 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
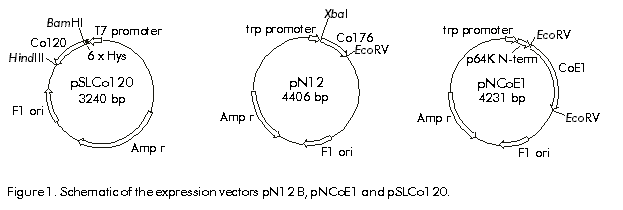
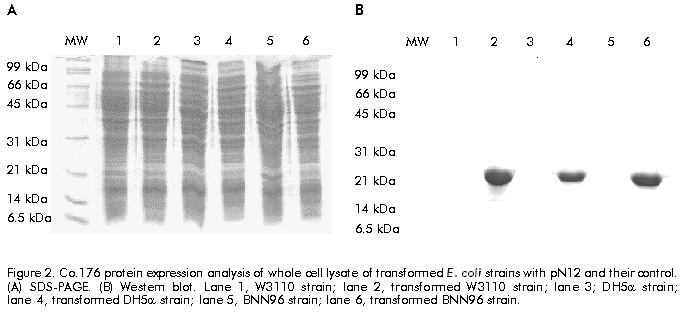
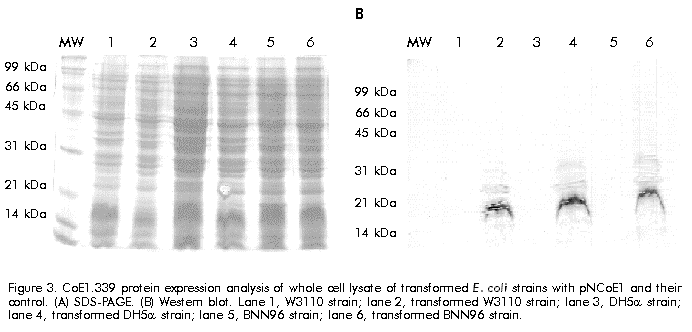
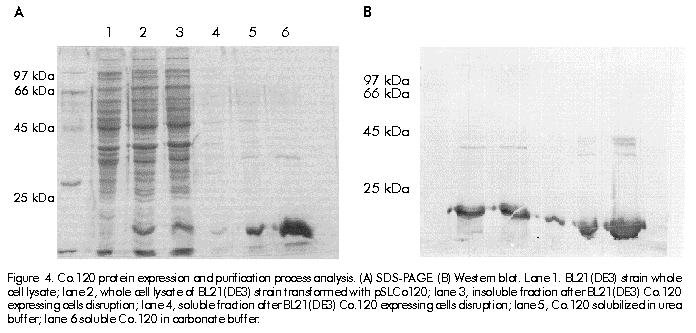
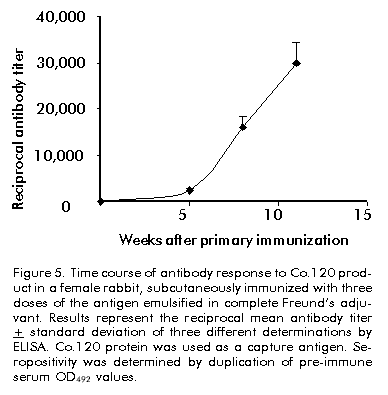
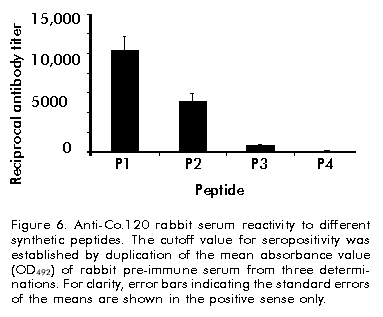
Biotecnología Aplicada 1999;16:226-231 Variable Level Expression of Hepatitis C Virus Core Protein in a Prokaryotic System. Analysis of the Humoral Response against It in Rabbits Santiago Dueñas-Carrera, Juan Morales, Nelson Acosta-Rivero, Lázaro J Lorenzo, Ciro García, Thelvia Ramos, Ivis Guerra, Maxlenin Peña Centro de Ingeniería Genética y Biotecnología. AP 6162, CP 10600, Habana, Cuba. Fax: (53-7) 21 4764; E-mail: juan.morales@cigb.edu.cu Received in Mach, 1999. Accepted for publication in June, 1999. Code Number:BA99036 ABSTRACT Experimental evidence suggests that the hepatitis C virus (HCV) core protein has several biological properties and is implicated as a viral factor in HCV-mediated pathogenesis. In this study, CoE1.339, Co.176 and Co.120 variants of the HCV capsid protein were produced at variable levels in Escherichia coli from pNCoE1, pN12 and pSLCo120 plasmids, respectively. The chimera CoE1.339 spans the first 339 amino acids of the viral polyprotein, fused to the 45 amino acids stabilizer peptide from P64K protein of Neisseria meningitidis. Co.176 is a non-fused variant encompassing amino acids 1–176 of the capsid protein sequence. Both CoE1.339 and Co.176 were expressed under the transcriptional control of the tryptophan promoter and the protein was only detected by Western blot. However, Co.120, a variant truncated at the C-terminus which contains amino acids 1–120 of the HCV core protein fused to a leader tag of six histidines, was successfully produced under the control of the T7 promoter in BL21 (DE3) cells. Like Co.176 and CoE1.339, Co.120 was efficiently recognized by human anti-HCV positive serum. Purified Co.120 antigen showed to be immunogenic in rabbits. Synthetic peptides covering amino acids 1–40 of the core protein showed the greatest reactivity to the rabbit anti-Co.120 serum. Keywords: E. coli, expression, HCV, hepatitis C virus, immunogenicity, nucleocapsid core protein, purification RESUMEN Nivel de expresión variable de la proteína de la cápsida del virus de la hepatitis C en un sistema procariótico. Análisis de la respuesta inmune humoral en conejos. La proteína de la cápsida del virus de la hepatitis C (VHC) posee numerosas propiedades biológicas y está implicada en los mecanismos de patogénesis viral. En este trabajo, las variantes CoE1.339, Co.176 y Co.120 de la proteína de la cápsida fueron producidas a niveles variables en Escherichia coli a partir de los plásmidos pNCoE1, pN12 y pSLCo120, respectivamente. La proteína quimérica CoE1.339 abarca los primeros 339 aminoácidos de la poliproteína viral, fusionados al péptido estabilizador de 45 aminoácidos de la proteína P64K de Neisseria meningitidis. Co.176 es una variante no fusionada correspondiente a los aminoácidos 1–176 de la proteína de la cápsida. Las variantes CoE1.339 y Co.176 fueron producidos a partir de sus respectivos genes bajo el control transcripcional del promotor triptófano, a niveles sólo detectables por Western blot. Sin embargo, la variante Co.120, compuesta por los primeros 120 aminoácidos de la proteína de la cápsida fusionados a un péptido líder de seis histidinas, fue producida con éxito en la cepa BL21(DE3). Al igual que Co.176 y