 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
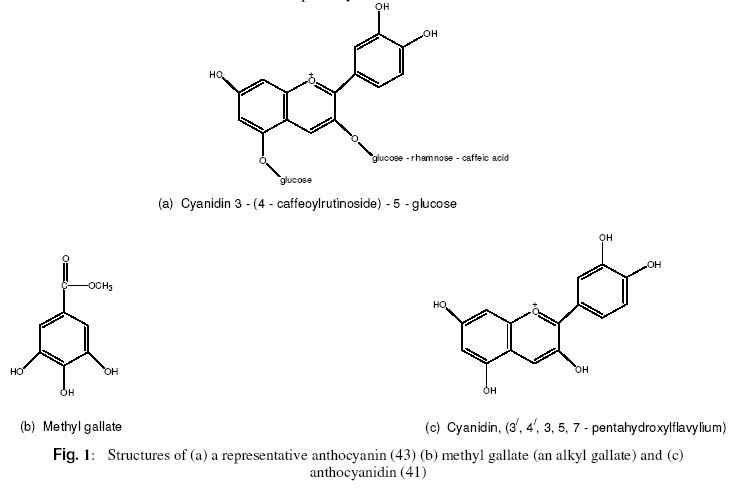
pH Dependent Prevention of Carbon tetrachloride–Induced Lipoperoxidation in Rats by Ethanolic Extract of Hibiscus rosasinensis Petal F. O. Obi* and E. Uneh Department of Biochemistry, University of Benin,
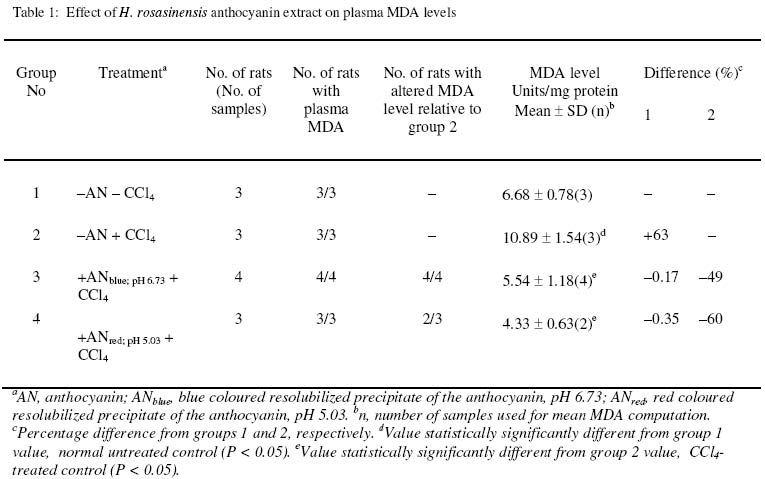
P.M.B. 1154, Benin City, Nigeria. Received 2 December 2002 Code Number: bk03007 Abstract The ability of high (6.73) and low (5.02) pH solutions of lead acetate precipitate of H. rosasinensis petal anthocyanin extract to prevent carbon tetrachloride (CCl4) induced lipid peroxidation in rats has been investigated using plasma malondialdehyde (MDA) level as an index of the process. Relative to its value in the plasma of CCl4 treated extract-free rats, the high pH form (pH 6.73; blue pigment) of the resolubilized precipitate significantly (P < 0.05) reduced the level of malondialdehyde in the plasma of rats to which it was administered prior to CCl4 treatment. When compared with CCl4 – treated, extract-free rats, statistically significant (P<0.05) reduction in the level of MDA was also observed in the plasma obtained from rats treated first with the low pH form (pH 5.02; red pigment) of the resolubilized precipitate before CCl4 exposure. These data suggest that H. rosasinensis petal partially purified anthocyanin extract may be protective against carbon tetrachloride-induced lipoperoxidation. Key Words:Lipoperoxidation, carbon tetrachloride, Anthocyanin, Hibiscus rosasinensis, Malondialdehyde INTRODUCTION Anthocyanins have long been known to be involved in plant pigmentation. They are responsible for the brilliant orange, pink, scarlet, red, mauve, violet and blue colour of flower petals and fruits of higher plants(1)and cortex colour of some varieties of yam, Dioscorea alata L and Dioscorea tryphida L(2,3,4). Anthocyanins (Fig. 1a) and their aglycones, the anthocyanidins (Fig. 1c) are part of the C15 phenolics known collectively as flavonoids – a series of C6 – C3 – C6 compounds. Anthocyanidins are derived in nature from the flavylium structure by the addition of hydroxyl groups some of which are subsequently methylated (5). Plant phenolics such as alkyl gallate (methyl – or propyl gallate) – Fig. 1b are natural antioxidants (6). Flavonoids, among which are anthocyanins (7) are widely distributed in the plant kingdom and it is recognized that some of them due to their phenolic structure show antioxidant activity towards a wide range of oxidizable substances (8). Hence it is believed that one of the many possible functions of these flavonoids is the protection of plants against free radical mediated damage (8). Also on account of their antioxidant potency vitamins C and E along with flavonoids have been linked to low incidence of atherosclerosis (9,10). Frankel et al (11) have reported on the ability of phenolic substances in red wine to inhibit the copper – catalysed oxidation of LDL, which is thought to be a crucial event in the pathogenesis of atherosclerosis. Other proponents of the involvement of flavonoids in the prevention of LDL oxidation and the attendant reduction in atherosclerosis are Durak et al. (12), Maxwell et al (13) and Renaud and DeLorgeril (14). Indeed the flavonoids in red wine are being fingered as the likely molecular entities responsible for what has been described as the “French Paradox” – the lower incidence of coronary atherosclerosis in the French population than that of other Western population even though the former consume a more fatty diet. Carbon tetrachloride induces lipid peroxidation and liver damage. The peroxidative effect is associated with increased production of malondialdehyde, diene conjugates (15, 16, 17, 18, 19) as well as 4-hydroxynonenal (4HNE) (20,21,22) which are degradation products of polyunsaturated fatty acid hydroperoxides. The lipid peroxidation and liver damage happens when trichloromethyl radical (*CCl3) obtained from CCl4 reacts with molecular oxygen forming trichloromethylperoxy radical (CCl3 – O – O*) (23,24,25,26,27,28). Therefore, its effect on the liver provides a model with which natural plant products have been assessed for antioxidant and/or hepatoprotective functions (29). In our previous investigations, the