Cuando la coincidencia no es total entonces para la proposición de estructura se tiene en cuenta la contribución de cada monosacárido a los valores de tiempo de retención en HPLC y migración electroforética (Tabla). Tabla. Contribución a los valores de RMI y trMan7de los monosacáridos más abundantes de los oligosacáridos N-enlazados a glicoproteínas.
(a) El error experimental en el valor de RMI es menor de 2%.(b)El error experimental en el valor de trMan7 es menor de 2%. (c)r - Calculado como el cociente de los valores de tiempo de retención relativo e índice de migración relativo. Este parámetro refleja el comportamiento de los diferentes tipos de monosacáridos dependiendo de su naturaleza diferenciandose claramenete entre oligosacáridos neutros y sialilados. Observe que entre los monosacáridos neutros la manosa se diferencia ligeramente del resto. Con respecto a la contribución de los ácidos siálicos, a pesar de la contribución diferenciada en cuanto a la configuración del enlace (a 2,3 o a 2,6) en términos de D RMI y D trMan7, el cociente "r" para ambos es prácticamente el mismo. 5. En el caso necesario, se diseñarían experimentos adicionales de digestión con exoglicosidasas específicas o de interacción lectina-oligosacárido para obtener información complementaria sobre la estructura, lo que eliminaría posibles ambigüedades en la asignación inicial. El resultado que se describe incluye: • Caracterización de la glicosilación de proteínas modelo reportadas en la literatura (a 1-antitripsina, IgG2a y orosomucoid o glicoproteína acida a 1). • Caracterización de la glicosilación de seis proteínas recombinantes expresadas en la levadura Pichia pastoris (dextranasa, a -amilasa, invertasa, Mucor rennina, antígeno de la garrapata Bm86 y subunidad catalítica de la enteroquinasa bovina). • Caracterización de la glicosilación de anticuerpos monoclonales de diferentes orígenes (IOR R3 de ascitis de ratón y obtenido en biorreactores). • Caracterización de la glicosilación de un panel europeo de sueros estándares de pacientes con artritis reumatoide y muestras de pacientes cubanos. • Caracterización de la glicosilación de la eritropoyetina humana recombinante expresada en células CHO. Importancia del resultado Esta metodología permite, de forma sencilla y económica y al alcance de cualquier laboratorio de bioquímica o biología molecular, la caracterización de la N-glicosilación de glicoproteínas naturales o recombinantes. Esta caracterización se torna más imprescindible cada día, debido a que se está introduciendo como medida regulativa para glicoproteínas de interés médico-farmacéutico, a partir del reconocimiento científico del papel decisivo que pueden jugar los residuos de oligosacáridos covalentemente enlazados a las proteínas, tanto en sus características físico-químicas como en su actividad biológica. Un ejemplo ilustrativo de esto lo constituye la hormona eritropoyetina, que cuando es glicosilada en forma incorrecta es perfectamente reconocida in vitro por los receptores celulares, sin embargo, su actividad in vivo es prácticamente despreciable por presentar un tiempo de vida medio de circulación muy pequeño. La caracterización de la glicosilación por métodos físico-químicos, además de requerir un equipamiento muy costoso como el de la resonancia magnética nuclear de alta frecuencia, necesita cantidades de muestra que pueden alcanzar las decenas o centenas de miligramos de glicoproteína. Este último aspecto se convierte en una limitación importante por el alto valor de estos productos. El desarrollo y la introducción de los métodos de perfil para la caracterización de la glicosilación de proteínas naturales o recombinantes resuelve este problema y permite, a su vez, una mejor caracterización del producto, incluso por lote de producción, en aquellos casos que lo requieran. Desde el punto de vista del aporte científico, esta metodología tiene ventajas sobre otras descritas, entre las que se encuentran: 1. Permite analizar, en un único sistema, tanto oligosacáridos neutros como cargados (sialilados, fosfatados, sulfatados, etc.), lo cual representa una limitación para otros métodos. 2. Presenta la mayor resolución reportada hasta el momento para los oligosacáridos cargados. 3. Constituye el primer reporte de agrupación de las estructuras de oligosacáridos de N-glicosilación en familias de homología, en dependencia de la estructura y grado de ramificación (series homólogas). 4. El método tiene carácter predictivo (dado por lo descrito en el punto anterior y por la contribución de cada monosacárido a los valores de tiempo de retención relativo e índice de migración relativo), lo que facilita el diseño de experimentos con exoglicosidasas específicas en los casos de estructuras no reportadas. 5. El tipo de derivado fluorescente que se utiliza también puede ser analizado por espectrometría de masas con ionización por electronebulización. Esta propiedad permite realizar análisis con esta técnica para corroborar los resultados en casos de ambigüedad. Agradecimientos Los autores agradecen la valiosa colaboración del Lic. Lorenzo Vega, del Instituto de Investigaciones del Transporte, y de la técnica Marlén Mata, del CIGB. Referencias 1. Quintero O, Montesino R, Cremata JA. Two-dimensional
mapping of 8-amine-1,3,6-naphthalene trisulfonic acid derivatives
of N-linked neutral and sialyloligosaccharides. Anal Biochem
1998;256: 23-32. Copyright 1999 Elfos Scientiae
Biotecnologia Aplicada 1999; Vol. 16 No. 3, pp 180-181 Identificación y caracterización molecular de geminivirus bipartitos en Cuba Yamila Martínez Zubiaur,1 Pedro L Ramos
González,2 Rudy Peral,2 1Centro Nacional de Sanidad Agropecuaria. Introducción Los geminivirus, en particular los miembros del género Begomovirus, son considerados el problema fitosanitario principal del cultivo del tomate en las regiones tropicales y subtropicales, y ocasionan pérdidas cuantiosas. Cuba no escapa a esta situación emergente y se han descrito porcentajes elevados de infección viral en las áreas comerciales del país desde finales de 1989. Los objetivos de este trabajo han estado encaminados a realizar la identificación molecular de los geminivirus presentes en las plantaciones cubanas de tomate, y desarrollar los métodos moleculares para el diagnóstico rápido y preciso de geminivirus en las áreas de producción. Se identificaron por primera vez en Cuba dos geminivirus bipartitos nuevos, los cuales fueron denominados virus "Habana" del tomate (HTV) y virus "Taíno" del moteado del tomate (TTMoV). Ambos virus poseen dos componentes genómicos (ADN-A y ADN-B) con secuencias diferentes, a excepción de la región común, en la cual se encuentran sitios y estructuras moleculares involucradas en la replicación y transcripción virales. El análisis de los marcos abiertos de lectura demostró que los virus caracterizados están filogenéticamente relacionados con los del género Begomovirus del nuevo mundo. La caracterización biológica y molecular de estos virus ha derivado en la obtención de diagnosticadores empleados en la actualidad para la determinación de los agentes etiológicos causantes de epifítias en cultivos de interés económico para el país. Resultados y Discusión El presente trabajo recoge los resultados de identificación molecular de dos geminivirus bipartitos nuevos presentes en las plantaciones cubanas de tomate. Se describen dos procedimientos para el aislamiento y la clonación de estos patógenos: 1) la clonación de genomas totales a partir de fragmentos amplificados por la reacción en cadena