 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
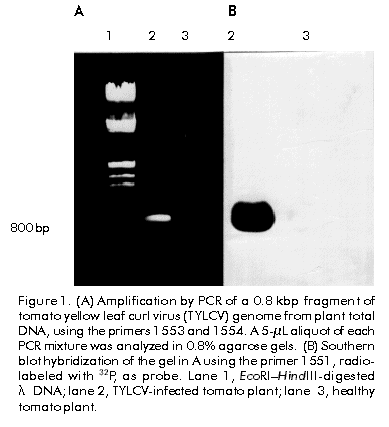
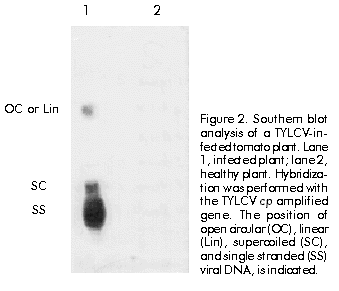
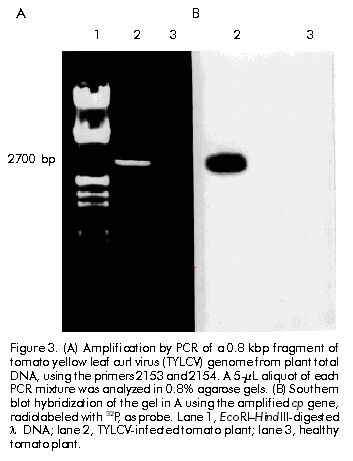
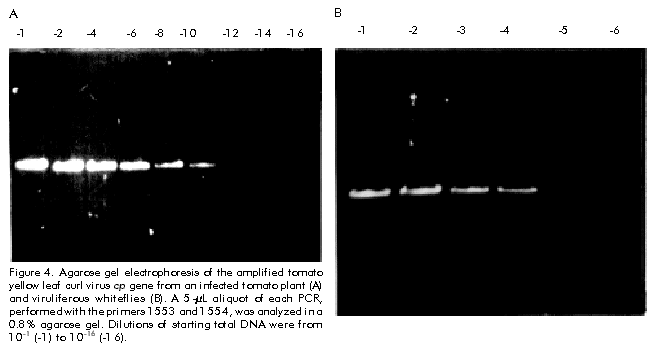
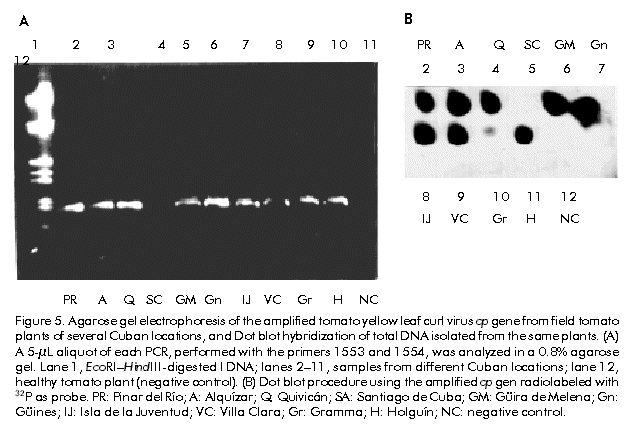
Biotecnología Aplicada 1999;16:237-241 Molecular Techniques for the Detection of Tomato Yellow Leaf Curl Geminivirus in Infected Plants and Viruliferous Whiteflies Lisset Herrera,1 Orlene Guerra,1 Pedro L Ramos,2 Rudy Peral,2 Ana L Echemendía,3 Nadia Ramírez,2 Vivian Doreste,2 Pedro Oramas2 1 Laboratory of Biochemistry. Bioplant Center. University of Ciego de Ávila. Carretera a Morón Km 9,5. CP 69450. Ciego de Ávila, Cuba. 2 Plant Biotechnology Division. Center for Genetic Engineering and Biotechnology. PO Box 6162, Havana, Cuba. Fax: (53-7) 21 8070; E-mail: Pedro.Oramas@cigb.edu.cu 3 Instituto de Sanidad Vegetal. Playa, Ciudad de La Habana, Cuba. Received in October, 1998. Accepted for publication in April, 1999. Code Number:BA99038 Abstract The tomato yellow leaf curl virus (TYLCV) causes major yield losses in tomato production in many tropical and subtropical regions. Therefore, there is an increased need to apply molecular methods for the detection and characterization of the main TYLCV isolates that affect Cuban tomato plantations. A DNA fragment containing the gene encoding the coat protein of a Cuban TYLCV isolate was amplified by polymerase chain reaction (PCR). DNA hybridization with specific radiolabeled probes corroborated the identity of the amplified product, and allowed to analyze the integration into a full-length TYLCV genome. The threshold of TYLCV detection by PCR in plants and viruliferous whiteflies was within the reported range. To detect TYLCV infections, samples collected from tomato plantations of several regions in Cuba were analyzed by PCR. The amplified coat protein gene was also effectively used as a DNA probe in Dot blot assays to detect geminivirus in plants. Keywords: Dot blot, geminiviruses, PCR, TYLCV, virus diagnosis Resumen Técnicas moleculares para la detección del virus del encrespamiento amarillo de la hoja del tomate en plantas infectadas y moscas virulíferas. El virus del encrespamiento amarillo de la hoja del tomate (TYLCV) causa grandes pérdidas en las plantaciones de este cultivo. Los métodos moleculares se hacen cada vez más necesarios para su diagnóstico y caracterización. Mediante el uso de la reacción en cadena de la polimerasa (RCP), se aisló el gen que codifica la proteína de la cápsida del TYLCV. El uso de sondas radiactivas específicas, corroboró la identidad del producto amplificado y permitió analizar la integración en el genoma del TYLCV. El límite de detección del virus mediante la RCP a partir de ADN total de plantas y moscas virulíferas estuvo en el rango reportado. Mediante la RCP se analizaron muestras de tomate de plantaciones de varias localidades de Cuba. El gen de la proteína de la cápsida amplificado también fue utilizado efectivamente como sonda en ensayos de Dot blot para detectar geminivirus en los cultivos. Palabras claves: diagnóstico de virus, Dot blot, geminivirus, RCP, TYLCV Introduction Tomato yellow leaf curl virus (TYLCV) is a whitefly-transmitted geminivirus that causes serious damage to tomato crops in many tropical and subtropical regions from Central America, the Mediterranean, Africa, Asia and Australia [1]. Affected plants produce either no fruit or a few small-sized fruits, depending on the stage of development at which the viral attack occurs, thus causing