 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biotecnología Aplicada 1999;16:232-235 Coamplificación del gen de la eritropoyetina humana en células de ovario de hámster chino dhfr– resistentes a metotrexato Diana García del Barco, Alina Rodríguez, Elsa Rodríguez, Caridad Tamayo, Julio R Fernández, María M Vásquez, María P Rodríguez, Ricardo Lleonart División de Genética de Células de Mamíferos. Centro de Ingeniería Genética y Biotecnología. AP 6162, CP 10600, La Habana, Cuba. Telf: (53-7) 21 8164; Fax: (53-7) 33 6008; E-mail: diana.garcia@cigb.edu.cu Recibido en diciembre de 1998. Aprobado en junio de 1999. Code Number:BA99037 Resumen Los procesos de amplificación génica desempeñan un papel importante en los mecanismos celulares de regulación. Para el estudio de la amplificación y expresión del gen de la eritropoyetina humana (EPO), se seleccionaron células de ovario de hámster chino (CHO) dhfr– cotransfectadas con genes quiméricos para la expresión de la EPO y la dihidrofolato reductasa (DHFR), y que fueron seleccionadas en presencia de concentraciones crecientes de metotrexato. Los resultados evidenciaron una amplificación génica de tres veces, acompañada por un aumento en la expresión de la EPO hasta 33 pg/célula/día. El amplicón, que contiene secuencias genómicas del hospedero, muestra rearreglos originados durante el proceso de integración-amplificación. Palabras claves: amplificación génica, CHO, EPO, eritropoyetina Abstract Erythropoietin gene co-amplification in methotrexate-resistant Chinese hamster ovary dhfr– cells. Gene amplification plays an important role in cellular regulatory mechanisms. To study the amplification and expression of the human erythropoietin (EPO) gene, Chinese hamster ovary (CHO) dhfr– cells co-transfected with chimeric genes for the expression of EPO and dihydrofolate reductase (DHFR) and selected in the presence of increasing concentrations of methotrexate, were used. The results evidenced a three-fold gene amplification, accompanied by an increase in EPO expression of up to 33 pg/cell/day. The amplicon, which contains host genomic sequences, shows rearrangements originated during the integration-amplification process. Keywords: CHO, EPO, erythropoietin, gene amplification Introducción La amplificación de genes de mamíferos desempeña un papel importante como mecanismo de regulación génica. En procesos neoplásicos, la amplificación génica ha sido asociada con la sobreexpresión de diferentes genes [1–6]. La amplificación génica ha sido empleada para la selección de clones altamente productores de proteínas recombinantes en células de mamíferos, como resultado del aumento paulatino de las concentraciones de agentes selectivos [7–13]. En células de ovario de hámster chino (CHO), los genes amplificados como el de la dihidrofolato reductasa (dhfr) se encuentran en una o más localizaciones cromosomales formando parte de amplicones que contienen grandes fragmentos génicos del hospedero [14–16]. Los mecanismos que gobiernan la amplificación génica parecen involucrar procesos de recombinación y replicación [2, 17]. Algunos autores proponen la formación, a partir de amplicones lineales, de círculos replicativos extracromosomales como vía para la amplificación [18]. No obstante, aún se desconoce mucho sobre los procesos de amplificación génica y la estructura de los amplicones. Su estudio es importante para la comprensión