CoE1.339, Co.120 fue reconocida por un suero positivo humano anti-VHC. La proteína Co.120 purificada resultó ser inmunogénica en conejo. Péptidos sintéticos que abarcan los primeros 40 aminoácidos de la proteína de la cápsida, mostraron la mayor reactividad contra el suero de conejo anti-Co.120. Palabras claves: E. coli, expresión, inmunogenicidad, nucleocápsida, purificación, VHC, virus de la hepatitis C Introduction Hepatitis C virus (HCV) is the major causative agent of parenterally transmitted non-A non-B hepatitis. HCV infection leads to a high rate of chronic hepatitis with an attendant risk of cirrhosis and eventual development of hepatocellular carcinoma [1]. Failure in the development of a protective immune response against this viral infection may be due to the low replication rate of HCV, which results in a poor presence of virions in the blood stream and, therefore, in an inadequate antigenic stimulation [2]. This virus possesses a positive-stranded genome encoding a single polyprotein precursor of approximately 3000 amino acids (aa) which is cleaved by both host and viral proteases to generate three putative structural proteins (core, E1 and E2), and at least six nonstructural proteins (NS2, NS3, NS4A, NS4B, NS5A, NS5B) [3]. The core protein is generated from the N-terminal 191 aa of HCV polyprotein. It is cleaved from the precursor polyprotein by the host signal peptidase. This core protein undergoes post-translational modifications before it is incorporated into mature virions [4]. The HCV core protein possesses numerous intriguing properties including in vitro RNA and ribosome binding activities [4], nucleocytoplasmic localization [5], a regulatory role on cellular and unrelated viral promoters [6, 7], an inhibitory role in apoptosis under certain conditions, and interaction with cellular proteins [8]. Of the various regions of HCV, antibodies against the core protein are detected most frequently in patients with chronic hepatitis C, and several B cell epitopes within the HCV core protein were determined [9]. The association of T-helper (Th) cell response to the HCV core protein with a benign clinical course has also been suggested [10]. The capsid protein has been widely expressed in insect cells [11], mammalian cells [12], yeast [13] and Escherichia coli [14]. These recombinant core proteins have been found useful for blood donor screening and for the diagnosis of hepatitis C [12]. In this paper, the expression of capsid protein variants containing different portions of the C-terminal hydrophobic region of the molecule, is described. A novel approach to obtain the full-length capsid sequence as a chimera core-E1 is being undertaken. Different E. coli strains and transcriptional control systems are being analyzed to achieve high levels of the HCV core protein. In this article, the use of the 45 aa N-terminal region of Neisseria meningitidis P64K protein as a stabilizer peptide for the production of the HCV proteins, is reported for the firt time. Finally, new data are shown about a preliminary evaluation of the humoral immune response induced in rabbits by an E. coli-derived core protein. Materials and Methods Peptides The following synthetic peptides covering different regions of the HCV capsid protein were used in this work: P1, PKPQRKTKRNTNRRPQDVKFPGGG QIVGGVY (aa 5–35); P2, DVKFPGGGQIVGGVYL LPRR (aa 21–40); P3, PGGGQIVGGVYLLPRR GPRLGV (aa 25–46); P4, GWLLSPRGSRPSWGPTD PRRRS (aa 95–116). Peptides were synthesized according