crop losses of up to 100% [2]. TYLCV, as a member of the geminivirus group, is characterized by a circular single-stranded DNA genome of approximately 2700 nucleotides (nt) encapsidated in twinned quasi-isometric particles (20 x 30 nm) from which the group derives its name [3, 4]. Geminiviruses are subdivided into three groups according to the International Committee on Taxonomy of Viruses, based on insect vectors (leafhopper or whitefly [Bemisia tabaci]), host range (monocotyledoneous or dicotyledoneous plants) and genome structure [5]. Due to the fact that TYLCV is transmitted by whitefly in dicotyledoneous plants, it has been classified as a member of the third subgroup. TYLCV generally presents a monopartite genome, although a bipartite form has been found in an isolate from Thailand [6]. Another study identifies a subset of geminiviruses in the third subgroup according to the geographical distribution as "new world" and "old world" geminiviruses [7]. The genome sequences of TYLCVs isolated from Israel, Sardinia, Murcia, Australia, and some other countries, have been published [8–11]. The TYLCV genome consists of six open reading frames: two on the virion strand including the coat protein gene (cp), which protects viral DNA during transmission by insect vectors and may contribute to vector specificity, and other four on the complementary strand, including the genes necessary for viral replication [4, 12]. Since the late 1980’s, a widespread of geminivirus epidemics have occurred in the Western hemisphere, representing a potential threat for tomato production [13]. Generally, commercial tomato cultivars are susceptible to TYLCV, being inefficient the chemical control of the viral vector because of the development of resistance to insecticides [14]. Breeding for resistance to TYLCV seems to be a promising control approach. The potential of genetic engineering to produce geminivirus-resistant plants is being investigated [4, 14, 16]. Effective management of TYLCV disease epidemics depends on the rapid and accurate identification of the causative agent; therefore, rapid and specific procedures for virus detection are needed. Serological methods have been used with limited success on whitefly-transmitted geminiviruses [17]. Nucleic acid hybridization for the detection of infection has resulted in extremely reliable and sensitive diagnostic procedures. Considerable progress has been made in the development of DNA probes for the detection of viruses, including geminiviruses [18, 19]. Polymerase chain reaction (PCR) has been used with success to detect and characterize viroids [20], cucumoviruses [21] and geminiviruses [19, 22]. Here, the use of PCR for the detection of TYLCV in vegetal material and whiteflies is described, based on the amplification of the cp gene, which was firstly isolated and characterized. It is also evaluated the feasibility to use the amplified product as a specific probe for the detection of geminiviruses. Materials and Methods Vegetal material and virus source Healthy sweetpotato whiteflies (B. tabaci) were kept on tomato plants (Lycopersicon esculentum) infected with TYLCV. These materials were kindly provided by Dr. González (Instituto de Sanidad Vegetal, Havana, Cuba). After a viral acquisition period of one week, the viruliferous whiteflies were maintained on healthy tomato plants var. Campbell 28, which were grown subsequently under greenhouse conditions at 25 ºC. In about 15 days, apical tissues from tomato plants showing typical yellow leaf curl virus symptoms