de la polimerasa (RCP), y 2) la clonación a partir del intermediario replicativo geminiviral. A través de la secuenciación de los genomas clonados, se identificaron dos geminivirus bipartitos (HTV y TTMoV) diferentes a los descritos previamente en la literatura. La infectividad de ambos virus fue comprobada mediante la transferencia de sus genomas a plantas hospederas por medio de la agroinoculación y la biobalística. Las comparaciones de las secuencias de los diferentes polipéptidos codificados por cada componente viral con las de otros geminivirus, demostraron que tanto TTMoV como HTV están relacionados filogenéticamente con otros virus aislados de la cuenca del Caribe, América del Sur y la Florida. La caracterización molecular y biológica de ambos virus permite profundizar en el conocimiento de la etiología de la principal epifítia que afecta los rendimientos de las plantaciones cubanas de tomate. Además, constituye un paso fundamental en el establecimiento de un diagnóstico certero y de una vía de seguimiento de la propagación de la enfermedad hacia otros cultivos con potencialidad como hospedero. El genoma viral, una vez caracterizado, constituye una fuente de genes para las numerosas estrategias de obtención de plantas transgénicas más tolerantes a las infecciones virales mediante la ingeniería genética. Los estudios de caracterización desarrollados en el presente trabajo han contribuido al conocimiento de la diversidad de geminivirus presentes en la región de Centroamérica y el Caribe. Principales aportes científicos • Identificación y aislamiento de dos geminivirus
nuevos en las plantaciones cubanas de tomate (TTMoV y HTV). No
existen antecedentes de estos virus en la literatura. Bibliografía Ramos PL, Guerra O, Doreste V, Ramírez N,
Rivera-Bustamante R, Oramas P. Detection of TYLCV in Cuba. Plant
Disease 1996;80:1208. Copyright 1999 Elfos Scientiae
Biotecnologia Aplicada 1999; Vol. 16 No. 3, pp 181-182 Aportes quimiotaxonómicos al estudio del Mycobacterium habana, especie cubana candidato vacunal Lilian M Mederos Cuervo,1 Pedro L Valero-Guillén,2 José A Valdivia Álvarez1 1Instituto de Medicina Tropical "Pedro Kourí". AP 601,
Marianao 13, Ciudad de La Habana, Cuba. Introducción A escala internacional, la tuberculosis (TB) ha mantenido una evolución desfavorable a partir de la década de 1980. En la actualidad, el resurgir de la enfermedad no afecta solamente a los países industrializados. La Organización Mundial de la Salud (OMS) ha manifestado que este fenómeno se debe a la crisis económica, a la pérdida de prioridad de los Programas Nacionales de Control de esta enfermedad, al incremento acelerado del Síndrome de Inmunodeficiencia Adquirida (SIDA) y a la aparición de cepas resistentes. Se ha estimado que si la situación actual prevalece, se producirán aproximadamente 90 millones de casos nuevos de tuberculosis en el mundo durante la década 1990-1999, y unos 30 millones de defunciones deben ocurrir hasta el año 2000 a causa de esta enfermedad. Por otra parte, la pandemia del SIDA ha tenido una influencia notable en la epidemiología de la TB, tanto en países en desarrollo como en los desarrollados. En Cuba, se reportó el primer caso de TB en un seropositivo al virus de inmunodeficiencia humana (VIH) en enero de 1993. Junto al incremento de Mycobacterium tuberculosis se aíslan otras micobacterias de interés clínico denominadas micobacterias no tuberculosas (MNT). Dentro del grupo de especies denominadas MNT, se debe resaltar el reporte de dos especies micobacterianas que han sido tema de investigación de diferentes estudios. Estas especies son M. simiae (1965) y la especie cubana denominada M. habana (1971), las cuales presentan semejanzas en algunas pruebas diagnósticas tanto convencionales como alternativas y han sido tema de discusión. Sin embargo, en la década de 1990, M. habana ha sido objeto de importantes estudios. Por ejemplo, en esta especie se reporta por primera vez la presencia de la fracción antigénica de 18 kDa de M. leprae (1995), lo que constituyó un hallazgo muy importante que hizo posible que esta especie (M. habana) fuera propuesta como cepa candidata para el desarrollo de una vacuna contra la lepra, hecho que no se había podido consolidar hasta entonces con ninguna otra especie micobacteriana. Al mismo tiempo, otro colectivo de investigadores demostró que esta especie cubana también protege contra la tuberculosis. En la actualidad, M. habana es propuesta como posible candidato vacunal contra la lepra, la tuberculosis y la úlcera de Burulí, puesto que se tienen evidencias de que protege contra estas enfermedades. Más tarde (1996), se reporta la presencia de tres fracciones de glicopeptidolípidos apolares involucrados en la respuesta protectora, estudio que se realizó solamente con la cepa de referencia TMC 5135 de M. habana. Por lo expuesto anteriormente, en este trabajo se realizó un estudio quimiotaxonómico con las 33 cepas de M. habana y la cepa de referencia M. habana TMC 5135 (pertenecientes a la colección de laboratorio de los autores, única en el mundo) y dos serotipos de M. simiae, con el objetivo de conocer la composición de diferentes moléculas importantes en la caracterización de especies micobacterianas (especialmente las fracciones de glicopeptidolípidos [GPL]) y comprobar si las 33 cepas de la colección mantenían el patrón reportado con anterioridad para tres glicopeptidolípidos apolares. Materiales y Métodos Se analizaron y compararon diferentes componentes lipídicos de 34 cepas de M. habana pertenecientes a la colección, con los de dos cepas de M. simiae Para este estudio se utilizaron las siguientes técnicas: cromatografía en capa delgada unidimensional y bidimensional, cromatografía en capa delgada de alta resolución, cromatografía de gases, cromatografía de gases acoplada a espectrometría de masas, cromatografía de gases acoplada a masas con bombardeo electrónico y cromatografía líquida de alta resolución. Los componentes estudiados fueron: ésteres metílicos de ácidos grasos, alcoholes, patrones de ácidos micólicos, glicolípidos y GPL presentes en dichas especies. Sobre la base de los resultados obtenidos, se estableció una comparación cualitativa en busca de posibles diferencias y semejanzas entre las especies estudiadas. Resultados Los patrones cromatográficos de ésteres metílicos de ácidos grasos, ácidos micólicos y glicolípidos en las dos especies fueron iguales. En estas cepas no se encontró la presencia de alcoholes y los productos de pirólisis fueron hasta el carbono C26. Sin embargo, en el estudio de GPL micobacterianos de las cepas de M. habana de la colección, se encontraron resultados no reportados previamente. Al analizar y comparar los patrones cromatográficos de las fracciones de GPL de las diferentes cepas de M. habana y M. simiae, se encontraron diferencias significativas, lo que permitió diseñar para M. habana cuatro modelos o patrones según la composición de GPL presentes no reportados hasta el momento. Además, se describió la presencia de un nuevo glicopeptidolípido. Esto constituye un hallazgo científico importante para esta especie micobacteriana cubana seleccionada como posible cepa vacunal y, a la vez, estos resultados podrán ser utilizados para elaborar nuevas estrategias para la selección de la cepa vacunal. Los GPL micobacterianos pueden ser utilizadas como control en la elaboración del futuro inmunógeno. Tabla. Modelos de glicopeptidolípidos diseñados para las cepas de M. habana y número de cepas agrupadas en cada uno de los modelos propuestos.