del papel que éstos desempeñan en los diferentes procesos celulares. En este estudio se aborda la amplificación del gen de la eritropoyetina humana (EPO) cotransfectado con el ADNc del gen dhfr murino en células CHO dhfr–, seleccionadas mediante el empleo de concentraciones crecientes de metotrexato (MTX). Materiales y Métodos Líneas celulares Para la expresión del ADNc de la EPO [19] en células de mamíferos, se cotransfectaron los plasmidios pKepo y pSV2dhfr en proporción 9:1 en la línea celular CHO dhfr– [20], mediante el procedimiento de precipitación de ADN con fosfato de calcio [16]. El plasmidio pKepo contiene el ADNc de la EPO bajo el control del promotor del virus SV40. El vector pSV2dhfr posee el gen de la dihidrofolato reductasa murina bajo el control de las secuencias reguladoras del SV40 [21]. Las células CHO dhfr– fueron mantenidas en medio completo (mínimo esencial alfa con nucleósidos, suero fetal de ternero 10%, penicillina 100 U/mL, glutamina 2 mM y piruvato 2 mM a 37 oC en atmósfera de CO2 5%. Todos los componentes del medio fueron adquiridos de Gibco BRL, Estados Unidos. En fase exponencial de crecimiento, 7,5 x 105 células fueron tratadas con tripsina y sembradas en placas de 60 mm (Nunc, Dinamarca). Veinticuatro horas más tarde estas células fueron transfectadas y 48 h después, tratadas con tripsina. La décima parte de la células transfectadas se sembró en medio de selección (mínimo esencial alfa sin nucleósidos, suplido con suero fetal de ternero dializado 10%, penicillina 100 U/mL, glutamina 2 mM y piruvato 2 mM). Las células escogidas por su capacidad de crecer en medio de selección, fueron mezcladas y seleccionadas paso a paso con concentraciones crecientes de MTX (20 nM; 80 nM; 400 nM; 1,6 mM y 6,4 mM). Para incrementar las concentraciones de MTX, se verificó si las células adoptaron su morfología habitual y si el tiempo de doblaje fue igual al de las células parentales. Cada paso de la selección originó una mezcla de células con capacidad para crecer frente a dicha concentración de MTX. Este conjunto policlonal de células fue identificado con el nombre de la mezcla inicial de células después de la selección en medio selectivo (DA/24), seguido del valor de la concentración de MTX a la cual fue seleccionado. Con la línea policlonal resistente a MTX 6,4 mM (DA/24 6.4), se practicó el método de dilución limitante con el objetivo de aislar células independientes y establecer una línea de origen clonal (M29). Análisis de la expresión de la EPO Para las cosechas, un millón de células fue sembrado en frascos de 25 cm2 (Nunc, Dinamarca) en 5 mL de medio completo. Los sobrenadantes de las líneas celulares DA/24, DA/24 6.4 y M29, fueron colectados 24 h más tarde y fueron ensayados con un sistema ELISA comercial para determinar EPO en sangre humana (Boehringer Mannheim, Alemania). A la vez, se hicieron determinaciones de la actividad biológica in vivo a través del ensayo en ratones hipóxicos [22], e in vitro mediante el empleo del método descrito por Krystal en 1983 [19, 23]. Para el análisis de los transcritos de la EPO en las diferentes líneas celulares, se realizó un ensayo de Northern blot según se ha descrito previamente [24]. Se empleó como sonda el ADNc de la EPO obtenido mediante digestión EcoRI–HindIII del plasmidio pKepo, marcado radiactivamente con 32P según el procedimiento descrito por Feinberg y Vogetstein [25]. Análisis