 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
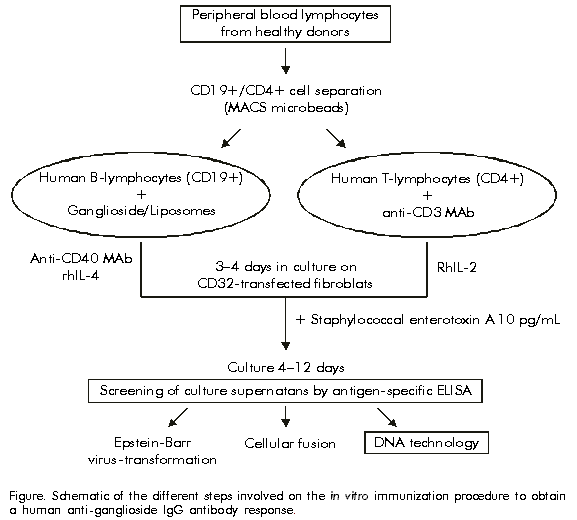
Biotecnología Aplicada 1999;16:246-248 Human Anti-ganglioside IgG Antibody Response Induced by in vitro Immunization with Liposomes Containing GM3(NeuGc) Ganglioside Mauro Alfonso,1 Peter Ifversen,2 Jesper Zeuthen2 1 Center of Molecular Immunology. PO Box 16040, Havana 11600, Cuba. E-mail: mauro@ict.cim.sld.cu 2 Department of Tumor Cell Biology. Danish Cancer Society, DK-2100 Copenhagen, Denmark. Received in November, 1998. Accepted for publication in June, 1999. Code Number:BA99040 ABSTRACT A new in vitro immunization system of human peripheral blood lymphocytes using liposomes containing gangliosides, is described. Purified human B lymphocytes were cultured in the CD40 system in the presence of liposomes containing different concentrations of GM3(NeuGc) ganglioside, ranging from 1 to 1000 ng/mL. After an initial primary in vitro immunization period (3–4 days), antigen-stimulated B cells and autologous activated CD4+ T lymphocytes previously activated with an anti-CD3 monoclonal antibody were cultured in the presence of staphylococcal enterotoxin A. Human antibody production was measured by an anti-ganglioside ELISA after 4 and 8 days of culture. Specific antibody responses against GM3(NeuGc) of IgM and IgG isotypes were induced after 4 days in culture, and a predominant antigen-specific IgG antibody production was observed 8 days after antigenic stimulation. Keywords: ganglioside, human antibody, in vitro immunization RESUMEN Respuesta de anticuerpos humanos anti-gangliósidos inducida por inmunización in vitro con liposomas que contienen al gangliósido NeuGcGM3. Se describe un nuevo sistema de inmunización in vitro de linfocitos humanos de sangre periférica mediante el uso de liposomas que contienen gangliósidos. Linfocitos B humanos purificados fueron cultivados en el sistema CD40 en presencia de liposomas que contenían al gangliósido NeuGcGM3 a diferentes concentraciones (1–1 000 ng/mL), y después de un período inicial de inmunización in vitro primaria (3–4 días), células B estimuladas por el antígeno y linfocitos T CD4+ autólogos activados previamente con un anticuerpo monoclonal anti-CD3, fueron cultivados en presencia de la enterotoxina A estafilocóccica. La producción de anticuerpos humanos en los sobrenadantes de cultivo fue analizada mediante un ELISA anti-gangliósidos después de 4 y 8 días de cultivo. Se obtuvieron respuestas de anticuerpos específicos de tipo IgM e IgG contra NeuGcGM3 después de 4 días de cultivo, mientras que 8 días después de la estimulación antigénica, se encontró una producción de anticuerpos humanos específicos para el antígeno predominantemente de tipo IgG. Palabras claves: anticuerpo humano, gangliósido, inmunización in vitro Introduction In vitro immunization of human B lymphocytes has been developed so as to achieve antigen-specific activation of human B cells and maturation of B cell immune response in vitro. This approach, in combination with some immortalization steps, such as Epstein-Barr virus infection to transform human B cells, and cellular fusion of human B lymphocytes with human-mouse heteromyeloma cell lines, has led to the production of long-term lymphoblastoid cell lines and hybridomas producing human antibodies of the desired specificity [1]. One way to generate human antibodies is to use in vitro immunization systems which allow the induction of primary and even secondary immune responses of human B lymphocytes. Advances on the in vitro immunization methodology developed by different laboratories, which include primary and secondary immunization in the CD40 system, have allowed the generation of human monoclonal IgG antibodies (MAb) with affinities higher than that of antibodies obtained by conventional techniques used for the generation of human MAbs [2]. It has been reported that antigen-specific activation of human B cells can be obtained after stimulating B cells with the antigen in the presence of autologous T helper cells and sub-optimal concentrations of staphylococcal enterotoxin A (SEA) as a T cell mitogen, providing a cognate interaction between B and T helper lymphocytes [3]. Gangliosides are glycosphingolipids containing neuraminic acid, which are present in the plasma membrane of most mammalian cell lines, but are differently expressed, both quantitatively and qualitatively, in certain types of cancer, and can be used as tumor markers and as targets for cancer immunotherapy [4]. Purified gangliosides are poorly immunogenic molecules. To obtain an immune response against this kind of T cell-independent antigens they have to be coupled to carrier proteins or incorporated into liposomes. One particular characteristic of the antibody response to these type of antigens is that the immunoglobulin class produced is mainly IgM, not usually suitable for cancer immunotherapy. In this report, a novel in vitro immunization methodology which allowed to induce human IgG antibody responses against ganglioside antigens, is