Mh1: Modelo habana 1, Mh2: Modelo habana 2, Mh3: Modelo habana
3, Mh4: Modelo habana 4 Bibliografía Mederos LM, et al. A chemotaxonomic study of 32
strains of Mycobacterium habana. Revista Española de
Quimioterapia 1997;10 (Sup 2). Copyright 1999 Elfos Scientiae
Biotecnologia Aplicada 1999; Vol. 16 No. 3, pp 182-184 Factores tóxico-nutricionales y neuropatía epidémica. Programa de investigaciones "SECUBA" Rafael Pérez Cristiá,1 Pedro Fleites
Mestre,1 Tomás Verdura Barrios,2 1Centro Nacional de Toxicología. Introducción Durante el período 1994-1997 se desarrolló el programa de investigaciones "SECUBA", con la colaboración de equipos de investigadores cubanos y franceses. Este programa fue propuesto en el Taller Internacional sobre Neuropatía Epidémica (NE), celebrado en julio de 1994, para dar continuidad a las investigaciones sobre la hipótesis tóxico-nutricional como causa de esta enfermedad. El objetivo de este programa fue estudiar los diferentes factores de riesgo tóxico-nutricionales asociados con la NE y su posibles variaciones estacionales en el transcurso de un año. Materiales y Métodos La población estudiada incluyó voluntarios masculinos sanos con edades entre 30 y 50 años: 100 fumadores y 100 no fumadores del municipio La Lisa, Ciudad de La Habana; 68 fumadores residentes en el municipio Caimanera, Guantánamo (municipio no afectado por la epidemia); y 51 enfermos de NE de la provincia Pinar del Río. Todos fueron sometidos previamente a examen clínico para descartar la presencia clínica o subclínica de NE en los sanos y para confirmar el diagnóstico de NE en los enfermos. Se realizaron encuestas de salud, hábitos de vida (hábitos tóxicos como el tabaquismo o exposición a tóxicos ambientales, entre otros) y hábitos nutricionales (durante una semana). De igual forma, se tomaron muestras de sangre y orina para el análisis de 50 marcadores biológicos (de estrés oxidativo, metabolismo proteico y energético, minerales y oligoelementos, vitaminas y otros parámetros). Los aportes de los alimentos y la cobertura de necesidades nutricionales fueron analizados mediante la base de datos NUTRISIS según los requerimientos de la población cubana. Con la información obtenida se confeccionó una base de datos y un programa en Access (versión 2.0) denominado EPIBASE. Los análisis estadísticos se efectuaron paralelamente por los grupos de trabajo cubano y francés según la estrategia establecida, con la ayuda de los programas estadísticos SAS (versión 6.11) en Unix y NCSS (versión 6.0). Se realizaron análisis univariados y multivariados (regresión logística) para todas las variables y grupos de estudios. Resultados y Discusión Los resultados principales de las investigaciones del Programa SECUBA señalan un incremento del estrés oxidativo en la población estudiada, expresado por valores elevados en los índices de peroxidación lipídica (TBARS), un estado deficitario de vitamina B2, y baja disponibilidad de carotenos, fundamentalmente de a y b -caroteno. Estos indicadores variaron en los cuatro períodos del estudio, de los cuales el segundo resultó el menos desfavorable (julio/95), con valores más bajos de TBARS, mayor ingestión de carotenos y valores séricos más altos de b -caroteno. Por el contrario, los niveles más críticos fueron encontrados en el tercero (octubre/95), con valores significativamente más altos de TBARS, bajos en la ingestión de carotenos y bajos de b -caroteno sérico, en comparación con el segundo. Otros factores presentes son el déficit de energía (por aporte insuficiente de grasas) y un fondo de carencia y subcarencia de vitaminas E, B1, B12, y ácidos grasos y calcio. Se comprobó que el hábito de fumar agudiza el déficit de carotenos y vitamina B2, sin diferencias significativas en estos aportes por la alimentación entre fumadores y no fumadores. Los enfermos de NE, en relación con los voluntarios de La Lisa, se caracterizaron por una menor defensa antioxidante asociada a niveles más bajos de a -caroteno, glutatión-peroxidasa y albúmina, y una mayor disfunción del metabolismo energético, con niveles más elevados de 3-hidroxibutirato. Esta condición fue desarrollada por un mayor desequilibrio nutricional correspondiente a un menor aporte dietético. Los factores de protección contra NE, al comparar los voluntarios de Caimanera con los de la Lisa, se relacionan con una mejor defensa antioxidante dada por mayores niveles séricos de b -caroteno y selenio, mejor estatus de vitamina B2 y mejor funcionamiento del metabolismo energético, expresado por niveles significativamente más bajos de 3-hidroxibutirato, y desde el punto de vista nutricional por un mayor aporte de grasas, proteínas, calcio y vitamina B2. En resumen, en este estudio se pudo comprobar un desbalance entre los factores antioxidantes y prooxidantes en detrimento de los primeros, como consecuencia de un desequilibrio nutricional que se agudiza en el caso de los enfermos de NE. Esto dio lugar a un incremento del estrés oxidativo por una deficiente defensa antioxidante. El consumo inadecuado de nutrientes antioxidantes por los malos hábitos dietéticos, asociado a desequilibrios en el aporte de otros, tiene el mayor peso en este desbalance y se caracteriza por una variación estacional. El programa SECUBA permitió poner en evidencia la presencia de un riesgo general por incremento del estrés oxidativo en las poblaciones estudiadas tanto sanas como afectadas por NE, y la importancia de adecuados aportes y niveles biológicos, entre otros, de nutrientes antioxidantes como protectores contra la NE de los efectos del estrés oxidativo. Los resultados del programa SECUBA demuestran la participación del estrés oxidativo y del déficit de antioxidantes en la patogenia de la NE, lo que se corresponde con los mecanismos fisiopatológicos y los cambios morfológicos característicos de las neuropatías tóxico-metabólicas o por déficit nutricional. Todo lo anterior permite considerar a la neuropatía epidémica como una enfermedad multifactorial desencadenada por factores tóxico-nutricionales y vinculada al estrés oxidativo. El programa SECUBA aporta un sistema validado para la vigilancia y control del balance antioxidantes-prooxidantes (estrés oxidativo) sobre poblaciones. Para esto dispone de un conjunto de marcadores biológicos integrados a un protocolo de información sobre salud, hábitos de vida y alimentación. Se desarrolló un programa automatizado (EPIBASE) que permite la estandarización de los diferentes estudios que se efectúen en este sentido, lo que constituye una herramienta de valor para abordar esta temática. Por otra parte, estos resultados permiten alertar sobre el posible incremento de enfermedades crónicas vinculadas al estrés oxidativo y al déficit de antioxidantes, y permiten desarrollar medidas de prevención vinculadas a un mayor consumo de alimentos ricos en nutrientes antioxidantes, lo que incidiría en la mejoría del cuadro de salud y en la calidad de vida de nuestra población. Se reconoce la colaboración de C Campa, G Sierra, N González, A Maciques, Instituto "Finlay"; T Hernández, T Conill, S Suárez, E Rodríguez, CENATOX; I García, L Ledesma, A Galán, IFAL; M Santín, C Serrano, Dirección Nacional de Epidemiología, MINSAP; C Santos, N Pérez, CIMEQ; L Regal, I Mesa, Sectorial de Salud La Lisa; H Gautier, Instituto de Hematología e Inmunología; M Morales, A Sotolongo, Centro Provincial de Higiene y Epidemiología de Pinar del Río; BE Eliot, M Sánchez, Hospital "Abel Santamaría", Pinar del Río; ME Montoya, Centro Provincial de Higiene y Epidemiología de Guantánamo. Se agradece, además, la colaboración de los técnicos O Toste, A Velázquez, J Delgado, A Mesa, CENATOX. También se reconoce la labor de los especialistas franceses que trabajaron en esta investigación: V Azais-Braesco, D Bauchart, JP Charconac, E Combe, Drr N, P Grolier, P Pichenot, F Rocca F, JC Tressol, del INRA, Theix; JF Dartigues, INSERM U330; A Favier, Facultad de Farmacia, Grenoble; y JC Renversez y MJ Richard, de Bioquímica A CHU, Grenoble. Bibliografía Verdura T, Arnaud J, Pérez Crisitá R, Tressol