hepatoprotective effect of H. rosasinensis petal anthocyanin (30,31) and its anthocyanidin (32) was assessed by using plasma enzyme levels as indices of damage and protection. However, it is possible that our results are a reflection of plasma enzyme inhibition by the extract and not antilipoperoxidative action per se. Hence in this present investigation, we aimed to resolve these conflicting views by measuring malondialdehyde level as the indicator of CCl4 – induced peroxidative reaction and prevention of the same process by H. rosasinensis petal anthocyanin extract. MATERIALS AND METHODS Experimental animals and materials White albino rats (Wistar strain) of an average weight of 169g bred in the Animal Unit of the School of Pharmacy, University of Benin, were used for this study. They were divided into four experimental groups of 3 or 4 rats and housed in standard rat cages (Techniplast, Italy). Absolute ethanol, chloroform, ethylacetate and sodium hydroxide were purchased from BDH Chemicals Ltd (Poole, England). The other chemicals are carbon tetrachloride (Hopkins and Williams), and 2 – thiobarbituric acid (E. Merck, Darmstadt). Corn oil (Mazola, produced for CPC, UK). Extraction and purification of anthocyanin The modified method of Takeda et al. (33) which involves the use of a mixture of formic acid, ethanol and distilled water (1:10:9) instead of 1M HCl; methanol and water (1:10:9) as described previously (31) was used for H. rosasinensis petal anthocyanin extraction. The initial extract from 50g petal in 170ml of the modified solvent was transferred into a separatory funnel and “washed” three times with equal volumes of ethylacetate to remove flavones. The third volume of the ethylacetate that was added and the extract were mixed thoroughly in the separatory funnel and left overnight. The ethylacetate-free layer containing the partially purified anthocyanin was thereafter obtained. 50ml of the ethylacetate-free extract and 50ml of 0.5% neutral lead acetate solution were mixed and kept at 4ºC for 48 hours to ensure complete precipitation of anthocyanin. About 90ml of a blue supernatant was discarded. The precipitate was resuspended in the remaining supernatant and transferred to a test tube. The content of the test tube was thereafter centrifuged at 5000 rpm for 5 minutes. A blue supernatant and a dark precipitate (anthocyanin) were obtained and the supernatant was discarded. 5 millilitres of 0.5% solution of sulphuric acid was added to the precipitate to remove lead as lead sulphate (PbSO4) and the precipitate simultaneously resolubilized to give a red solution. The mixture was filtered to remove the PbSO4 and the filtrate made up to 39 ml with distilled water. 2ml of the filtrate was evaporated to dryness in a pre-weighed watch glass to estimate the amount of partially purified anthocyanin obtained. 1.95 ml of absolute ethanol was added to the remaining 37 ml of the diluted filtrate to obtain a solution of the anthocyanin in 5% aqueous ethanol. The extract in 5% aqueous ethanol was divided into two equal parts. One half had a pH value of 5.02 and retained the red colour. The other half was made less acidic by increasing the pH to 6.73 with a few drops of 5M sodium hydroxide solution. Both were put into brown plastic bottles to avoid possible effect of sunlight and left at -20ºC until required. Treatment of animals with partially purified anthocyanin and carbon tetrachloride Rats in group 1 (normal control) were not treated with either anthocyanin or carbon tetrachloride but were given neat 5% aqueous ethanol (2.5 ml/kg body weight) orally, since 5% aqueous ethanol was used as vehicle for anthocyanin administration. Rats in group 2 (test control) were treated with CCl4 alone. An aliquot (1.02ml/kg º 30 mg CCl4/kg) of a mixture of CCl4 and corn oil (1:50 v/v) was administered subcutaneously to the rats. Rats in group 3 (high pH test group) were pretreated with the blue anthocyanin fraction (pH 6.73) orally at a dose of 500mg/kg once daily, five days a week for two weeks. Rats in group 4 (low pH test group) were treated with the red anthocyanin fraction (pH 5.02) in the same manner and for the same duration described for group 3 rats. At the end of the first week, rats in groups 2, 3 and 4 received CCl4 