to the solid-phase method [15] on 4-methylbenzhydrylamine (MBHA) resin (1 mmol/g, Fluka, Switzerland), using the tert-butyloxycarbonyl/Benzyl strategy. Peptide-resin bond was cleaved with hydrogen fluoride (HF) using the "Low-High" procedure in the presence of appropriate scavengers and washing three times with ether [16]. Peptides were extracted with 30% acetic acid. The peptides were purified by reverse phase high performance liquid chromatography (RP-HPLC) (Vydac C18, 10 x 250 mm). Peptides were solubilized in dimethyl sulfoxide/H2O (1:24, v/v) at a concentration of 2 mg/mL. Human serum Samples of human serum were obtained from Centro de Inmunoensayo (Havana, Cuba) where they were previously screened for anti-HCV antibodies by UMELISA HCV (Centro de Inmunoensayo) and Ortho HCV 2.0 ELISA (Ortho Diagnostic Systems, USA). Plasmids and construction of expression vectors Plasmids pXEI.Co176, pBCoE1 and pBEI.Co120 are derived from pBluescript and comprise 528, 1017 and 360 nucleotides (nt) at the 5' end of the structural region coding sequence of the Cuban HCV strain genome, respectively [17]. Plasmids pM100 and pM104 are vectors for the expression of foreign sequences under the control of the tryptophan promoter (ptrp) and the T4 terminator. In the pM100 plasmid, the sequence coding for the 45-aa N-terminal region of the N. meningitidis P64K protein is located upstream of the multiple cloning site for fusion type production of heterologous proteins. For direct production of the capsid protein, the pN12 plasmid was generated. The sequence coding for a 176-aa truncated capsid protein was obtained by double digestion XbaI/EcoRV of pXEI.Co176. This HCV DNA fragment was inserted into compatible sites of the expression vector pM104. The resulting plasmid was digested with NcoI and XbaI, treated with S1 nuclease and religated, originating the expression plasmid pN12. For fusion type expression, the pNCoE1 vector was first generated. A 1017-nt fragment coding for the capsid protein and the first 148 aa of the E1 protein, was obtained when the pBCoE1 plasmid was digested with EcoRV. This segment was inserted downstream from the sequence coding for the 45-aa N-terminal region of the N. meningitidis P64K protein in the pM100 plasmid, also treated with EcoRV, to produce pNCoE1. The pSLCo120 plasmid was obtained to express the hydrophilic and immunologically relevant domain of the core protein fused to a leader tag o box histidines under the control of the T7 promoter. The 360-nt DNA fragment, obtained from BamHI/HindIII digestion of pBEI.Co120, was ligated to the compatible sites of the plasmid pRSET-A (Invitrogen, USA) to create pSLCo120 (Figure 1). To exclude cloning artifacts, plasmids pN12, pNCoE1 and pSLCo120 were confirmed by sequence analysis. Figure 1. Schematic of the expression vectors pN12 B, pNCoE1 and pSLCo120. Bacterial strains XL-1 Blue, an E. coli K12 strain, was used as host for generating plasmid. The following E. coli strains were used for the expression of the capsid variants: W3110 (F- mcr A mcr B IN(rrnD-rrnE) hsdR- hsdM+), BNN96 (F- D(lac) U169 lon- 100 araD139 rps L(Strr) supF mcr A trpC22), DH5a (F- enda1 hsd17 (rk-mk+) sup E44 thi-1 recA1 gyr A96 relA1 D(lacZYA-argF)U169 deoR (f80dlacD (lacZ) M15), BL21(DE3) (F- ompT [lon]hsdSB (rB-mB); E. coli strain B with DE3, a l prophage containing the T7 RNA polymerase. Cell growth and expression conditions For expression studies of Co.176 and CoE1.339 proteins, E. coli strains DH5a, W3110 