JC, Fleites P, Chassagne M, et al. Trace element status in
Cuba. Relationships with Epidemic Neuropathy through SECUBA
Protocol. En: Neve, et al, editors. Therapeutic Uses of
Trace Elements. New York: Plenum Press; 1996. p.391-4. Copyright 1999 Elfos Scientiae
Biotecnologia Aplicada 1999; Vol. 16 No. 3, pp 184-186 Obtención de plantas transgénicas de caña de azúcar (Saccharum officinarum L) mediada por Agrobacterium tumefaciens: una nueva metodología para la transformación genética de esta gramínea Gil A Enríquez-Obregón,1 Roberto I
Vázquez-Padrón,1 Ariel D Arencibia,1 1Centro de Ingeniería Genética y
Biotecnología. AP 6162, La Habana, Cuba. Introducción La caña de azúcar (Saccharum officinarum L.) es uno de los cultivos más extendidos en las regiones tropicales y subtropicales. La industria azucarera y la obtención de productos químicos como el furfural, dextranas y alcohol, dependen de las zafras. Además, los subproductos del proceso de producción de azúcar representan una valiosa fuente para la alimentación animal. La ingeniería genética ha permitido obtener variedades de este cultivar con características no presentes en su fondo genético. Mediante electroporación de células intactas y biolística, se han obtenido plantas transgénicas de caña de azúcar resistentes al ataque del bórer (Diatraea saccharalis) [1] y al herbicida BASTA® [2], respectivamente. Las metodologías de transformación que utilizan Agrobacterium tumefaciens para transferir ADN a las células vegetales, ofrecen la ventaja de transferir genes al genoma vegetal en forma limitada, con reproducibilidad y sin necesidad de utilizar equipos costosos. En este trabajo se desarrolló una metodología para introducir, de manera estable, genes foráneos en células de caña de azúcar mediante la utilización de A. tumefaciens. Como explantes se utilizaron meristemos y callos de la variedad comercial Ja60-5, tratados previamente con una mezcla de compuestos antioxidantes. Este tratamiento disminuyó la muerte celular, lo que mejoró la competencia del tejido vegetal y, de este modo, la frecuencia de transformación genética. Procedimientos y Resultados Construcción del vector binario pGT GUSBAR Para el establecimiento de las condiciones de transformación se utilizó la cepa At 2260 transformada con el plásmido pGT GUSBAR, que porta los genes reporteros uidA (codifica la b -glucuronidasa [GUS] de E. coli) y bar (que codifica la fosfinotricina-acetiltransferasa), bajo señales apropiadas para la regulación de la transcripción en plantas monocotiledóneas [3]. Evaluación del efecto de los compuestos antioxidantes sobre la necrogénesis de los explantes y callos de caña de azúcar Para obtener explantes de caña de azúcar con índices de necrogénesis bajos en la superficie de las zonas de corte, y con una elevada competencia para ser transformados por At, se evaluaron tres compuestos antioxidantes: el ácido ascórbico, la cisteína y el nitrato de plata, durante el estadio de precocultivation (Tabla 1). Se tomó como criterio de viabilidad celular la fracción meristemática que permaneció sin teñir frente al reactivo Evans Blue. Cada compuesto se probó en dos concentraciones diferentes de manera individual en medio líquido P+5 [4]. En todos los casos se observó una disminución en la necrogénesis del explante después de 60 h de incubación (Tabla 1). Tabla 1. Efecto de los tratamientos antioxidantes sobre el crecimiento de A. tumefaciens, la viabilidad del tejido meristemático y la calidad del callo.
1 Compuestos antioxidantes ensayados en medio P+5 líquido.2Promedio de tres experimentos independientes de determinación de DO620 en cultivos de A. tumefaciens en medio YEB a 28 ºC por 20 h. 3Los por cientos de viabilidad representan el promedio de diez determinaciones independientes realizadas al estereoscopio en la fracción del explante que permanece sin teñir luego de tinción con el colorante Evans Blue, versus el área total del explante. 4La calidad del callo fue considerada en cada caso por inspección visual y se clasificaron como: Tipo II. Callo friable amarillo con alta capacidad de embriogénesis Tipo III. Callo blando, blanco y opaco no embriogénico 5Medio P+5 sin compuestos antioxidantes usado como control negativo. A pesar de que la hipersensibilidad del explante disminuyó en cada tratamiento, los mejores resultados se obtuvieron cuando se utilizó la mezcla de estos compuestos. En este caso, la necrosis en el meristemo fue inhibida hasta en 90%. Este resultado evidencia la posibilidad de sinergismo entre los antioxidantes para disminuir la respuesta de hipersensibilidad en la célula vegetal. El efecto de los antioxidantes sobre la formación de los callos fue estudiada utilizando explantes meristemáticos. Los tratamientos AAS1, CIS1, Ag1 y AO permitieron la formación de callos amarillentos y friables con alta capacidad de regeneración, similares a los obtenidos con los explantes sin tratar (Tabla 1). Cuando se utilizaron altas concentraciones de los compuestos antioxidantes, los callos generados resultaron opacos y blandos (tipo III) con una baja capacidad de regeneración. Efecto de los compuestos antioxidantes sobre la interacción del tejido vegetal con A. tumefaciens El tratamiento denominado AO fue escogido para estudiar el efecto de los antioxidantes en la transferencia de genes mediada por At. Como criterio para medir la eficiencia de transformación, se tomó la fracción de callos GUS-positivos y resistentes a BASTA. Igualmente, se evaluaron explantes de plantas cultivadas in vitro (IN) o procedentes del campo (PC) tratadas con la mezcla AO. Ciento por ciento de los explantes infectados fueron GUS-positivos cuando a los medios de cultivo se les adicionó compuestos antioxidantes (IN1, PC1) durante la pre- y la cocultivación. En contraste con este resultado, la eficiencia de la infección fue menor cuando se utilizaron medios simples. No se detectó actividad GUS en los explantes no infectados. Los explantes pertenecientes a los tratamientos IN1 y PC1, mostraron una tasa de formación de callos de 15% y 25%, respectivamente. La formación de callos fue muy inferior (0-13%) en el resto de las variantes. El tejido meristemático no infectado no proliferó en el medio selectivo. La presencia del ADN foráneo en los callos resistentes a BASTA fue analizada por PCR e indirectamente por un ensayo histoquímico de actividad GUS. Observamos una mejor callogénesis a partir de las plantas crecidas en condiciones de campo en comparación con las procedentes de plantas in vitro (Tabla 2). Estos resultados son similares a los reportados en uva (Vitis vinifera) [5] para la que se observó una reducción en la necrosis de los callos y un aumento de los eventos de transformación con la utilización de varios compuestos antioxidantes. Tabla 2. Efecto de los compuestos antioxidantes en la transferencia génica de A. tumefaciens a la caña de azúcar.
1 IN: explantes de plantas de caña de azúcar cultivadas in vitro; PC: explantes de plantas de caña de azúcar crecidas en condiciones de campo.2La actividad GUS fue evaluada dos días después de la cocultivación en diez muestras seleccionadas al azar, mediante la utilización del procedimiento histoquímico estándar. 3El total de explantes infectados por Agrobacterium aparece entre paréntesis. 4Análisis por PCR del ADN genómico de un callo para cada tratamiento. ND: muestras no disponibles. Transformación de meristemos Los cuatro protocolos de transformación ensayados se muestran en la Tabla 3. Los callos provenientes de los meristemos transformados fueron cortados y colocados en medio selectivo por tres meses con subcultivación cada mes, lo que permitió la obtención de callos friables con alta capacidad de regeneración. Tabla 3. Comparación de cuatro protocolos de transformación.
