 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biokemistri, Vol. 13, No.1, Jan, 2003, pp 23-30 Synergistic interaction between two linear inhibitors on a single enzyme: Vanadate and L-phenylalanine inhibition of rat liver alkaline phosphatase Sylvia O. MALOMO1, Femi J. OLORUNNIJI1*, Rotimi O. ARISE2, Joseph O. ADEBAYO3, Olaniyi T. ADEDOSU4 and Adewale A ODUTUGA2 1Enzyme Research Unit, Department of Biochemistry, Faculty
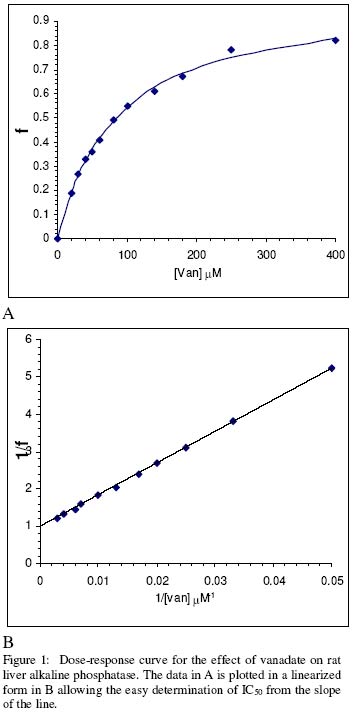
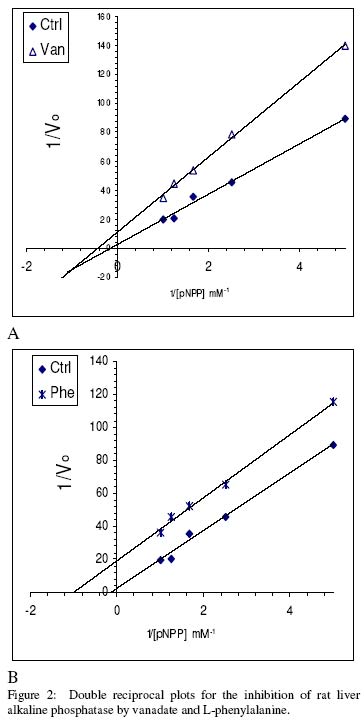
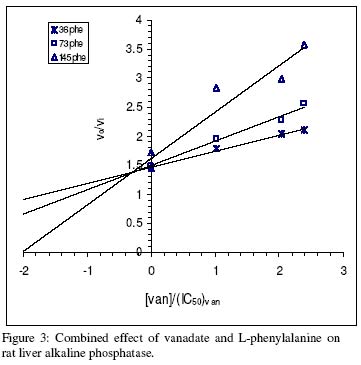
of Science, University of Ilorin, Ilorin, Nigeria Received 27 September 2002 Code Number: bk03004 Abstract The combined effect of two linear inhibitors of rat liver alkaline phosphatase (ALP). vanadate (Van) and L-phenylalanine (L-phe) were studied using a modification of the common Yonetani-Theorell procedure proposed for studying synergistic inhibition. The modes of inhibition of ALP by Van and L-phe as analysed using the double reciprocal plots of the Michaelis-Menten equation were mixed and uncompetitive inhibition respectively. Analysis of the combined effect of the two inhibitors showed that their inhibitory effects were mutually enhanced. The mechanistic aspects and practical applications of the procedure are discussed. Key words: alkaline phosphatase, synergistic inhibition INTRODUCTION Kinetic and mechanistic studies on the combined effect of two linear inhibitors on a single enzyme are of vital interest. It is often necessary to determine if different inhibitors bind to the same site on an enzyme. Such exclusive binding is often investigated by examining how the enzyme responds to the simultaneous effect of both inhibitory ligands. (Cook et al, 1994) The mechanisms of catalysis of enzymes forming ternary complexes have also been investigated using this strategy. (Reckskowski and Ash, 1994) Of vital importance is the potential for enhanced chemotherapeutic effect when two different antagonists enhance their mutual effects when applied to a receptor. Positive synergistic interactions have been reported as a possible new strategy in optimizing chemotherapeutic effects (Scholten et al, 1997) and it is likely that synergy will receive greater attention in the coming years as a practical strategy in rational drug design. It is therefore important to gain proper insight into the mechanism of synergistic interactions. Synergistic response is obtained when the combined effect of two agents deviates from that expected from the dose-response curves of the agents. Hence, the terms 'positive synergism' and 'negative synergism' describe situations in which the two inhibitors enhance and decrease their effects respectively. Positive synergism is often simply referred to as synergy and negative synergism as antagonism. Several kinetic models have been proposed for the analysis of the combined effects of two or more inhibitors on the same enzyme. (Martinez-Irujo et al, 1998) Perhaps the most widely used of the procedures is that of Yonetani and Theorell. (See Martinez-Irujo et al, 1998) This treatment leads to the generation of a 'synergism factor' γ , the value of which provides information on the type of synergistic interaction or its absence. Factor γ describes the mutual influence of the two agents on the binding and effect of one another. Inhibitor binding and hence effect is independent when γ is 1, while values of γ greater or lower than 1 represents mutual antagonism and enhancement respectively. The analysis of combination effects by the method of Yonetani and Theorell involves determining the combined effect of both inhibitors under the condition of keeping one inhibitor concentration constant and varying that of the second. The requirement to have knowledge of the mechanism of inhibitor action via steady state kinetics is a limitation of this procedure and results obtained have often proved to be meaningful only when interpreted in the light of the inhibitors' mechanism of action. Several attempts have been made to simplify the analysis of combined inhibition with a view to reduce the amount of data needed for the analysis and to make the results obtained mechanism-compliant as much as possible. Martinez-Irujo et al (1998) proposed a modification of the Yonetani-Theorell plot in which the IC50 of the inhibitor whose concentration is kept constant is incorporated in the analysis. Here, we have applied the modified procedure of Martinez-Irujo to investigate possible