and BNN96 were used. Competent cells were transformed with pN12 and pNCoE1 expression plasmids. Recombinant cells were grown in Luria Bertani (LB) broth, LB with 5% of glucose (LBG) and minimum medium for bacteria (MM). When needed, ampicillin was added to a final concentration of 50 mg/mL. For expression experiments, an aliquot from 5 mL overnight cultures was diluted in 25 mL of fresh medium to an optical density at 640 nm (OD640) of 0.1 (Photoelectric colorimeter, Model AE330, EOW, LTD). Protein expression was repressed by the addition of tryptophan (Sigma, USA) at 20 mg/mL. Growth was continued with vigorous shaking until early exponential phase (0.6–0.8 OD640). At that point, induction was performed with b-indoleacrylic acid (Sigma, USA) at 50 mg/mL. Cells were harvested at late stationary phase (1.5 OD640) by centrifugation at 3000 xg for 20 min at 4 ºC. For the expression of Co.120 protein, pSLCo120 was introduced into E. coli BL21 (DE3) competent cells. Recombinant BL21 (DE3) cells were grown overnight at 37 ºC in 25 mL of LB broth containing 50 mg/mL ampicillin. A portion of the culture was diluted in 300 mL of fresh medium to 0.1 OD640 and cultured at 37 ºC with vigorous shaking. At the early exponential phase of growth (0.5 OD640), isopropyl-b-d-thiogalactopyranoside (IPTG) (Sigma, USA) was added to a final concentration of 2 mM and the cells were incubated at 37 ºC to produce recombinant protein. Five hours later, the cells were harvested by centrifugation at 3000 xg for 20 min at 4 ºC. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and Western blot Samples were separated in 15% SDS-PAGE [18] and either stained with Coomassie brilliant blue R250 (Sigma, USA) or electrotransferred [19] to a Hybond-C membrane (Amersham, UK). The amount of expressed proteins was estimated from a stained gel using a densitometer CS-9000 (Shimadzu, Japan). After electrotranfer, the membrane was treated for 1 h at room temperature with phosphate-buffered saline (PBS) (0.1 M NaCl, 2 mM KCl, 10 mM Na2HPO4, 1 mM KH2PO4, pH 7.2) containing 5% skim milk, then incubated with a 1:20 dilution of a patient’s serum for 1 h at 37 ºC. After washing with PBS, the membrane was incubated with a 1:1000 dilution of horseradish peroxidase-conjugated goat anti-human IgG (Amersham, UK) for 1 h at 37 ºC. The enzymatic reaction was developed using 0.2 mg/mL 9-amino-4-ethyl carbazole (Sigma, USA) and 0.05% (v/v) H2O2. Enzyme-linked immunosorbent assay (ELISA) The 96-well microtiter plates (Costar, Cambridge, MA) were coated with 100 µL of Co.120 (10 µg/mL) or synthetic peptides (1 µg/mL) diluted in coating buffer (50 mM carbonate buffer, pH 9.6) at 4 ºC overnight. After three washes with PBS containing 0.05% Tween 20 (PBST), the wells were blocked with 200 µL of PBST containing 1% skim milk at room temperature for 1 h. Thereafter, 100 µL of the rabbit serum were added at different dilutions in PBST (1:10 to 1:60,000), and the plates were incubated at 37 ºC for 1 h. The plates were washed five times with PBST and 100 µL of 1:3000 diluted horseradish peroxidase-conjugated goat anti-rabbit IgGs (Amersham, UK) were added. The plates were incubated at 37 ºC for 1 h washed again five times with PBST and incubated with 100 µL of a solution containing 0.05% (w/v) o-phenylenediamine (Sigma, USA) and 0.05% (v/v) H2O2 at room temperature for 10 min. Subsequently, 50 µL of 3 N H2SO4 were added to stop the reactions and the absorbance at 492 nm was determined in a SensIdent Scan (Merck, Germany). The cutoff value was established by duplication of the mean absorbance value of rabbit pre-immune serum from three determinations. Purification of Co.120 protein The 300-mL culture of Co.120-producing BL21 (DE3) cells was harvested 6 h after induction with 2 mM IPTG by centrifugation at 3000 xg for 20 min at 4 ºC. Cell disruption was performed by using the French press (Braund, 1500 kg/cm2) at 1 g (wet weight)/mL in 10 mM Tris-6 mM EDTA. After clarification by centrifugation at 10,000 xg for 20 min at 4 ºC, the insoluble part of the cell lysate was used for purification of the capsid protein by wash pellet procedure. Briefly, the insoluble fraction was washed twice with 0.5 M urea, 1% Triton X-100, 10 mM EDTA, 300 mM NaCl, 10 mM 2-mercaptoethanol, 50 mM Tris-HCl, pH 8.0. Co.120 protein was subsequently solubilized by increasing the concentration of the urea up to 2 M. After centrifugation at 10,000 xg for 20 min at 4 ºC, the supernatant was loaded onto a Sephadex G-25 column equilibrated with carbonate buffer pH 10.6. Desalting by gel filtration rendered a soluble Co.120 protein with 95% of purity. Rabbit immunization After purification of Co.120, a female rabbit (4 months old) was subcutaneously immunized with 200 mg of Co.120 antigen emulsified in complete Freund’s adjuvant to raise anti-core protein antibodies. The rabbit was boosted at 3, 6 and 9 weeks after primary immunization. Serum samples were taken at 0, 2, 5, 8, and 11 weeks after the first immunization. Results Expression of Co.176 and CoE1.339 proteins pNCoE1 and pN12 were first assayed for expression of CoE1.339 and Co.176 proteins, respectively. BNN96, DH5a and W3110 E. coli strains were used as hosts. The CoE1.339 protein was predicted to be a core-E1 chimera comprising the first 339 aa of HCV polyprotein precursor fused to the leader 45-aa N-terminal region of N. meningitidis P64K protein. The Co.176 protein was designed to be a 176-aa non-fused capsid variant. When bacterial extracts were analyzed by SDS-PAGE, it was not possible to detect by Coomassie brilliant blue staining the expression of Co.176 in recombinant cells (Figure 2A). Nevertheless, in an immunoidentification assay using an anti-HCV positive human serum, Co.176 was detected at the predicted molecular weight of 21 kDa (Figure 2B). Figure 2. Co.176 protein expression analysis of whole cell lysate of transformed E. coli strains with pN12 and their control. (A) SDS-PAGE. (B) Western blot. Lane 1, W3110 strain; lane 2, transformed W3110 strain; lane 3; DH5a strain; lane 4, transformed DH5a strain; lane 5, BNN96 strain; lane 6, transformed BNN96 strain. Likewise, the CoE1.339 protein was only detected by Western blot (Figure 3). A specific recognition of chimeric protein bands by the same anti-HCV positive human serum was observed, with molecular weights ranging from 31 kDa to 22 kDa. Figure 3. CoE1.339 protein expression analysis of whole cell lysate of transformed E. coli strains with pNCoE1 and their control. (A) SDS-PAGE. (B) Western blot. Lane 1, W3110 strain; lane 2, transformed W3110 strain; lane 3, DH5a strain; lane 4, transformed DH5a strain; lane 5, BNN96 strain; lane 6, transformed BNN96 strain. To increase the expression level of these core protein variants, the effect of growth medium composition was studied. Three media were tested: minimal medium of bacteria, in which the minimal requirements for growth were supplied; LB; and LBG. Recombinant CoE1.339 and Co.176 proteins could not be