 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
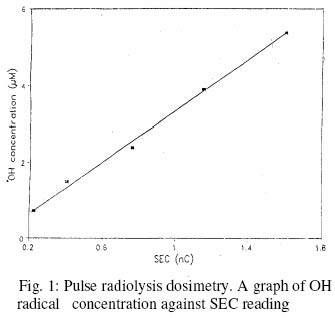
Electron Transfer from Azide Radical to Histidine Generates Histidinyl Radical M. S. Sule*1 and J. Butler2 1. Department of Biochemistry, BayeroUniversity, Kano, Nigeria Received 19 September 2002 Code Number: bk03003 Abstract The formation of histidinyl radical (HR), which is a product of electron transfer reaction between histidine and some free radicals, was studied by pulse radiolysis. The reaction between histidine and azide radicals was found to produce HR, which has a distinct absorption spectrum with peaks at 300, 480 and 520 nm. The formation of HR was found to depend solely on the concentration of azide radicals, with the first order rate of 1.46 x 104 s-1. The implication of the formation of HR in an enzyme, which has histidine as the crucial amino acid to its activity, is discussed. Keywords: Histidine, Histidinyl radical, Azide radical. INTRODUCTIONHistidine is the only amino acid with the imidazole ring in its side chain. It is one of the amino acid classified as having polar and positively charged side chain. Histidine has been reported to be crucial to the activity of many enzymes, such as lactate dehydrogenase (1), fructose diphosphate aldolase (11), chymotrypsin (7,11), ribonuclease (1), subtilisin (8), superoxide dismutase (12), trypsin (4) and yeast alcohol dehydrogenase (6). The imidazole group of the amino acid, which has been shown to act as proton donor or acceptor depending on whether its protonated or not (10), participates in general - acid or general - base catalysis in these enzymes. Histidine side chain has a pKa close to 7 (11), and at near neutral pH, there is reasonable balance between proton - donating (protonated) form and the proton- accepting (deprotonated) form. It is thought that suitably positioned proton - donating and proton-accepting side-chains, such as imidazole, in an enzyme could act in a concerted fashion, so that electrons are both 'pulled' and 'pushed', thereby promoting the conversion of substrate to product (10). Irradiation of water causes the eventual formation of the primary radicals of water radiolysis, i.e. the hydroxyl radical, hydrated electron and the hydrogen atom (14). These radicals have been shown to react rapidly with all the amino acids which occur in proteins (17), as such their reactions with proteins would not lead to the identification of the target amino acid/s. In contrast, inorganic radicals, such as thiocyanate, bromide, chloride and iodide radical anioins, have been shown to react specifically with only certain amino acids at certain pH (4,15). One of the amino acid that the inorganic radicals react rapidly with is histidine. It was reported that thiocyanate, bromide, chloride and iodide radical anions specifically react with histidine to produce histidinyl radicals (HR) e.g. reaction 1 (in which HisH represents the histidine with protonated imidazole ring) (3).
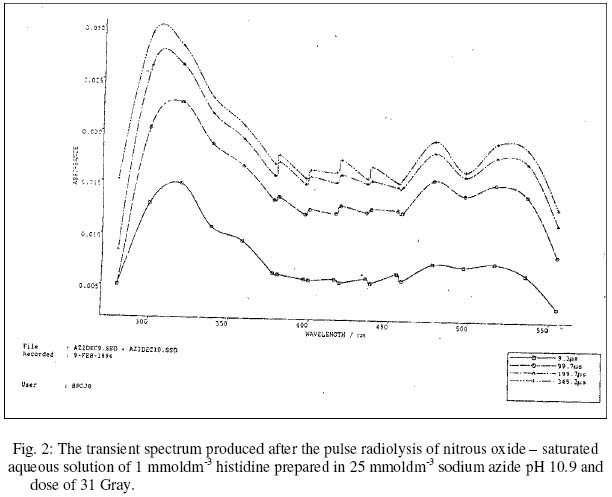
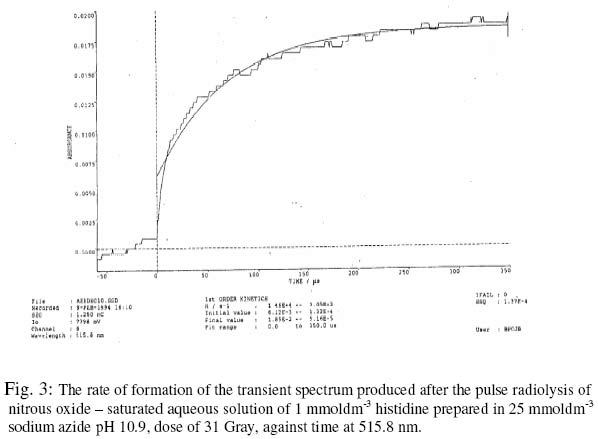
The fact that the products of reaction between free radicals and some amino acids are capable of transferring their electrons to other amino acids within a protein (16) is a major concern of the use of free radical inactivation in the determination of crucial amino acids in the activity of enzymes. However, whereas tryptophan, methionine and tyrosine have been shown to participate in such electron transfer in proteins such as B - lactoglobulin (16), histidine has not been reported to be involved in it. This makes the study of free radical inactivation of enzyme, in which histidine was concluded as the crucial amino acid as in chymotrypsin (7), to be more reliable than those in which tryptophan, methionine or tyrosine was the crucial amino acid. The use of azide radicals in radiation chemistry studies started much later than the other inorganic radicals. Its advantages over the other inorganic radicals have been reported to be as follows (15): Firstly, in contrast to other inorganic radicals, it was found to have a low absorption above 300 nm where the oxidation products