synergism in the effect of two linear inhibitors (Vanadate and L-Phenyalanine) of rat liver alkaline phosphatase. MATERIALS AND METHODS Reagents, chemicals and enzyme substrates p-nitrophenylphosphate was obtained from Sigma Chemical Company, St. Louis, US. Sodium salt of vanadate and L-phenylalanine were products of British Drug House, Poole, UK. All other chemicals used in the study were of high quality research grade. Preparation of rat liver alkaline phosphatase Crude homogenates of rat liver was concentrated for ALP activity using ammonium sulphate according to a modified procedure of Hung & Melnykovych (1977). Rat liver was homogenized in 0.25M sucrose solutions at 4oC and the crude homogenate was centrifuge at 5000 rpm for 20 minutes at the same temperature. To the supernatant fraction was added a 0.55g/ml (4.17M) solution of (NH4)2SO4 gradually with stirring until 30% saturation was reached. Further granules of (NH4)2SO4 were added slowly until 50% saturation was achieved. The precipitate was collected by centrifugation at 5000 rpm for 20 minutes and re-dissolved in 0.1M carbonate-bicarbonate buffers, pH 10.1. The activities of the ALP prepared this way and used in this study were highly reproducible and gave linear results with a correlation level sufficient for kinetic work. Determination of ALP activitiesAlkaline phosphatase activity was measured by the rate of hydrolysis of 5mM p-nitrophenyl phosphate (pNPP) at 25oC in 0.1M Na2CO3/NaHCO3 buffers, pH 10.1 as previously described (Ahlers, 1975). Enzyme activity is expressed as the μM of p-nitrophenol released per minute. Protein concentration was determined using the Folin-phenol method of Lowry et al (1951) with Bovine Serum Albumin (BSA) as standard. Spectrophotometric readings were taken on Spectronic 21 UV-VIS spectrophotometer. Determination of IC50 from dose-response curve of Vanadate and kinetics of inhibition by Vanadate and PhenylalanineIn order to determine the concentration-dependent effect of Van on ALP activity, various concentrations of Van were included in the enzyme reaction mixture and the corresponding activities were recorded. The data were fitted to a dose-response curve according to the following equation (Martinez-Irujo et al, 1998). f =1(/1+IC50/[Van]) Where f is the fractional inhibition exerted by [Van], f = (vo-vi)/vo and IC50 is the concentration of the inhibitor that caused 50% inhibition of the enzyme activity. The IC50 value could be read directly from the plot as the [Van] giving an f value of 0.5. For a more accurate and definite determination of IC50, we transformed the equation by plotting 1/f against1/ [Van]. This gave a straight line with a y-intercept of unity and a slope corresponding to the IC50 value. We found that the two approaches gave very close values for the parameter. In order to determine the effects of Van and L-phe on Michaelis-Menten kinetic constants, Km and Vmax of the liver ALP, the substrate-dependent change in enzyme activity was studied in the absence and presence of the inhibitors separately. Kinetic constants were determined using the Enzfitter computer software (Elsevier-Biosoft) Measurement of the combined effects of Van and PheThe presence and nature of synergy or otherwise in the combined effect of Van and L-phe were investigated according to the procedure of (Martinez-Irujo et al, 1998). The ratio of uninhibited reaction to the inhibited (Vo/Vi) was determined at fixed concentrations of L-phe and varied concentrations of Van and plotted against [Van]/IC50 at the different fixed concentrations of L-phe. The behaviour of two inhibitors with the potential to affect each others binding and effects is given by the expression (Martinez-Irujo et al, 1988)
Where For the specific case in which I1 is a mixed inhibitor and I2 an uncompetitive inhibitor as is the case in this study, the expression reduces to This generated a series of lines each representing the fixed concentrations of L-phe. The lines intersect at a point corresponding to -γ(1+αKs/[S])/(1+Ks[S]) where α represent the change in substrate affinity induced by Van; Ks, the enzyme-substrate dissociation complex; and factor γ represents the mutual influence of the two inhibitors on the binding of each other. Factor α is calculated from the ratio of Km values for the inhibited and uninhibited reactions RESULTS & DISCUSSIONThe dose-response curves for the inhibitory effect of Van and is shown in Figure 1A. The pattern of inhibition displayed a hyperbolic shape indicating that inhibition of ALP by Van was a saturable process. The double reciprocal transformation of the curve shown in Figure 1b was a straight line from which the slope of 85.47μM corresponding to the IC50 was determined. This value was essentially equal to that of 85. 