detected by SDS-PAGE for none of the three media tested (data not shown). Expression and purification of Co.120 Expression vector pSLCo120 was introduced into E. coli BL21 (DE3) competent cells for expression of Co.120 protein after induction with 2 mM IPTG. The whole cell lysates of control and transformed BL21 (DE3) were analyzed by SDS-PAGE (Figure 4A). The truncated capsid protein Co.120 showed a slightly retarded migration on SDS-PAGE gels. Although the expected molecular weight of Co.120 was approximately 18 kDa, this protein was detected at 21 kDa. Densitometric analysis revealed that about 15% of cellular proteins was estimated to be Co.120 (Figure 4A, lane 2). This fusion protein was absent in the control lysate. For specific detection of the protein, Western blot analysis with an anti-HCV positive serum was performed (Figure 4B). The results were in accordance with those observed for SDS-PAGE. Co.120 was detected by immunoblotting, where, unlike the negative control, it yielded HCV-specific signal. Figure 4. Co.120 protein expression and purification process analysis. (A) SDS-PAGE. (B) Western blot. Lane 1. BL21(DE3) strain whole cell lysate; lane 2, whole cell lysate of BL21(DE3) strain transformed with pSLCo120; lane 3, insoluble fraction after BL21(DE3) Co.120 expressing cells disruption; lane 4, soluble fraction after BL21(DE3) Co.120 expressing cells disruption; lane 5, Co.120 solubilized in urea buffer; lane 6 soluble Co.120 in carbonate buffer. Since Co.120 protein was the only variant efficiently produced in E. coli, it was selected for further studies. To obtain purified Co.120 protein a simple purification procedure was adopted. The protein was purified from 300-mL cultures. Most of the Co.120 protein (95%) remained insoluble after cell disruption for the conditions used in this study (Figure 4A). After desalting and elimination of the chaotropic agent by gel filtration chromatography of the solubilized insoluble fraction, Co.120 protein was obtained with a 95% homogeneity. Evaluation of the humoral response induced by Co.120 The time course of the antibody response against Co.120 protein is shown in Figure 5. Seroconversion was followed using a 1:10 dilution of serum and the antibody titers were evaluated using serial dilutions of the serum samples in an ELISA assay for Co.120 antigen. The immunized rabbit seroconverted two weeks after the primary immunization. Booster doses provoked an increase in the antibody titer of up to 1:30,000. Figure 5. Time course of antibody response to Co.120 product in a female rabbit, subcutaneously immunized with three doses of the antigen emulsified in complete Freund’s adjuvant. Results represent the reciprocal mean antibody titer + standard deviation of three different determinations by ELISA. Co.120 protein was used as a capture antigen. Seropositivity was determined by duplication of pre-immune serum OD492 values. Eleven weeks after primary immunization, the rabbit serum was tested against synthetic peptides covering different regions in the Co.120 sequence. No reactivity was found against the C-terminal P4 peptide. High level antibody titers were detected to peptides comprising the N-terminal region. The P1 peptide, comprising amino acids 5–35 of the capsid protein, resulted the most immunoreactive peptide against the anti-Co.120 rabbit serum (Figure 6). Figure 6. Anti-Co.120 rabbit serum reactivity to different synthetic peptides. The cutoff value for seropositivity was established