were collected for further analysis. Viral DNA isolation Total DNA was extracted from both TYLCV-infected and healthy plant tissues, according to the procedure described by Dellaporta et al. [23], with the exception that after the first precipitation step with isopropanol, the pellet, diluted in 700 µL of distilled H2O, was treated with 100 µg/mL of RNAse and extracted twice with an equal volume of phenol:chloroform (1:1) [24]. Isolated DNA was resuspended in 400 µL of distilled H2O. Total DNA isolation from whiteflies was performed according to the procedure reported by Mehta et al. [25]. Briefly, five whiteflies, which were previously maintained on TYLCV-infected tomato plants, were collected and placed into a microfuge tube. After grinding in the presence of STE solution (0.1 M NaCl, 10 mM Tris pH 8, 1 mM EDTA pH 8), the resulting suspension was centrifuged at 8000 xg for 10 min. The final pellet was dissolved in 100 µL of distilled H2O. Total DNA preparations from tomato plants and whiteflies were separated on 1.2% agarose gels in order to analyze their quality [24]. PCR conditions Reaction mixtures of 100 µL contained 1 µg of total DNA, 0.2 mM of each dNTP (dATP, dCTP, dGTP and dTTP), 100 pmol of each primer, 50 mM KCl, 10 mM Tris-HCl pH 8.3, 1.5 mM MgCl2, 100 µg/mL gelatin and 3 U of Taq DNA Polymerase (Enzibiot, Cuba). Successive amplification cycles were developed in a thermal cycler (MJ Research, USA). Oligonucleotides were designed according to the nucleotide sequence of the reported TYLCV isolates [8–11]. Amplification of TYLCV cp gene was developed by using the primer 1554 (5'-GTGCATATGCCGAAG CGACCG-3') which corresponds to the 5' end of cp gene, and the primer 1553 (5'-AAGGATCCATTTTAT TAATTTGATATTGAA-3'), which is complementary to the positive strand in the 3' end. The amplification profile consisted of denaturation for 3 min at 92 ºC, annealing for 5 min at 55 ºC and extension for 2 min at 72 ºC; 35 cycles of 1 min at 92 ºC, 2 min at 55 ºC and 2 min at 72 ºC, and a final step of extension for 10 min at 72 ºC. The primers 2153 (5'-CGGGATCCAACTTGGAC ACCTAATGGC-3', 61–80 nt, viral strand) and 2154 (5'-CCTTAAGAAGTAAGACACCGATACACCG-3', 41–60 nt, complementary strand), were used for the amplification of the TYLCV genome. The initial amplification cycle consisted of 2 min at 92 ºC for denaturation, 5 min at 65 ºC for annealing and 5 min at 72 ºC for extension, followed by 34 cycles of 1 min at 92 ºC, 2 min at 65 ºC and 1 min at 72 ºC. The last cycle was completed with 10 min of extension at 72 ºC. Southern blot analysis Total DNA isolated from TYLCV-infected plants and the PCR products were separated by electrophoresis in 1.2% and 0.8% agarose gels, respectively [24]. After an initial depurination step by soaking the gel in 250 mM HCl in order to ensure the efficient transfer of viral supercoiled DNA (total DNA), gels were consecutively soaked in a denaturing solution (1.5 M NaCl, 0.5 M NaOH) for 20 min and in a neutralization solution (1.5 M NaCl, 0.5 M Tris-HCl pH 8.0) for 20 min. DNA was transferred to Hybond N membranes (Amersham, UK) by capillarity and was vacuum-fixed at 80 ºC during 2 h. Both, the amplified cp gene and the oligonucleotide 1551 (5'-ACGCCCGTCTCGAAGGTTCG-3'), labeled with 32P, were used as probes. Hybridizations were performed at 62 ºC for 12 h using stringent conditions [26]. Analysis of field tomato plants Samples of tomato plants (var. Campbell 28) were collected from ten Cuban locations (Pinar del Río, Alquízar, Quivicán, Güira de Melena, Güines, Isla de la