53μM read directly from the curve in Figure 1A. It is recommended that the double reciprocal plot be used in determining the IC50 value as this gives an unambiguous value read directly from the slope of the curve. The kinetics of inhibition by Van and L-phe were also investigated independently and the results are shown in Figure 2. From the double reciprocal plot shown in figure 2A, it is evident that Van is a mixed inhibitor of rat liver ALP. The Km and Vmax of ALP were decreased from 7.81 to 2.28mM and from 0.45 to 0.09 respectively. The calculated inhibition constants from the plots were 4μM and 26.7μM for kiu and kic respectively. The inhibition constants, kic and kiu, represent the binding affinities of the inhibitor to the free enzyme and the enzyme-substrate complex respectively (Segal, 1975) It appears from the results that the affinity of Van for the ES complex is higher than its binding affinity for the free enzyme. The inhibitory effect of vanadate for phosphate-metabolizing enzymes has been ascribed to its ability to mimic the anionic character of inorganic phosphate. (Stankiewicz et al, 1995) In fact, a mutual antagonistic effect between vanadate and phosphate appears to be a fundamental principle in metalloenzymology. It has been shown that vanadate-containing haloperoxidases rapidly lose their activity in phosphate-containing buffers and that reconstitution of the apoenzyme of bromoperoxidase by vanadate is inhibited in the presence of phosphate. (Tromp et al, 1991). These reports strongly suggest a common binding site for vanadate and phosphate in haloperoxidases and phosphatases. One may thus expect that inhibition of ALP by vanadate would follow a strictly competitive mode. Mixed inhibition of ALP by vanadate reported in this study might be examined in the light of a report by Shirazi et al (1981) who observed deviation from competitive inhibition, and in fact, tendency toward mixed inhibition by a series of phosphate analogues. The authors suggested that the ability of some of the analogues to serve as weak substrates of the enzyme might account for the observed deviations. Perhaps there is the need to analyze the pattern of vanadate inhibition of rat liver ALP via other procedures with finer mechanistic 'resolution power' The enhanced binding of ALP-pNPP complex as seen from this study may suggest that vanadate inhibition of ALP may involve other mechanisms apart from competition with phosphate for binding to the active center. Figure 2B shows that, in contrast to Van, L-phe was an uncompetitive inhibitor of ALP. The parallel orientation of the line representing the inhibited reaction with that of the uninhibited one is diagnostic of uncompetitive inhibition (Cornish-Bowden, 1995) This agrees with earlier reports that rat intestinal alkaline phosphatase is inhibited uncompetitively by L-phenylalanine (Ghosh and Fishman, 1966; Hoylaerts et al, 1992) One diagnostic and crucial test for uncompetitive inhibition is that the inhibitor decreases the Km and Vmax by the same factor. As shown in Table 1, L-phe decreased the Km and Vmax of ALP by similar ratios of 0.20 and 0.29 respectively. The seeming disparity observed in this study can be explained on the basis of deviations due to experimental error. One may point out however, that the values are reasonably close enough to substantiate diagnosis of uncompetitive inhibition rather than other possibilities. The selective binding of the ALP-pNPP complex by L-phe as implied by its uncompetitive mode of action is of some interest. Uncompetitive mode of enzyme inhibition is extremely rare in single substrate reactions. The reported cases of uncompetitive inhibition of ALP by L-phe may provide opportunities for characterizing the mechanistic basis for uncompetitive enzyme inhibition. In the case under consideration, perhaps it would be interesting to investigate what amino acid residues are exposed for hydrophobic interactions in ALP after the substrate is bound. The different, almost exclusive mode of inhibition of ALP by Van and L-phe makes them suitable candidates for synergistic interactions in combined inhibition studies. Martinez-Irujo et al (1998) suggested a