De manera similar se infectaron callos friables, con la obtención de actividad transitiva y proliferación celular en medios selectivos con higromicina [6]. Posteriormente, se regeneraron plantas a partir de los callos resistentes a BASTA o higromicina. Análisis molecular y bioquímico del material transformado Se seleccionaron tanto los callos como las plantas resistentes a BASTA que fueron GUS-positivos y mostraron la presencia del ADN foráneo en el genoma mediante Southern blot. Ensayo de resistencia al herbicida BASTA en condiciones de invernadero y de campo El crecimiento y la morfología de las plantas transgénicas fue similar con respecto a las no transformadas. Todas las plantas fueron rociadas con una dosis promedio de 2,5 g/L de BASTA. Las plantas no transformadas, así como las plantas con bajos niveles de expresión del gen bar, fueron afectadas seriamente. Al finalizar las pruebas bajo condiciones de invernadero, se seleccionaron 360 plantas para otras pruebas en condiciones de parcela. Las plantas transgénicas resistentes al herbicida, así como las plantas no transformadas (n = 40) fueron adaptadas a las condiciones de campo. Finalmente, se seleccionaron 112 individuos altamente resistentes a BASTA, los cuales procedían de 59 callos transformados. Todas las plantas no transformadas murieron (Tabla 4). Tabla 4. Resistencia a BASTA en condiciones de campo.
La novedad de este trabajo radica en el estableciemiento de una nueva metodología para la transformación de la caña de azúcar con una eficiencia mayor que la de los métodos físicos descritos, así como la generación de plantas transgénicas resistentes al herbicida BASTA. Referencias 1. Arencibia A, Vázquez R, Prieto D, Téllez P,
Carmona E, Coego A, et al. Transgenic sugarcane plants
resistant to stem borer attack. Mol Breeding 1997;3:247-55. Copyright 1999 Elfos Scientiae
Biotecnologia Aplicada 1999; Vol. 16 No. 3, pp 186-189 Desarrollo de las aplicaciones de los marcadores moleculares en el mejoramiento de la caña de azúcar en Cuba María T Cornide, Orlando Coto, Eduardo
Canales, Antonio Sigarroa, Jesús E Sánchez, Dpto. de Biotecnología de las Plantas. Centro Nacional de
Investigaciones Científicas. Ave 25 y 158, Cubanacán,
AP 6990. Ciudad de La Habana. Cuba. Introducción La caña de azúcar es un híbrido complejo de naturaleza autopoliploide, apareamiento cromosómico bivalente con un sistema de reproducción preferentemente alógamo, y con capacidad de multiplicación vegetativa. El conocimiento y manejo de estas características determinan la eficiencia de los estudios genéticos y de los programas de mejoramiento. Se considera que las principales limitaciones del mejoramiento cañero tradicional, vía casi exclusiva de obtención de variedades nuevas, son las siguientes: 1. La estrecha base genética empleada con estos fines, tanto por la escasa representación de formas, como por la elevada consanguinidad entre los híbridos comerciales usados como progenitores. 2. La baja heredabilidad de los caracteres evaluados y el efecto de competencia en la población de posturas provenientes de semilla botánica durante la primera etapa del esquema, en la que se aplica la selección más rigurosa. 3. La elevada contribución del componente genotipo-ambiente en la varianza fenotípica total de los principales criterios de selección en las etapas clonales, lo que aumenta la duración del programa y obliga a la realización de ensayos replicados para localidades, cepas y épocas de siembra y cosecha. Por estas razones, el empleo de los marcadores del polimorfismo del ácido desoxirribonucleico o marcadores moleculares (MM), junto a los métodos tradicionales de mejoramiento, constituye una vía poderosa para acelerar el conocimiento genético y aumentar la eficiencia del mejoramiento en la caña de azúcar. El presente trabajo abarca un conjunto de resultados teórico-prácticos que permiten identificar y elegir con más eficiencia la variabilidad genética para la obtención de nuevas variedades, y para la aplicación de esta tecnología de gran impacto, estandarizada para cuatro sistemas de MM (RFLP, AFLP, RAMP y RAPD), en la búsqueda de marcadores para la selección en la caña de azúcar y en otros cultivos de interés. Las investigaciones se realizaron en el marco de un proyecto CITMA del PNCT de Biotecnología Agrícola y de un proyecto CNIC financiado por el Consejo de Estado (1996-1998). Caracterización molecular de materiales de interés en el mejoramiento Caracterización del germoplasma básico utilizado en el mejoramiento de la caña De los 15 clones ancestrales de mayor participación en la genealogía de las variedades cubanas, ocho son también germoplasma básico de los programas de mejoramiento de la Florida y Louisiana [1] y, al igual que en estos casos, los clones de S. officinarum de mayor participación son: Black Cheribon, Chunnee, Bandjarmasin Hitam, Loethers, Lahaina y Fiji. Por primera vez en Cuba se estudió la diversidad genómica (por RFLP) de 27 clones de fundación de las variedades modernas de la caña de azúcar (S. officinarum, S. spontaneum, S. robustum, S. barberi, S. sinense y Erianthus spp.), que están entre los más usados como germoplasma básico en el programa de nobilizaciones [2]. Las combinaciones de RFLP mediante el empleo de sondas heterólogas de maíz, resultaron de utilidad en la identificación de materiales de caña de azúcar con diferente grado de relación y complejidad genéticas, lo que confirmó resultados previos sobre la sintenia entre estas especies. Se reportaron diez bandas raras con presencia preferente o exclusiva en algunos clones de Erianthus spp. (B7, E13, H9), S. spontaneum (C2, C9, EC3); S. officinarum (EC8, EC9, H4) y S. robustum (EC7) de futura utilidad en trabajos de clasificación taxonómica. Se encontró una banda común sólo presente en los clones de S. robustum y S. officinarum (C14). No se encontraron bandas exclusivas de los clones de estas especies, ni en los clones de S. barberi o S. sinense que pudieran apoyar el probable origen secundario de estas últimas. Por esta razón, la diversidad común entre especies, revelada por los RFLP del trabajo, tiene un origen más amplio. Los miembros de Erianthus spp. pueden ser identificados con el uso de las bandas: B7 (UMC81-BamHI, 4,4 kpb); H9 (UMC39-HindIII, 2,9 kpb) y H13 (UMC81-EcoRI, 3,4 kpb) [3]. Se desarrolló una metodología para el procesamiento de la información con vistas a la determinación de los fondos genéticos. Se demostró la mayor diversidad de los clones S. spontaneum en relación con las de otras especies. Se encontraron dos genofondos en los clones de S. spontaneum y 3 subgrupos en los de S. officinarum, lo que aumenta la eficiencia de la elección de este material como progenitores masculinos y femeninos para la nobilización. Quedó demostrada la superioridad de la caracterización integral con la incorporación de la diversidad genómica (RFLP) para la recomendación de formas originales, la cual resulta difícil y poco confiable por métodos tradicionales cuando se realiza sólo. Caracterización de nuevas formas originales La reducida representación de formas originales limita la eficiencia del avance generacional para obtener nuevos progenitores en la caña de azúcar, en especial en Cuba. Para ampliar la colección de formas originales, y con esto la eficiencia del programa de avance generacional para obtener nuevos progenitores en la caña de azúcar, se caracterizaron por RFLP 35 clones nuevos (8 de Miscanthus spp. y 27 de S. spontaneum) colectados en Laos junto a clones de S. spontaneum y S. officinarum representativos de los genofondos identificados en el material de fundación. Se evidenció la presencia de tres genofondos nuevos de S. spontaneum no coincidentes con los del trabajo anterior, lo que demuestra la eficiencia de la colecta efectuada. Esto permitió ampliar la variabilidad de partida de nuestro mejoramiento cañero con mayor precisión y rapidez. No se detectaron duplicados [4]. Los clones clasificados como Miscanthus spp. están en estudio con la utilización de sondas específicas del género para corroborar su clasificación taxonómica. Caracterización de híbridos comerciales Por primera vez, 45 híbridos y somaclones comerciales cubanos fueron caracterizados por su diversidad genómica (35 combinaciones de RFLP y 18 de RAPD) y fenotípica (5 caracteres), lo que apoya su elección