of the amino acids absorb. Secondly, it may be capable of reacting in hydrophobic environments of proteins since it is uncharged. It is thought to react with amino acids and other biomolecules by electron transfer, like the other inorganic radicals. Due to the advantages of using azide radicals over other inorganic radicals in free radical - induced enzyme inactivation studies as well as the reliability of the study in order to confirm the importance of histidine in the activity of enzyme; this paper reports on the formation of HR using azide radicals by pulse radiolysis (PR) and the significance of its formation in enzymes with histidine as the crucial amino acid. The dosimetry of the PR equipment is also reported. EXPERIMENTALPulse radiolysis (PR) is a technique in which reactions involving free radicals and other unstable chemical species are studied. The radiation sources suitable for PR must be able to deliver short pulses of high energies sufficient to produce adequate number of chemical species. In this study, Linear (electron) Accelerator (LINAC), which satisfies the condition, was used. The PR facility consists of LINAC, lamp, sample cell, wavelength selector, detector, digitizer and computer (9). The LINAC is shielded in a concrete chamber. The arrangement of the equipment is in such a way that electron beam from the LINAC goes through a secondary emission chamber (SEC) before hitting the PR cell holding the sample. The SEC produces a charge, which is proportional to the pulse dose. The solution to be pulsed, contained in a glassware thoroughly washed with redistilled water, was gently bubbled with the appropriate gas in a special flask easily sealed off by taps. The solution was then taken into the radiation cell using argon gas under pressure. From outside the chamber, a suitable pulse length was selected and a pulse of electrons was automatically delivered to the solution upon pressing the appropriate button. The details of the spectrum of any transient species formed were passed to the computer and could be displayed at specified times after the pulse. Dosimetry of the PR equipment. In studies involving the use of radiation, it is desirable to first of all develop a method of quantifying the doses of radiation delivered by the radiation source. The dosimetry experiment was carried out by pulsing air - saturated aqueous solution (0.01 moldm-3) of potassium thiocyanate at different SEC reading. The irradiation of the above solution caused the hydroxyl (OH) radicals formed to be scavenged by thiocyanate ions to form thiocyanate radical anion (TRA) according to the reactions below: OH. + SCN- → SCN. ------------- 2 SCN. +SCN- ====== (SCN)2.- -----------3 The concentration of TRA could be calculated from its absorption at 500 nm, where its extinction coefficient was found to be 7100 dm3mol-1cm-1 (2). Since all the OH radicals formed was expected to react according to reaction 1, then the molar concentration of OH radicals is the same as the molar concentration TRA. The concentration of OH radicals was calculated at 500 nm using Beer - Lambert's Law at the SEC readings of 0.225, 0.400, 0.765, 1.150 and 1.600 number of charges (nC), using a pathlength of 2.5 cm. Formation of Histidinyl radicalThe reaction of azide radicals with histidine was studied by the PR of nitrous oxide - saturated aqueous solution of 1 mmoldm-3 histidine prepared in 25 mmoldm-3 sodium azide (NaN3), pH 10.9 and a dose of 31 Gray. The nitrous oxide scavenges hydrated electron (eaq- ), resulting in the increase in OH radicals concentration. The 0H radicals react with the azide ion to produce the azide radicals. H2O /\/\/\/\/\/\/ → H. , OH. , eaq- ---------- 4 eaq- + N2O+ H2O → N2 + OH. + OH- --5 OH. + N3- → N3. + OH- ------------6 The determination of the rate constant of the reaction was achieved by following the increase in absorption of the transient species formed with time at 515.8 nm. A fit time range of 0 - 350 us was selected, and the first order kinetic constant was displayed as described previously (9). RESULTS AND DISCUSSIONThe concentration of OH radicals for all the SEC readings