described. Materials and Methods Antigen, MAbs and growth factors The ganglioside GM3(NeuGc) was isolated from horse erythrocytes [5]. Transfected mouse 3T6 fibroblasts 3T6-CD32 (IGCL 307) and 3T6-CD32/B7.1 (IGCL 306) were obtained from Innogenetics (Belgium). Recombinant human interleukins (rhIL-2 and rhIL-4) were purchased from Boehringer Mannheim (Mannheim, Germany). The hybridoma cell line G28.5 clone (anti-CD40) was obtained from ATCC (HB-9110). The mouse IgG1 anti-human CD3 MAb, UCHT1, was purchased from Pharmingen. Preparation of liposomes Liposomes containing the ganglioside GM3(NeuGc) were prepared as previously reported Mauro Alfonso et al. [6]. Briefly, 50 mg of ganglioside were mixed with equimolar amounts of dipalmitoylphosphatidylcholine and cholesterol, solubilized in chloroform/methanol (2:1 v/v). After exhaustive evaporation of the solvent in a rotary evaporator, 1 mL of phosphate-buffered saline (PBS) pH 7.2 was added and the lipid film was dispersed by vigorous shaking by vortexing. The preparation was then sonicated in a water bath ultrasonicator and used for in vitro immunization experiments. Separation of cells Buffy coats from normal donors were obtained from the University Hospital Blood Bank (Copenhagen, Denmark) and peripheral blood lymphocytes (PBL) were separated by density gradient centrifugation using Lymphoprep (Nycomed A/S, Norway), according to standard protocols [7]. The separation of CD4+ and CD19+ cell subpopulations was performed according to the procedure recommended by Miltenyi Biotec (Bergisch Gladbach, Germany). Briefly, PBLs isolated by density gradient centrifugation were washed with PBS containing 5 mM EDTA and 0.5% bovine serum albumin (BSA), were resuspended in 80 mL of washing buffer per each 107 cells and incubated for 15 min at 4 ºC with 20 mL of MACS colloidal supermagnetic microbeads conjugated with a monoclonal anti-human CD4 antibody per each 107 cells (MACS CD4 Microbeads, Miltenyi Biotec, Germany). The cell suspension was then applied on top of a prefilled and washed CS column, which was placed in the magnetic field of a MACS separator and the cell suspension was allowed to pass through. Magnetically labeled cells (CD4+) were retained in the column, and the effluent was collected as the CD4-fraction. The column was washed, removed from the MACS separator, and the CD4+ cells were eluted by rinsing with five column volumes of washing buffer. The CD4- fraction was then incubated with MACS CD19 Microbeads for 15 min at 4 ºC and applied on top of an RS+ column for positive selection of CD19+ cells. The separation of the CD19+ and CD19- cell subpopulations was performed as described above. Purified T (CD4+) and B (CD19+) cell suspensions were washed with PBS and used for in vitro immunization experiments. Flow cytometry To evaluate the efficacy of cell separation, aliquots of the cell suspension were taken at each step and analyzed by flow cytometry by using specific anti-CD4 and anti-CD19 antibodies conjugated to fluorescein isothiocyanate and R-phycoerythrin, respectively. Only cell subpopulations with more than 95% of either CD4+ or CD19+ cells were used for in vitro immunization experiments. In vitro immunization The primary in vitro immunization procedure using PBLs from healthy donors and the ganglioside GM3(NeuGc) incorporated in to liposomes as immunogenic preparation, has been previously described [8]. Our new approach is based on the use of purified B (CD19+) and autologous T helper lymphocytes (CD4+) for in vitro immunization with gangliosides. Purified B cells (CD19+ fraction) were seeded into flat bottomed 96-well tissue culture plates (2.5 x 104 cells per well), together with irradiated (6500 rad) 3T6-CD32 transfected fibroblasts (1.5 x 104 cells per well) and cultured for 3 days in a medium supplemented with 10% heat-inactivated human serum, 0.5 mg/mL of G28.5 MAb (anti-CD40), 10 U/mL rhIL-4, and liposomes containing GM3 (NeuGc) at a final concentration of 1, 10, 100 and 1000 ng/mL. Simultaneously, purified T cells (CD4+) were cultured on irradiated (6500 rad) 3T6-CD32/B7.1 transfected fibroblasts in the presence of 100 ng/mL of a mouse anti-human CD3 MAb, and stimulated with 50 U/mL of rhIL-2. After 3 days in culture CD4+ cells were washed, irradiated (2000 rad) and then co-cultured with the B cells stimulated with liposomes-GM3(NeuGc) in the presence of 10 pg/mL of SEA (Toxin Technology, USA). Culture supernatants were collected after 4 and 8 days and analyzed for the presence of GM3(NeuGc) antibodies by an enzyme-linked immunosorbent assay (ELISA). The Figure shows a schematic of the different steps involved in the in vitro immunization procedure to obtain an anti-ganglioside IgG antibody response. Figure. Schematic of the different steps involved on the in vitro immunization procedure to obtain a human anti-ganglioside IgG antibody response. ELISA procedures A solid-phase anti-ganglioside ELISA was performed as previously reported [8]. Gangliosides (200 ng/well) in 50 mL of methanol were dried in 96-well