del ADN genómico La extracción del ADN genómico se realizó mediante el procedimiento descrito por Ross y Nelson [26]. Para la determinación del número de copias del gen de la EPO por genoma celular, se realizó un Southern blot en el cual se aplicaron muestras de plasmidio pKepo equivalentes a 1, 50 y 100 copias del gen de la EPO por genoma celular. Se normalizaron las señales mediante rehibridación con una sonda de b-actina de ratón. En las digestiones con enzimas de restricción (New England Biolabs, Estados Unidos), se emplearon 20 µg de ADN y se siguieron las recomendaciones del fabricante. El análisis por Southern blot se realizó según se ha descrito previamente [19]. En las hibridaciones se usaron diferentes sondas. Para la detección del gen de la EPO se usó su ADNc, mientras que para la detección del gen de resistencia a ampicillina (amp) se utilizó un fragmento de 364 pb de este gen, obtenido mediante digestión ScaI–BglI del plasmidio pUC19. En todos los casos el marcaje radiactivo se realizó según lo descrito anteriormente para el Northern blot. Los lavados finales se realizaron con fosfato de sodio 40 mM, SDS 1%, pH 7,2, durante 30 min a 65 oC. Resultados y Discusión Expresión del gen de la EPO en células CHO dhfr– Después de la cotransfección de los plasmidios pKepo y pSV2dhfr en proporción 9:1 en la línea celular CHO dhfr–, se obtuvieron 46 clones, lo que representa una frecuencia de transfección de 6,4 x 10-5. Este valor de frecuencia es similar al obtenido por otros autores [7, 8, 13]. A medida que se seleccionaban células resistentes a concentraciones crecientes de MTX, la producción de EPO aumentaba hasta que alcanzó el valor de 33 pg/célula/día (Tabla). En el clon M29 la expresión resultó ser de 25 ± 5 pg/célula/día. El hecho de que el clon M29 tuvo niveles de EPO menores que la línea DA/24 6.4, responde a que el primero es una población clonal obtenida de la mezcla heterogénea que compone la línea DA/24 6.4, donde hay poblaciones celulares con diferentes niveles de expresión. La EPO secretada al medio de cultivo poseía una actividad biológica de 100 000 ± 7 000 U/mg (N = 3) in vitro y mayor de 120 000 U/mg in vivo. Tabla. Amplificación génica y expresión de la EPO.
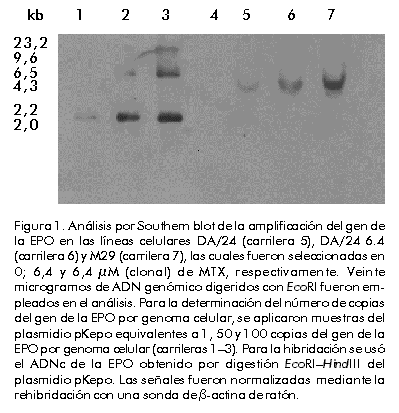
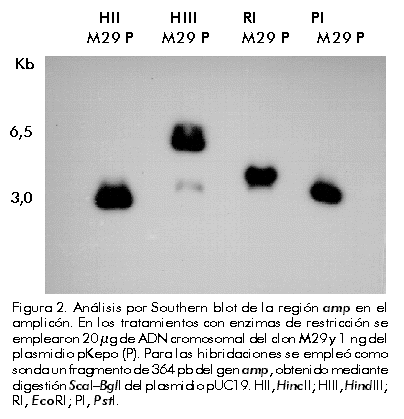
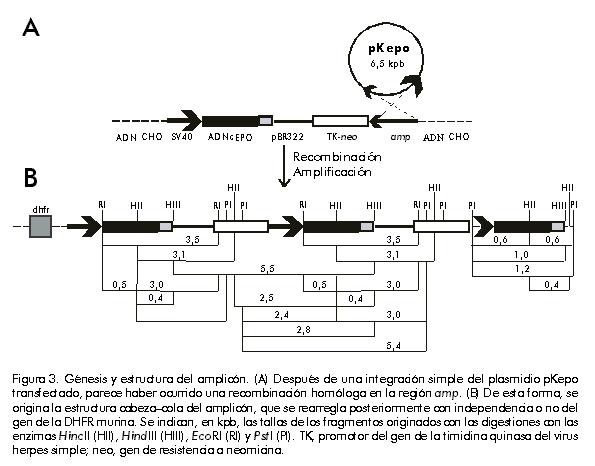
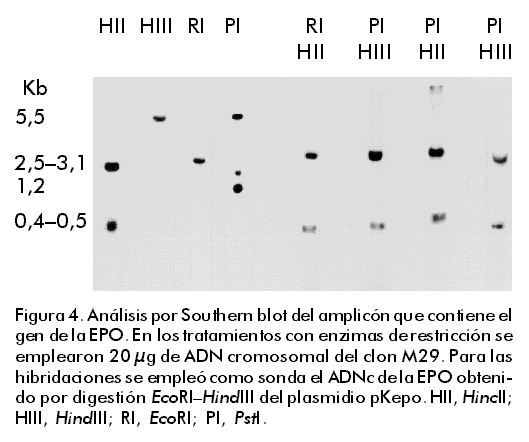
Integración y amplificación génica Para el análisis de la integración y amplificación génicas, se escogieron las líneas celulares DA/24, DA/24 6.4 y M29, seleccionadas en 0, 6,4 y 6,4 µM (clonal) de MTX, respectivamente. Estas líneas contenían 20, 40 y 60 copias del gen quimérico de la EPO por genoma celular, lo que evidenció una amplificación génica de tres veces (Figura 1). Esta amplificación génica se reflejó también en un aumento de la síntesis de EPO por estas líneas celulares (Tabla). Aunque la amplificación obtenida fue relativamente baja, permitió la selección de clones estables con alta producción de EPO. El clon M29 fue mantenido durante más de 90 generaciones en ausencia de MTX con los mismos niveles de EPO (en condiciones de producción en frascos de rotación, diferentes a las ensayadas en el laboratorio, los niveles mantienen en 11 ± 2 pg/célula/día; N = 10). La continuación del proceso de selección en MTX pudiera permitir la identificación de clones con una mayor amplificación, pero se pudiera correr