by duplication of the mean absorbance value (OD492) of rabbit pre-immune serum from three determinations. For clarity, error bars indicating the standard errors of the means are shown in the positive sense only. Discussion It is still unclear whether HCV infection can generate a protective immune response in humans or chimpanzees. In vitro neutralization of HCV with human plasma from an HCV-positive patient prior to inoculation of seronegative animals has been shown to prevent HCV infection [20]. However, neutralizing antibodies have been detected in patients only early after infection, suggesting that the antibody repertoire does not remain stable because of the presence of circulating quasispecies [21]. In contrast to the highly variable envelope glycoproteins, the virion-associated core protein shows a greater sequence conservation among HCV strains [22]. Humoral and cytotoxic activities against this antigen have been abundantly observed in humans and mice [10, 21, 23, 24]. Clinical observations also suggest that it may be possible to enhance viral clearance during persistent infection if the host cellular immune response to HCV core antigenic determinants could be augmented then [23]. Due to the attractive features of the HCV core protein and the goal of supplying large amounts of inexpensive and pure antigen for further experiments, it was decided to produce the nucleocapsid protein in a prokaryotic system. This system has been used favorably to synthesize different HCV capsid protein variants [14, 25, 26]. In some cases, small stabilizer peptides have been employed to increase protein expression [14, 27]. Specifically, the N-terminal domain of the N. meningitidis P64K protein has demonstrated to be effective in stabilizing the expression of several heterologous antigens [27, 28]. This E. coli fusion expression system was assayed with the aim of producing a chimeric core-E1 protein. Different capsid fusion proteins and a chimeric capsid-NS3 recombinant protein, comprising the total capsid region, have been expressed [14, 26]. However, overexpression of the CoE1.339 capsid variant under control of ptrp from pNCoE1 could not be achieved. ptrp is a very strong and autoregulated promoter that has been successfully used for high level expression of different proteins in E. coli [27]. Bacterial proteolytic processing during expression could be the reason why several bands of CoE1.339 protein were detected by Western blot, and the undetectable levels of this core protein variant on SDS-PAGE gels may be explained by an inefficient protein synthesis. The expression of the entire capsid region in a bacterial system may cause problems of solubilization due to the hydrophobic nature of the non-hybrid recombinant protein. Previous experiments demonstrated that E. coli cells, transformed with a plasmid containing only the full-length capsid gene (aa 1–191) under the control of the tac promoter, died shortly after induction by IPTG [25]. The posibility of overexpression of a 176-aa capsid protein from pN12 plasmid lacking the DNA coding sequence for the highly hydrophobic H1 domain [4], was assessed. The removal of the hydrophobic segment spanning residues 174–191, which acts as a signal sequence for translocation of the E1 glycoprotein, naturally occurs by two cotranslational endoproteolytic cleavages [4]. However, the Co.176 protein was also produced at low levels in different E. coli strains. Probably, the remaining hydrophobic domain spanning amino