Polysorp immunoplates (Nunc, Denmark) for 1 h at 37 ºC, and the plates were blocked with 1% BSA in 0.05 M Tris-HCl buffer pH 7.8 for 30 min at 37 ºC. In vitro immunized cells from culture supernatants were incubated for 2 h at 37 ºC, and after washing with PBS, the secondary antibody consisting of alkaline phosphatase-conjugated goat anti-human IgG or IgM (Jackson Immunoresearch Laboratories Inc., USA) diluted 1:5000 was added to the plates and incubated for 1 h at 37 ºC. The wells were washed again and a substrate solution of 1 mg/mL p-nitrophenylphosphate (Sigma, USA) in diethanolamine buffer pH 9.8 was added. After 30 min, the absorbance of the reaction product was measured using an ELISA reader (Organon Teknica, Netherlands) at 405 nm. Results and Discussion Previous in vitro immunization experiments developed by the authors using GM3 ganglioside incorporated into liposomes [8, 9] and other ganglioside antigens such as GM3(NeuGc) and GM2 [10], only lead to the establishment of lymphoblastoid cell lines producing specific human anti-ganglioside IgM antibodies, even when complete tetanus toxoid or a synthetic T helper cell epitope of tetanus toxin (ptt) are incorporated in the liposomes to provide T cell help for the antibody response. It has been demonstrated that the use of immunogenic preparations containing B and T cell epitopes in SCID mice [11] and for in vitro immunization, followed by a restimulation with the antigen in the presence of autologous T cells and culture on the CD40 system [2], leads to the generation of high-affinity human IgG MAbs [12]. However, different experiments performed by the authors under similar conditions but using gangliosides such as GM3 and GM2, have not produced similar results. One explanation could be that the poor immunogenicity of ganglioside antigens, together with the limitations of the in vitro immunization systems that do not provide an appropriate microenvironment for the occurrence of the events involved in cellular activation, hinders the attainment of IgG antibody responses against this type of glycolipid antigens. As an alternative to generate human IgG anti-ganglioside antibodies, an in vitro immunization methodology was developed, in which human B lymphocytes are stimulated with immunogenic liposomes containing GM3(NeuGc) antigen on the CD40 culture system, followed by the addition of stimulated autologous T cells and SEA. It has been demonstrated that B cell activation on this system depends on antigen doses and on the amount of SEA used, and that the antibody production by antigen-specific B cells can be increased due to the crosslinking of the surface immunoglobulins and to B–T cell interactions [3]. Specific antibody responses against GM3(NeuGc) were induced in our experiments. IgM and IgG antibody responses could be detected after 4 days of culture of B and autologous T lymphocytes, and mainly an antigen-specific IgG antibody production was observed after 8 days of culture (Table). Similar results have been obtained using GD3 ganglioside incorporated into liposomes as the immunogenic preparation (data not shown). Table. Specific IgG antibody production after in vitro immunization with liposomes containing GM3(NeuGc) ganglioside, autologous T helper lymphocytes and SEA.
This alternative method is based on the binding of the superantigen SEA to MHC II molecules on B cells, permitting a polyclonal activation of T cells through the interaction with the T cell receptor. This kind of pseudo antigen-specific signal, which is provided to T cells, causes CD40 upregulation on B cells and CD40 ligation has been shown to lower the threshold for sIg activation. Then, the activated B cells are stimulated to proliferate and to produce immunoglobulins [13]. This characteristic of SEA allows its use as a potent tool for in vitro immunization systems, since it provides a high probability of T–B cell interaction even if T and B cells are not specific for the same antigen. The results of our in vitro immunization experiments showed that the human anti-ganglioside antibody response was dependent on the presence of the ganglioside in the immunogenic preparations and on the doses used for immunization, indicating that the antibody response was driven by the antigen and was not due to the presence of pre-existing antigen-specific clones among the pool of lymphocytes used for in vitro immunization. By using the immunization procedure described here, it was possible to induce an anti-ganglioside IgG antibody response not commonly found for in vitro immunized human B cells and even less using T-independent ganglioside antigens. These human anti-ganglioside MAbs are potentially useful for therapeutic applications in cancer patients. Acknowledgment This work was supported by the European Community Biotechnology Program, contracts BIO2 CT-920269 and BIO4 CT-950252. References
Copyright Elfos Scientiae 1999 The following images related to this document are available:Line drawing images[ba99040a.gif] | ||||||||||||||||||||||
| |||||||||

{kind=link}