modification of the Yonetani-Theorell plot for studying facilitation or hindrance when two different inhibitors simultaneously act on the same enzyme. The authors tested the fitness of the model by analyzing data on the effects of adenosine diphosphate and o-phenanthroline on horse liver ethanol dehydrogenase. The two inhibitors studied were competitive inhibitors of the enzyme and the results obtained from the synergy analysis agreed well with the original Yonetani-Theorell analysis. In this study, the suggested method of Martinez-Irujo et al (1998) was used in analyzing the combined effect of Van and L-phe. Figure 3 shows the combined effects of the two inhibitors (Van and L-phe) on rat liver ALP. The intersection point of the three lines gave a value of -0.261 on the [Van]/(IC50)Van axis from which a value of 0.08 was calculated for factor γ. This value indicates a positive interaction in the combined effect of Van and L-phe in the inhibition of ALP. It is noteworthy that the three lines did not give a perfect meeting point for the estimation of γ. However the average value was taken by solving the simultaneous equation of the lines of best fit passing through the data points. Inability to generate a perfect meeting point for the lines can be attributed to the limitations imposed by working with partially purified enzyme preparations. Ahlers (1975) pointed out the advantage of limited purification of tissue ALP in studies designed for kinetic measurements. In order to carry out such studies under reproducible conditions, it was pointed out that it is better to have large stabilized enzyme preparations than small preparations with a higher activity. In a study on human placental and germ-cell alkaline phosphatase, the molecular mechanism of uncompetitive inhibition by L-phe was linked to its ability to bind to an arginine residue in the active site of the enzyme (Hoylaerts et al, 1992) The authors proposed, based on data from recombinant enzyme in which site directed mutagenesis were carried out, that after binding the phosphorylated substrate to the active site, the guanidium group of Arg-166 that is normally involved in positioning phosphate is redirected to the carboxyl group of L-phe, thus stabilizing the inhibitor at the active site. Bearing in mind the highly anionic character of vanadate, we are suggesting that the mutual enhancement of inhibition by both Van and L-phe may be rationalized on the basis of that report. The negative groups of Van after it is bound to the enzyme active site may create a 'docking site' for binding by L-phe. This, of course presumes that Van binds obligatorily before L-phe when both are simultaneously exposed to the enzyme. The competitive mode of inhibition postulated for vanadate inhibition of ALP in earlier studies (Ghosh and Fishman, 1966; Hoylaerts et al, 1992) and the mixed mechanism we found in this study indeed show that it is likely that Van binds before L-phe. However, there is no direct evidence yet to substantiate this assumption. Hoylaerts et al (1992) also suggested that uncompetitive inhibition of human ALP by L-phe might result from coordination of the amino acid unprotonated amino group with the active site Zn2+ ion, thereby interfering with the hydrolysis of the phosphoenzyme intermediate. This is of significant interest in explaining the mutual enhancement of the inhibitory effect of Van and L-phe. Vanadates with its negative character can readily complex Zn2+ and other divalent metal ions present in the enzyme and necessary for catalytic activity. The findings from this study proved that the procedure of Martinez-Irujo et al (1998) for detecting and characterizing synergistic interactions in combined inhibition could also be applied to cases in which the inhibitors act via different mechanisms. However, it is still important to develop analytical procedures that do not rely on the mechanism of inhibition of either of the combined inhibitors. Progress in this direction will definitely facilitate the use of combined inhibitors in chemotherapy and other mechanistic areas of enzyme studies. REFERENCES
© 2003 Nigerian Society for Experimental Biology. The following images related to this document are available:Photo images[bk03004t1.jpg] [bk03004f1.jpg] [bk03004f3.jpg] [bk03004f2.jpg] |
| |||||||||

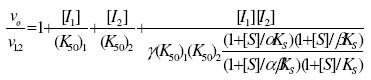
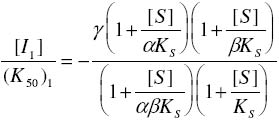
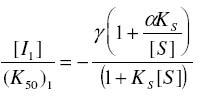
{kind=link}
{kind=link}
{kind=link}
{kind=link}