Juventud, Villa Clara, Granma, Holguín, and Santiago de Cuba). Three samples of each location were analyzed by PCR using the primers 1553 and 1554, using the amplification conditions described above for the cp gene. The amplification products were visualized by electrophoresis in 0.8% agarose gels. The same samples were also analyzed by Dot blot hybridization. Denatured total DNA preparations were spotted on a Hybond N membrane and placed in SSC 20X (3 M NaCl, 0.3 M sodium citrate pH 7.4 for 10 min. The membrane was dried, and DNA was fixed with UV light in a UV crosslinker (Amersham, UK) and hybridized with the cp amplified gene. Hybridization was developed as it was described above for Southern blot [24]. Oligonucleotide synthesis Oligodeoxinucleotide primers were synthesized by the standard phosphoramidate chemical methodology on a Gene Assembler Plus oligonucleotide synthesizer (Pharmacia Biotech, Sweden). Results and Discussion A DNA fragment of approximately 800 bp amplified by PCR using the primers 1553 and 1554 was observed on 0.8% agarose gels for the sample corresponding to infected plants, while no amplification products were obtained from nucleic acids extracted from a healthy plant (Figure 1A). Amplification yield was estimated to be 0.8 µg per 100 µL from an input of 1 µg of total DNA. Although a similar fragment length has been reported for the cp gene from several TYLCV isolates [8–11, 26], DNA hybridization assays were developed to confirm the identity of the amplified product. Figure 1. (A) Amplification by PCR of a 0.8 kbp fragment of tomato yellow leaf curl virus (TYLCV) genome from plant total DNA, using the primers 1553 and 1554. A 5-µL aliquot of each PCR mixture was analyzed in 0.8% agarose gels. (B) Southern blot hybridization of the gel in A using the primer 1551, radiolabeled with 32P, as probe. Lane 1, EcoRI–HindIII-digested l DNA; lane 2, TYLCV-infected tomato plant; lane 3, healthy tomato plant. The amplified cp gene was hybridized with a probe obtained from the primer 1551 through labeling with 32P, which includes a sequence highly conserved among the reported TYLCV isolates [8–11, 27]. As a result, a strong positive signal over 800 bp was observed on the autoradiogram only for the infected plant (Figure 1B). Primer 1551 corresponds to the first of three conserved regions (1: nt 11–17, 2: 67–76, and 3: 221–246) found in cp gene of TYLCV isolates from Israel, Italy and Thailand [28]. Additionally, the viral DNA conformations of TYLCV could be identified in the TYLCV-infected tomato plant using the amplified 800 bp cp gene as probe. Autoradiographic analyses indicated that the probe was able to detect double and single stranded DNA forms of the viral genome in infected plants (Figure 2). Figure 2. Southern blot analysis of a TYLCV-infected tomato plant. Lane 1, infected plant; lane 2, healthy plant. Hybridization was performed with the TYLCV cp amplified gene. The position of open circular (OC), linear (Lin), supercoiled (SC), and single stranded (SS) viral DNA, is indicated. The integration of the amplified cp gene into TYLCV genome was analyzed by hybridization analysis with a full-length TYLCV genome. Firstly, a PCR assay was carried out in order to amplify the TYLCV genome using the primers 2153 and 2154. The nucleotide sequences of these primers hybridize within the common region of TYLCV isolates. A DNA fragment of 2700 bp was observed only in the sample corresponding to the infected plant, which coincides with the length reported for the TYLCV genome [3, 