acids 120–176 [14] is still affecting the production of the core protein in E. coli. Capsid protein variants lacking the whole C-terminal hydrophobic region have been successfully expressed in a prokaryotic system [14, 25]. In this paper, the overexpression of the hydrophilic portion of the capsid protein containing the B cell antigenic determinants of this antigen, was confirmed [9]. Previously, C-terminal truncated capsid protein variants were easily purified and evaluated for recognition by anti-HCV positive human sera, when expressed as tandem units or fusion proteins [14, 25]. The combination of the strong inducible T7 promoter and BL21 (DE3) strain, a lambda lysogen carrying the T7 RNA polymerase gene, was successful for the expression of the aminoterminal portion of the HCV capsid protein (Co.120). Co.120 protein is mostly associated with the insoluble fraction after cell disruption for the conditions specified in this study. This protein, synthesized to high amounts (15% of total cellular proteins), probably accumulates to form cytoplasmic inclusion bodies. Co.120 purification, mainly based upon wash pellet procedure, resulted simple and effective. The purified Co.120 protein was able to induce high level antibody titers in rabbit. Previously, a capsid variant spanning the first 120 aa of the viral polyprotein has disclosed its immunogenic potential in three different mouse strains. Comparable levels of antibodies and Th cell proliferative responses to the HCV core protein were detected in Balb/c, C3H/He and C57BL/6 mouse strains [24]. These results in rabbit are in agreement with the intensity of the antibody response developed against Co.120 protein in mice. The data obtained in this study about the humoral immune response induced by Co.120 in rabbit confirm an earlier work in which peptides corresponding to the amino terminal region of the core protein have shown the greatest immunoreactivity to anti-HCV positive sera [9]. Some of the core synthetic peptides corresponding to sequences predicted to be highly hydrophilic had the predicted high immunoreactivity. However, the slightly hydrophobic P3 peptide resulted immunoreactive to the rabbit anti-HCV positive serum, in contrast with the strongly hydrophilic and non-reactive P4 peptide. Peptides closest to the N-terminus of the core protein had the highest reactivity to the rabbit anti-Co.120 serum. The purified Co.120 protein is now under further experiments intended to investigate the anti-core antibody response in HCV-infected individuals. Reactivity of Co.120 to human anti-HCV positive serum rises the possibility of introducing this protein in HCV diagnostic tools. Recombinant Co.120 antigen is also useful as a laboratory reagent and for generating anti-core specific sera. Due to the low-level antibody titers against the core protein in anti-HCV positive individuals and the inherent risk of manipulating HCV-infected sera, the anti-Co.120 rabbit sera result very advantageous for the analytical detection of this antigen. Acknowledgments We are grateful to Dr. Carlos Duarte and Dr. Alexis Mussachio for critical and constructive reading of the manuscript. We also want to thank Dr. Juan Roca for providing the pRSET-A plasmid. References
Copyright Elfos Scientiae 1999 The following images related to this document are available:Line drawing images[ba99036a.gif] [ba99036d.gif] [ba99036e.gif] [ba99036c.gif] [ba99036b.gif] [ba99036f.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}