el riesgo de que éstos fueran inestables una vez retirado el agente selectivo del medio de cultivo. Amplificaciones similares han sido reportadas por otros autores [7, 8, 12, 13, 27]. Figura 1. Análisis por Southern blot de la amplificación del gen de la EPO en las líneas celulares DA/24 (carrilera 5), DA/24 6.4 (carrilera 6) y M29 (carrilera 7), las cuales fueron seleccionadas en 0; 6,4 y 6,4 µM (clonal) de MTX, respectivamente. Veinte microgramos de ADN genómico digeridos con EcoRI fueron empleados en el análisis. Para la determinación del número de copias del gen de la EPO por genoma celular, se aplicaron muestras del plasmidio pKepo equivalentes a 1, 50 y 100 copias del gen de la EPO por genoma celular (carrileras 1–3). Para la hibridación se usó el ADNc de la EPO obtenido por digestión EcoRI–HindIII del plasmidio pKepo. Las señales fueron normalizadas mediante la rehibridación con una sonda de b-actina de ratón. El análisis por Southern blot del ADN de la línea celular M29, indicó que el fragmento integrado en el genoma del hospedero había perdido la región correspondiente al gen de resistencia a la ampicillina (Figura 2). Este resultado sugiere que la formación de concatámeros cabeza–cola en el fragmento integrado ocurrió a través de un proceso de recombinación homóloga en la región que incluye al gen amp (Figura 3). Otros autores también han reportado la pérdida de secuencias de origen procariota durante el proceso de integración–amplificación [7, 16]. Figura 2. Análisis por Southern blot de la región amp en el amplicón. En los tratamientos con enzimas de restricción se emplearon 20 µg de ADN cromosomal del clon M29 y 1 ng del plasmidio pKepo (P). Para las hibridaciones se empleó como sonda un fragmento de 364 pb del gen amp, obtenido mediante digestión ScaI–BglI del plasmidio pUC19. HII, HincII; HIII, HindIII; RI, EcoRI; PI, PstI. Figura 3. Génesis y estructura del amplicón. (A) Después de una integración simple del plasmidio pKepo transfectado, parece haber ocurrido una recombinación homóloga en la región amp. (B) De esta forma, se origina la estructura cabeza–cola del amplicón, que se rearregla posteriormente con independencia o no del gen de la DHFR murina. Se indican, en kpb, las tallas de los fragmentos originados con las digestiones con las enzimas HincII (HII), HindIII (HIII), EcoRI (RI) y PstI (PI). TK, promotor del gen de la timidina quinasa del virus herpes simple; neo, gen de resistencia a neomicina. Según se observa en la Figura 4, parece que hubo un solo sitio de integración del gen quimérico de la EPO en el genoma celular. Si hubo más de un sitio de integración, sólo uno de ellos fue coamplificado durante el proceso de selección. Sin embargo, después del proceso de selección, el amplicón contenía, además, un fragmento que comprende parte del promotor del virus SV40 y el ADNc de la EPO (Figuras 3 y 4). Este rearreglo debió originarse después de la integración durante el proceso de amplificación y puede ser funcional o no a nivel transcripcional. El análisis por Northern blot de los transcritos de la EPO (Figura 5) evidencia la existencia de dos transcritos: uno de aproximadamente 3,0 kb, y otro de menor talla (» 1,0 kb) que podría corresponder al producto de transcripción del fragmento rearreglado. Figura 4. Análisis por Southern blot del amplicón que contiene el gen de la EPO. En los tratamientos con enzimas de restricción se