4]. When this product was hybridized with the amplified cp gene, only in the infected plant a strong hybridization signal was observed (Figure 3). Figure 3. (A) Amplification by PCR of a 0.8 kbp fragment of tomato yellow leaf curl virus (TYLCV) genome from plant total DNA, using the primers 2153 and 2154. A 5-µL aliquot of each PCR mixture was analyzed in 0.8% agarose gels. (B) Southern blot hybridization of the gel in A using the amplified cp gene, radiolabeled with 32P, as probe. Lane 1, EcoRI–HindIII-digested l DNA; lane 2, TYLCV-infected tomato plant; lane 3, healthy tomato plant. Detection of TYLCV in whiteflies and field tomato plants Once the identity of the amplified 800 bp product was confirmed, the same methodology was used to detect TYLCV in other plants and viruliferous whiteflies. The presence of TYLCV in whiteflies previously maintained on TYLCV-infected tomato plants, was detected using the primers 1553 and 1554 in a PCR assay. The amplified DNA was observed on agarose gels up to a 1:10-5 dilution of total DNA from five whiteflies (Figure 4). In the case of the infected plants, the cp gene could be amplified from total DNA diluted up to 10-14. Similar detection limits in PCR have been described by Navot et al. [28] and Mehta et al. [25] for whiteflies and plants using other primer combinations. Figure 4. Agarose gel electrophoresis of the amplified tomato yellow leaf curl virus cp gene from an infected tomato plant (A) and viruliferous whiteflies (B). A 5-µL aliquot of each PCR, performed with the primers 1553 and 1554, was analyzed in a 0.8% agarose gel. Dilutions of starting total DNA were from 10-1 (-1) to 10-16 (-16). Additional assays to detect the presence of TYLCV in tomato fields were carried out using the procedure described here. Samples from ten locations across Cuba were collected, mainly from tomato fields showing typical symptoms of infection by TYLCV [29]. Total DNA was isolated from each sample for further PCR amplifications using the primers 1553 and 1554. As a result, it was possible to detect TYLCV in fields from Pinar del Río, Alquízar, Quivicán, Güira de Melena, Güines, Isla de la Juventud (Western region), Villa Clara (Central region), and Granma and Holguín (Eastern region), while in the sample from the plantation from Santiago de Cuba (Eastern region), no amplification signal was observed, which indicated the absence of TYLCV infection in the specific sample (Figure 5A). Simultaneously, a Dot blot procedure was performed with the same DNA samples from Cuban fields. In this case, the amplified cp gene labeled with 32P was used as probe. The hybridization signals obtained on the autoradiogram showed that the same results are obtained using both methods (Figure 5B). TYLCV symptoms have been observed and studied in Cuba since 1987 [29], and the present results allow to confirm at molecular level that TYLCV is widely distributed in the majority of Cuban tomato fields. In previous related works, it was found that two Cuban TYLCV isolates showed a 97.3% [27] and 94.9% identity [30] to a TYLCV isolate from Israel, respectively, when the nucleotide and amino acid sequences of the cp gene and its expression product were compared. Further epidemiological and molecular studies will be necessary in order to characterize the main Cuban TYLCV strains. Figure 5. Agarose gel electrophoresis of the amplified tomato yellow leaf curl virus cp gene from field tomato plants of several Cuban locations, and Dot blot hybridization of total DNA isolated from the same plants. (A) A 