used was calculated, and a graph of OH radical concentration against SEC reading is shown in figure 1. The radiation chemical yield (G Value) of OH radicals in air saturated irradiated aqueous solution was found to be 2.8 (15). This means that 100 electron - volt (eV) would give 2.8 OH radicals. Therefore, 1 Gray (i.e. 6.24 x 1018 eVdm-3) would give 1.75 x 1017 molecules dm-3 or 2.9 x10-7 moldm-3 of 0H radicals (after dividing with the Avogadro's number). Using this relation, the dose in Gray could be calculated from the various OH radicals concentration. The reaction between azide radicals and histidine at pH 10.9 was followed by the formation of transient species with the absorption spectrum shown in figure 2. The rate of formation of the transient species at 515.8 nm is shown in figure 3. From the graph, the first-order rate constant was found to be 1.46 x 104 s-1. The absorption spectrum of the species produced, with highest peak at 300 nm and minor peaks at 480 and 520 nm, was found to be similar to that produced after the reaction of bromide radical anion and histidine, which was confirmed to be that of HR (3). Based on this, therefore, it can be concluded that HR was produced from the reaction of azide radicals with histidine, just like the reactions between histidine and the halide radical anions (3). The HR was produced according to be reaction 7 below. HisH + N3. → His.+ N3- + H+ -----------7 Even though the highest peak of absorption of the HR is at 300 nm, the rate of its formation was studied at 515.8 nm i.e. in the visible region of the spectrum rather than in the ultraviolet region, which is not very suitable for use in the kinetics programme (9). The rate of formation of HR (k1=1.46 x 104 s-1) was fast, and was found to fit the first-order kinetics of the computer kinetics programmes designed for the study. This was because the reaction is pseudo first - order, as the rate of the reaction (reaction 7) depends only on the concentration of azide radicals, since histidine was present in excess when compared to the concentration of the radicals. From reaction 6, the concentration of azide radicals produced would depend on the concetration of the OH radicals formed. The G value of OH radicals in nitrous oxide-saturated aqueous solution was reported to be 5.6 (13). This means that 100 eV would give 5.6 OH radicals. Therefore, I Gray would give 3.5 x 1017 molecules dm-3 or 5.8 x 10-7 moldm-3 of OH radicals. Since the dose used in this study was 31 Gray, then 17.98 umoldm-3 of OH radicals was produced. If all the OH radicals produced reacted with azide ion to form azide radicals as expected due to the excess azide ion present, then the concentration of azide radicals was also 17.98 umoldm-3. This was quite low when compared to the concentration of histidine i.e.1 mmoldm-3 or 1000 umoldm-3, and hence the dependence of the rate of reaction 7 on the azide radicals concentration alone. The fact that histidine has not been reported to be involved in electron transfer reaction with other amino acid residues in an enzyme means that any HR formed is most likely to be as a result of direct reaction between a free radical and histidine residue. The formation of HR when a free radical reacts with an enzyme indicates that the enzyme contains a reactive histidine residue, which could be involved in the activity of the enzyme or not. This could also mean that the enzyme contains histidine residue, which is not buried in the 3 dimensional structure of the enzyme protein. If the histidine residue is involved in the activity of an enzyme, then its modification to HR would lead to the inactivity of the enzyme. This is as a result of the modification of the imidazole ring of the amino acid, which would make it to no longer accept or donate proton to a substrate. This makes it incapable of being involved in catalysis by general - acid or general - base mechanism. REFERENCES
© 2003 Nigerian Society for Experimental Biology. The following images related to this document are available:Photo images[bk03003f3.jpg] [bk03003f2.jpg] [bk03003f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}