como progenitores [5, 6]. Estos resultados permiten auxiliar la elección de estos clones con la incorporación de la diversidad de su genofondo al análisis tradicional, basado sólo en caracteres fenotípicos para aumentar las posibilidades de obtener genotipos transgresivos. Se evaluó la diversidad genética de 11 clones del programa de selección recurrente de madurez temprana, provenientes de cinco cruces biparentales y dos policruces, y escogidos por su comportamiento azucarero promisorio en relación con sus progenitores y con un grupo de variedades testigos. Se utilizaron sondas genómicas de caña de azúcar (SSCIR) previamente escogidas por su alto polimorfismo y por su calidad autorradiográfica. Los estimados de las distancias entre cada par de clones permitieron conocer, de forma preliminar, la efectividad de esta etapa del programa. Para su identificación, se escogieron 15 bandas polimórficas de SSCIR194-BamHI. Los RFLP diferenciales hacen posible una rápida elección de las combinaciones enzima-sonda (CES) que se emplearán en los análisis de las progenies con fines de mapeo futuro. Desarrollo del uso de MM como descriptores En los últimos años ha habido un uso creciente de los MM en la conservación de recursos fitogenéticos, la cual se espera que aumente y se diversifique en apoyo a la protección legal internacional de nuevos productos biotecnológicos. Los resultados de los autores demostraron la posibilidad de emplear los patrones de polimorfismo de los MM como descriptores moleculares en la identificación de los materiales estudiados, teniendo en cuenta su reproducibilidad, la facilidad de evaluación, el nivel de polimorfismo, el costo y la representatividad de las bandas, esta última expresada por su asociación con genofondos, categorías taxonómicas o características de interés en el mejoramiento. Los descriptores propuestos se incorporaron a la base de datos del banco de germoplasma de la EPICA, en Camagüey, y está aprobada su inclusión en la nueva versión del sistema automatizado del banco de germoplasma de caña de azúcar (BANCOGER). Con la realización de esta propuesta, se inició en el país el uso de descriptores moleculares en la actividad conservacionista de los recursos fitogenéticos. Se dispone de los patrones de polimorfismo molecular (RFLP, AFLP, RAMP) recomendados en los estudios de diversidad, organizados en cinco secciones: clones de fundación del complejo Saccharum (27); nuevos clones de la expedición Cuba-Laos (35); híbridos comerciales cubanos de interés en el mejoramiento (41); clones promisorios del programa de recurrencia de la madurez temprana (11); y somaclones comerciales y clones transgénicos promisorios (8). Los patrones que se van a emplear pueden ser seleccionados directamente por cualquier usuario. En el mundo la metodología garantiza una alta confiabilidad al facilitar la elección por el usuario de las bandas con mayor valor discriminatorio, las cuales diferencian subgrupos o fondos genómicos de los clones de la Sección, y de las bandas raras o únicas que identifican los clones dentro de cada subgrupo. Diversidad genética. Predicción del comportamiento familiar. Aplicación del BLUP En la caña de azúcar se ha demostrado la eficiencia de la selección familiar y del valor predictivo del potencial de una familia expresado a través de las medias de los caracteres más importantes en la etapa de posturas. Su estimación se realiza en costosos estudios de progenies que evalúan un número reducido de progenitores; por lo tanto, un método predictivo de las medias familiares basado en las características parentales, facilitaría la realización de la selección familiar. Se compararon tres indicadores de diversidad entre los progenitores (distancia genealógica, DG; distancia multivariada, DI; distancia genómica por RFLP, DP) por su valor predictivo de las medias familiares, con cinco indicadores basados en estudios de progenies para el brix y los componentes del peso del tallo en una etapa temprana del esquema de selección [7]. El estudio se realizó en cruces factoriales de diez progenitores cubanos importantes. Se introdujo el método del mejor predictor lineal insesgado (BLUP) para potenciar el uso de las distancias en la estimación de las medias familiares. Para ello, los autores modificaron el modelo de Bernardo [8] según cinco variantes de la matriz de varianza-covarianza. Se demostró: • La utilidad de emplear el polimorfismo molecular revelado por
RFLP para evaluar la verdadera diversidad genómica entre dos
genotipos de caña de azúcar, independientemente de su
origen idéntico por descendencia o similar en estado, en
relación con el coeficiente de coancestría y la
distancia fenotípica. Estudios metodológicos Se realizó un conjunto de estudios para perfeccionar la representación gráfica de la diversidad de MM para determinar fondos genómicos en la caña de azúcar. Se reportó la presencia y magnitud del sesgo de los estimados de distancias debido a variantes RFLP comunes entre genotipos no emparentados (dAB). Sin embargo, la sobreestimación de los estimados de similitud genética por este sesgo en los materiales caracterizados, no resultó importante en la determinación de los genofondos. Esto se debe a que la resolución de los análisis multivariados en la determinación gráfica de los genofondos de RFLP, dependió de la unidad de polimorfismo considerada (bandas individuales o patrones de bandas de cada CES) y no de la fuente de sesgo anterior o de la presencia de variantes parcialmente correlacionadas entre diferentes CES revelados con una misma sonda. A estos efectos se recomienda considerar como variantes las bandas polimórficas individuales reveladas de las CES previamente seleccionadas por su alto polimorfismo y su facilidad de conteo. La confiabilidad de la determinación de los genofondos por la metodología propuesta por los autores, fue confirmada al mantenerse el agrupamiento de clones por los métodos multivariados más usados. Estudios genético-estadísticos y paquete especializado de programas computadorizados de apoyo Se diseñó y validó un paquete de programas (mat-gen) (Registro 02707-2707) para la determinación de las relaciones entre genotipos con variantes binarias por 17 indicadores de asociación de tres tipos de ponderación [9]. Este paquete facilita los estudios de diversidad molecular, al agrupar las operaciones más frecuentes que el usuario realiza. Como novedad, se brinda la opción de determinar y corregir el sesgo por variantes comunes entre individuos no emparentados, mediante tres tipos de indicadores y la comparación de los estimados de asociación mediante la técnica de Jackknife, de gran utilidad especialmente en estudios de poliploides. Se confeccionó un software para el mapeo en poliploides (POLIMAP), validado con datos de caña de azúcar procedentes del CNIC [10]. Los mapas permiten la búsqueda de MM para la selección en la caña y en otros cultivos tropicales. Los autores calcularon las proporciones de segregación y desarrollaron las fórmulas de estimación para determinar: la dosis del marcador en alopoliploides y autopoliploides sin doble reducción (2x-16x); las relaciones de ligamiento y su configuración (método de dos puntos) con fragmentos de 1-3 dosis; las autofecundaciones no controladas; el tipo y nivel de ploidía (2 métodos); y la fracción de recombinación de las 11 configuraciones más informativas. Agradecimientos Loa autores hacen constar la valiosa participación del Ing. Julio Vallina, los doctores Rolando González y Antonio Chinea, la Ing. Ibis Jorge, investigadores del Instituto Nacional de Investigaciones de la Caña de Azúcar (INICA), el Lic. Guillermo Selman Housein, investigador del Centro de Ingeniería Genética y Biotecnología, Aleida Veitía y Rusbel Michel, técnicos del CNIC, y Belkis Pino, técnica del Centro Nacional de Sanidad Agropecuaria (CENSA). Además, los autores agradecen a los doctores Guillermo Gálvez Rodríguez, Roberto de Armas y Rodobaldo H. Ortíz los criterios especializados brindados durante la proyección y ejecución de estos trabajos, y a los licenciados Dannelle Echemendía y Rafael Fando el apoyo brindado en el trabajo de laboratorio. Bibliografía Arencibia A, Carmona E, Cornide MT, Wang G, Castiglione S,
Cuzzoni E, et al. Field performance end genome analysis of
resistant transgenic sugarcane (Saccharum spp.) plants.