5-µL aliquot of each PCR, performed with the primers 1553 and 1554, was analyzed in a 0.8% agarose gel. Lane 1, EcoRI–HindIII-digested l DNA; lanes 2–11, samples from different Cuban locations; lane 12, healthy tomato plant (negative control). (B) Dot blot procedure using the amplified cp gen radiolabeled with 32P as probe. PR: Pinar del Río; A: Alquízar; Q: Quivicán; SA: Santiago de Cuba; GM: Güira de Melena; Gn: Güines; IJ: Isla de la Juventud; VC: Villa Clara; Gr: Gramma; H: Holguín; NC: negative control. Because TYLCV and other geminiviruses are distributed worldwide in tropical and subtropical regions causing serious damages to tomato plantations, the screening and early diagnosis of viral diseases are very necessary to avoid enormous yield losses. The current approaches to detect viral infections are based on indicator plants or serological assays, but in some cases like whitefly-transmitted geminiviruses, they have not been sufficiently effective. The vast difficulties to obtain sufficient quantities of viral antigens for the production of antiserum, together with the imprecise evaluation of symptoms, makes molecular procedures an important tool for a more accurate detection of plant viruses. With the aim to develop a diagnostic procedure for the detection of TYLCV, the cp gene from a TYLCV Cuban isolate was amplified by PCR using the primers 1553 and 1554. The amplification product was firstly characterized through successive hybridization assays aimed at characteristic regions within the TYLCV genome. These primers could be used for the amplification of the TYLCV cp gene from whiteflies and field tomato plants, providing a simple and efficient method for TYLCV detection. By using PCR-based methods, many difficulties associated with serological methods can be overcome, such as low antibody titer, limited availability of antibodies against geminiviruses, and cross-reactivity of antibodies with heterologous antigens [3, 31]. Another alternative is the use of the amplified 800 bp fragment corresponding to the cp gene as a probe in DNA hybridization assays. This DNA fragment has been already cloned into a suitable Escherichia coli vector by the authors’ group, and it can be used as a specific and economical probe for the detection of geminivirus-infected plants using a hybridization procedure like Squash, Dot, Southern and other blotting techniques. Both PCR amplification and DNA hybridization are feasible to use for geminivirus detection, particularity for large-scale diagnosis. These are simple and rapid methodsthat allow the analysis of numerous samples in a short time. However, each has its peculiarities, for example, the necessity to use radioisotopes like 32P for Dot blot procedure or a thermal cycler for PCR. The selection of one of them will depend upon the particular conditions of the laboratory. The procedures described here should be a useful tool for epidemiological studies on the spreading of viruliferous vectors or viruses over crops. Also, they could be used in breeding programs for disease resistance and in the development of antiviral strategies involving recombinant DNA approaches. Acknowledgments This research was supported by the ICGEB project CRP/CUB 94-05/b2. We thank Dmitri Prieto for helpful comments on the manuscript. References
Copyright Elfos Scientiae 1999 The following images related to this document are available:Line drawing images[ba99038b.gif] [ba99038d.gif] [ba99038c.gif] [ba99038e.gif] [ba99038a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}