emplearon 20 µg de ADN cromosomal del clon M29. Para las hibridaciones se empleó como sonda el ADNc de la EPO obtenido por digestión EcoRI–HindIII del plasmidio pKepo. HII, HincII; HIII, HindIII; RI, EcoRI; PI, PstI. Figura 5. Análisis por Northern blot de la expresión del gen de la EPO. Se emplearon 10 mg de ARN total del clon M29 y de células CHO dhfr– (control negativo), y se realizó la hibridación con la sonda de EPO descrita en la Figura 3. Para la construcción de estas líneas celulares, el gen quimérico que contiene el ADNc de la DHFR murina fue cotransfectado con el plasmidio pKepo. Por consiguiente, aunque el gen dhfr forme parte del amplicón, se encuentra integrado en un sitio diferente, separado del gen de la EPO por secuencias del hospedero que estarían atrapadas en el amplicón (Figura 3B). Sin embargo, como se ha evidenciado en otros casos [16], puede existir un segundo amplicón que contenga solamente el gen quimérico de la DHFR y que sea el responsable de la existencia de clones con gran resistencia a MTX y con bajos niveles de EPO. Este último mecanismo pudiera explicar también la presencia de 20 copias por genoma del gen de la EPO en la línea celular DA/24, transfectada antes de la selección con MTX (Figura 1 y Tabla). Los rearreglos durante el proceso de integración–amplificación reportados en este trabajo, parecen ser frecuentes en la obtención de líneas celulares de mamíferos con altos niveles de expresión de proteínas recombinantes y en otros procesos celulares [2, 7, 16, 28]. Aunque este trabajo no pretende abordar los mecanismos que gobiernan el proceso de amplificación génica en células de mamíferos, los resultados obtenidos apoyan el modelo de amplificación de círculos extracromosomales formados a partir de amplicones lineales [18]. Este modelo al menos permite explicar por qué la gran mayoría de las copias del gen quimérico de la EPO contenidas en el amplicón carece de la región amp. Además, la ausencia de fragmentos de alto peso molecular (mayores de 6 kbp) después de la digestión con HincII o HindIII (Figura 4), puede explicarse por diferencia en el número de copias existente entre los fragmentos detectados en el Southern blot (amplificados) y los extremos del inserto que se encuentran presente en una o pocas copias. Este resultado también es congruente con el mecanismo de amplificación de círculos extracromosomales. Los resultados de este trabajo ejemplifican cómo se estructuran los amplicones en células CHO dhfr– cotransfectadas con construcciones que contienen el gen dhfr y genes quiméricos para la producción de proteínas recombinantes a niveles elevados después de la selección de clones resistentes a altas concentraciones de MTX. Esta información puede contribuir a la estructuración de los procesos de amplificación para lograr clones celulares estables y capaces de producir proteínas recombinantes a altos niveles después de la eliminación del agente selectivo. Agradecimientos Se agradece la ayuda técnica prestada por Amílcar Arenal y Alina Aguirre, de la División de Genética de Células de Mamíferos en el Centro de Ingeniería Genética y Biotecnología de La Habana, Cuba. Referencias
Copyright Elfos Scientiae 1999 The following images related to this document are available:Line drawing images[ba99037a.gif] [ba99037e.gif] [ba99037c.gif] [ba99037d.gif] [ba99037b.gif] | ||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}