Transgenic Research . Abreviaturas AFLP: polimorfismo de la longitud de fragmentos amplificados
(Amplified Fragment Length
Polymorphism) Copyright 1999 Elfos Scientiae
Biotecnologia Aplicada 1999; Vol. 16 No. 3, pp 190-191 Desarrollo de la crioconservación de germoplasma vegetal en Cuba María T González-Arnao,1,2 Caridad Urra,2 Marcos Martínez,3 María de los Ángeles Torres,4 Manuel Martínez,5 Tomás Moreira,2 Rodobaldo Ortiz5 1Dpto. Biología Vegetal. Facultad de Biología.
Universidad de La Habana. Calle 25 No. 455, entre I y J, Vedado,
Ciudad de La Habana, Cuba. Fax: (53-7) 32 1321; E-mail: arnao@fbio.oc.uh.cu
Introducción Este trabajo resume el aporte científico de Cuba al desarrollo de tecnologías para la preservación segura y a largo plazo de germoplasma vegetal de origen tropical. La crioconservación consiste en el almacenamiento a temperaturas ultrabajas, preferiblemente la del nitrógeno líquido (-196 °C). Actualmente, este método es el único factible para la preservación de material biológico de interés durante períodos teóricamente ilimitados. Las muestras crioconservadas se mantienen en el reducido espacio de termos y están exentas de los inconvenientes climáticos, biológicos y geográficos del hábitat natural. Se definen metodologías para crioconservar varias formas cultivables in vitro (ápices, callos embriogénicos y brotes meristemáticos) de cinco cultivos de importancia económica, alimentaria y biotecnológica como la caña de azúcar, el plátano, el café, la piña y los cítricos. Los métodos desarrollados para los ápices de caña, piña y cítricos, constituyen los primeros reportes en el ámbito internacional. Descripción del resultado Con el programa de investigaciones sobre la caña de azúcar, se logró establecer metodologías de crioconservación para ápices [1-4] y callos embriogénicos [5], que permiten preservar los recursos genéticos de este cultivo y las líneas celulares empleadas en los programas de mejoramiento y semilla artificial, respectivamente. Los estudios abarcaron la extensión exitosa de dichas tecnologías a un gran número de variedades de diferente complejidad, adaptación in vitro y diversidad geográfica. Se evaluó la estabilidad genética en condiciones de campo mediante métodos bioquímicos, histológicos y a través de la revisión de numerosas características morfológicas, lo que permitió comprobar que el almacenamiento prolongado (hasta catorce meses) en nitrógeno líquido, no indujo modificaciones genéticas ni afectó los niveles de viabilidad y la obtención de nuevas plantas [4]. En el cultivo plátano, el comienzo de las investigaciones requirió la obtención, por cultivo in vitro, de una estructura nueva (brotes meristemáticos) más resistente al frío y a los tratamientos crioprotectores [6]. Se logró la crioconservación exitosa de tres genotipos en peligro de extinción pertenecientes al banco de germoplasma, y se realizaron los estudios de estabilidad genética en condiciones de campo. En el caso del café, las metodologías desarrolladas para callos embriogénicos [7] permitieron conservar el material de partida de los trabajos de mejoramiento. Para los cítricos, se definió un protocolo de crioconservación para callos embriogénicos mediante el empleo de un cultivar comercial de naranja dulce [8]. También se estableció una metodología para conservar los ápices de diferentes especies patrones [9, 10] y actualmente se investiga con material de plantas adultas. En el caso de la piña, el método de crioconservación establecido se extendió con éxito a diferentes variedades [11] y se aplica a mayor escala en la actualidad. Importancia del trabajo Los aportes tecnológicos realizados para el desarrollo de la crioconservación de germoplasma vegetal tiene un gran impacto científico nacional e internacional. Por otra parte, el trabajo tiene un alto valor estratégico y social, ya que la implantación de estos métodos permite conservar especies que se encuentren en peligro de extinción, contribuye a eliminar las pérdidas por condiciones climáticas adversas u otras causas, y asegura la preservación de la biodiversidad. Desde el punto de vista económico, es posible reducir el número de réplicas en el banco de germoplasma en campo, con el consiguiente ahorro de superficies cultivables, mano de obra, etc. A escala de laboratorio, influye en la disminución de los gastos por concepto de compra de reactivos, consumo de electricidad y dedicación de personal especializado. Bibliografía González-Arnao MT. Cryopreservation of plant
germplasm using the encapsulation-dehydration technique. Technical
Bulletin IPGRI: Low temperature technologies; 1997. Referencias 1. González-Arnao MT, Engelmann F, Urra C, Lynch PT.
Crioconservación de meristemos apicales de plantas in
vitro de caña de azúcar mediante el método
de encapsulación -deshidratación.
Biotecnología Aplicada 1993;10:225. Copyright 1999 Elfos Scientiae
Biotecnologia Aplicada 1999; Vol. 16 No. 3, pp 191-193 Humanización de epitopos T en las regiones variables murinas de un anticuerpo que reconoce al receptor del factor de crecimiento epidérmico: obtención de inmunoglobulinas de menor inmunogenicidad Cristina M Mateo de Acosta del Río, Josefa
Lombardero Valladares, Ernesto Moreno, Rolando Pérez
Rodríguez Introducción Existen numerosas evidencias experimentales que apuntan a la relación existente entre el sistema EGF/EGF-R (factor de crecimiento epidérmico/receptor del factor de crecimiento epidérmico) y el cáncer, por lo que no es sorprendente que este sistema se haya convertido en un blanco atractivo para el diseño de nuevos medicamentos antitumorales. Diferentes grupos de trabajo, incluido el del colectivo de autores, han obtenido anticuerpos monoclonales bloqueadores de la señalización del receptor. Estos anticuerpos inhiben el crecimiento de tumores humanos en ratones atímicos. Sin embargo, los anticuerpos murinos han resultado demasiado inmunogénicos en humanos, lo que dificulta sus aplicaciones terapéuticas. Materiales y Métodos Se rediseñó el Anticuerpo Monoclonal R3 (AcM R3) como una inmunoglobulina humana IgG1, mediante la transplantación de los CDR (regiones determinantes de la complementariedad) del anticuerpo murino, en los marcos (FR) de las regiones variables humanas de las cadenas pesadas y ligeras Eu y Rei, respectivamente. Mediante la utilización de este sistema como modelo, se evaluó un nuevo método de humanización mucho más simple que el método tradicional, basado en el carácter de antígeno T-dependiente de las inmunoglobulinas y en la alta homología existente entre las inmunoglobulinas murinas y humanas. El método consiste en sustituir aquellos residuos murinos incluidos en los posibles epitopos T de los marcos de las regiones variables, por los que se encuentran en la misma posición de las inmunoglobulinas humanas de mayor homología, excepto aquellos residuos críticos en la conformación de los CDR. Los posibles epitopos T en las regiones VH (región variable de cadena pesada) y Vk (región variable de cadena ligera) del R3, fueron predichos por el programa AMPHI. Resultados y Discusión Se obtuvo un anticuerpo monoclonal reformado h-R3 neutralizante del EGF-R, con una Ki de 10-8 M, el cual ha sido objeto de seis solicitudes de patente. Para obtener este anticuerpo fue necesario clonar y secuenciar la región variable del AcM R3 murino, clonar las cadenas ligera y pesada murinas del mismo en vectores de expresión de células superiores, que tienen incluidas las regiones constantes humanas Ck (para la cadena ligera) y gamma 1 en el vector de expresión de la cadena pesada, con lo que se obtuvieron las construcciones genéticas quiméricas de las cadenas pesada y ligera del R3, es decir, las regiones variables murinas y regiones constantes humanas. Estas construcciones genéticas se introdujeron por transfección en células de mieloma NSO (mieloma no productor de inmunoglobulinas) y se expresó el anticuerpo quimérico R3, que fue capaz de reconocer el EGF-R. El anticuerpo humanizado se obtuvo mediante transplante de los CDR del anticuerpo murino, en los marcos de las regiones variables humanas de las cadenas pesada y ligera Eu y Rei, respectivamente. Este anticuerpo perdió la capacidad de unión al EGF-R. A partir del mismo, se construyó un anticuerpo reformado al que se le reintrodujeron residuos murinos en algunas posiciones de los marcos humanos. Para la construcción de este anticuerpo reformado, se hizo un análisis de los residuos de las regiones marcos que pudieran influir en la conformación que adoptan los CDR para el adecuado reconocimiento del antígeno y para tratar de mantener los mismos, para no afectar la zona de unión del anticuerpo a su antígeno. En el marco de estos estudios, se construyeron variantes de anticuerpos híbridos quiméricos y humanizados. Cuando las células NSO se cotransfectaron con un híbrido construido con la cadena ligera humanizada y la cadena pesada quimérica, el anticuerpo obtenido se comportó igual que el R3 murino. La combinación inversa, es decir, el híbrido construido entre la cadena ligera quimérica y la cadena pesada humanizada, se comportó igual que el R3 humanizado. Estos resultados indicaron que los cambios introducidos por la humanización de la cadena ligera no afectaron el reconocimiento del anticuerpo por su receptor, por lo que el trabajo genético de reintroducir algunos residuos murinos en los marcos humanos para recuperar la actividad biológica del anticuerpo, se concentró en la cadena pesada. Para recuperar la capacidad de inhibición de la unión del EGF a su receptor por el R3 humanizado, se hizo necesario introducir en los marcos humanos algunos residuos de aminoácidos que se encontraban en la misma posición en la inmunoglobulina murina original. A esta nueva construcción genética se le llamó "anticuerpo reformado". Como se explicó anteriormente, estos cambios se concentraron en la cadena pesada humanizada, manteniéndose intacta la cadena Vk humanizada. Estas mutaciones se realizaron en tres grupos diferentes: mutaciones en las posiciones 66-67, 75-76 y, por último, 93, considerando su localización en diferentes regiones y la influencia que pudieran tener en el sitio de combinación del antígeno. Se construyeron seis versiones del anticuerpo reformado R3, en las que uno, dos o tres de los grupos de residuos se mutaron para introducir en dichas posiciones los aminoácidos que aparecen en la secuencia del R3 murino en la misma posición (Tabla). Tabla. Región variable de la cadena pesada de los anticuerpos reformados de R3.
A: secuencia de ADN de la región VH del
anticuerpo humanizado. Los residuos murinos de los marcos de VH, LYS66 y ALA67 fueron reintroducidos porque se encuentran en la frontera entre CDR2 y FR3, y las mismas mostraron ser necesarias en la recuperación de la afinidad por el EGF-R en el anticuerpo 425, que presenta una alta homología con la región variable de la cadena pesada del R3. Estos cambios no resultaron importantes, puesto que el anticuerpo reformado recuperó muy poco la capacidad de inhibir la unión del EGF a su receptor. Se construyeron, además, anticuerpos reformados con SER75, THR76 y THR93 de origen murino. Las versiones de R3 reformado, R3-S75T76 y R3-T93, mostraron cambios importantes en la capacidad de inhibición de la unión del EGF-I125 a su receptor, con valores de Ki entre 1,34 y 1,59x10-8 M, mientras que las versiones R3-S75T76T93 o R3-K66A67S7 5T76T93, que incluyen los residuos SER75, THR76 y THR93, recuperaron la inhibición de la unión del EGF-I125 a EGF-R, con una Ki de aproximadamente 1x10-8 M. Este valor es muy cercano al que tiene el anticuerpo murino R3 (0,7-0,8x10-8 M). Esta versión de anticuerpo reformado fue la escogida para hacer los experimentos in vivo. Solo tres mutaciones en los marcos humanos por los correspondientes residuos de ratón, permitieron recuperar 80% de la afinidad por el antígeno. Este anticuerpo reformado resultó mucho menos inmunogénico en monos que los anticuerpos quimérico y murino correspondientes. Se describió por primera vez en la literatura la importancia del residuo 76 de la región variable de la cadena pesada en la conformación del sitio de unión del anticuerpo. Sólo existe otro anticuerpo recombinante neutralizante del EGF-R, el cual se encuentra en ensayos clínicos en estos momentos. El c225 es un anticuerpo quimérico con afinidad similar a h-R3 que en la actualidad se encuentra en desarrollo por Imclone Systems Inc., NY, y debe recibir aprobación en el año en curso por parte de la FDA para iniciar la fase III de ensayos clínicos en pacientes con cáncer de cabeza y cuello. El anticuerpo reformado h-R3, que en principio debe resultar menos inmunogénico que c225 por estar reformado, recibió la aprobación del Centro Estatal para el Control de Medicamentos (CECMED) para iniciar la fase I de ensayos clínicos en pacientes con tumores de origen epitelial en Cuba, y del HPB para iniciar ensayos clínicos en Canadá. Estos ensayos en Canadá comenzaron en noviembre de 1998. De lo anterior se deriva la novedad científica y tecnológica de este resultado. Se desarrolló un nuevo método de reducción de la inmunogenicidad de anticuerpos recombinantes mucho más simple desde el punto de vista de la manipulación genética y que consume mucho menos tiempo de trabajo, comparado con el método clásico de humanización reportado en la literatura. Este método se sustenta en la naturaleza de antígenos T-dependientes de las inmunoglobulinas, y en la gran homología existente entre las regiones variables de las inmunoglobulinas humanas y murinas. El carácter innovativo del método consiste en que sólo la humanización de las regiones anfipáticas (posibles epitopos T), que constituyen entre 10 y 27% de las regiones variables, permite una reducción considerable de la inmunogenicidad, lo que se logra con unas pocas mutaciones puntuales. Es decir, con este método, una vez construido el anticuerpo quimérico, sólo hay que introducir algunas mutaciones puntuales que sustituyan residuos de aminoácidos humanos en las regiones marcos murinas, de forma tal que el quimérico modificado reconozca al antígeno con afinidad similar a la del anticuerpo quimérico y sea menos inmunogénico que éste, sin necesidad de hacer un anticuerpo humanizado. Reconocimientos Los autores dejan constancia de su reconocimiento a Alejo Morales Morales, Gumersinda Bombino, Irene Bousulei, Mayra Ramos y Normando Iznaga por su importante colaboración. Bibliografía Mateo C, Lombardero J, Armour K, Moreno E, Pérez R.
Reshaping of a human monoclonal antibody to epidermal growth factor
receptor to recover binding affinity. Biotecnología
Aplicada1997;14:49. Copyright 1999 Elfos Scientiae The following images related to this document are available:Photo images[ba99031a.jpg] [ba99031b.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}