in corn oil (29) as described above. The CCl4 treatment was repeated at the end of the second week when the experiment was terminated. Preparation of plasma samples 24 hours after the last CCl4 treatment, each rat was anaesthetised in a chloroform saturated chamber. The thoracic region was opened to expose the heart. Blood was obtained by heart puncture by means of a 5 ml hypodermic syringe and needle and placed in ice-cold heparinized 5 ml sample tubes. The blood samples were centrifuged at 5000 rpm for 5 minutes. The plasma samples were collected and stored at -20ºC (at most overnight) until required for malondialdehyde analysis (34). Determination of plasma protein and malondialdehyde Total plasma protein was quantified by the biuret method. The method adopted for the analysis of malondialdehyde was that of Hunter et al (35) as modified by Gutteridge and Wilkins (34). Blood plasma (0.2 ml) was put in a test-tube containing 3 ml glacial acetic acid and 3 ml of 1% thiobarbituric acid (TBA) in 2% NaOH. The mixture was placed in boiling water for 15 minutes, allowed to cool and the absorbance of the pink coloured product was read at 532nm (34,36). Statistical analysis The data represent the mean ± SD for n number of rats (see table 1 column 6 for the values of n). The mean values of the various treatment groups were compared using ANOVA and the least square difference test (37). The significance level was set at P < 0.05. RESULTS Changes in plasma MDA level induced by CCl4, 24 hours after administration and the effect of prior treatment with the low and high pH forms of the resolubilized lead precipitate of H. rosasinensis petal anthocyanin extract are presented in Table 1. A significant increase in the level of plasma MDA was observed in the CCl4 treated anthocyanin-free rats (P < 0.05). Also observed was a significant (P < 0.5) decrease in plasma MDA level in groups 3 and 4 rats due to prior treatment with the low and high pH forms of the anthocyanin extract, respectively, when compared to CCl4 – treated anthocyanin-free rats. DISCUSSION In this study plasma MDA was used to measure both CCl4 – induced lipoperoxidation and protection of H. rosasinensis anthocyanin against the same effect of CCl4 in rats. In agreement with the report of previous workers (15, 28) our results show that CCl4 caused elevation in plasma MDA which is indicative of lipoperoxidation (20, 38). Treatment of two different groups of rats, one with high and the other with the low pH form of the resolubilized partially purified anthocyanin before CCl4 treatment caused 49% (ANblue, pH 6.73) and 60% (ANred, pH 5.02) less lipoperoxidation respectively than with CCl4 alone (Table 1). See in table 1, that the plasma MDA of groups 3 and 4 rats which were exposed to the high and low pH forms of the extract respectively, before CCl4 treatment are lower than that of group 2 rats exposed to CCl4 only. One of the various mechanisms that have been proposed for CCl4 –induced tissue injury is that a trichloromethyl radical (·CCl3) is produced from carbon tetrachloride by reductive dechlorination (18, 39). The trichloromethyl radical abstracts a hydrogen atom from polyunsaturated fatty acids to form chloroform and a lipid radical, the lipid radical may then react with molecular oxygen to initiate lipid peroxidation which is thought to ultimately cause the cytotoxic response (17,40). Since the mechanism described here suggests an underlying process of oxidation, we hypothesized that if anthocyanin possesses antioxidant action, it would prevent lipid peroxidation. The result presented in the table clearly demonstrates that anthocyanin from H. rosasinensis is an excellent antagonist of CCl4 – induced lipoperoxidation. There are a wide variety of naturally occurring compounds that can protect tissues from chemically – induced damage. Among these are antioxidants such as carotenes, tocopherol, catechols etc (19). It is therefore possible that if these other principles were coextracted, they alone or in conjunction with the anthocyanin are responsible for the impaired lipoperoxidation. However as discussed previously (31) based on the extraction procedure and the nature of the solvents, these other antioxidants are not likely to have contributed meaningfully, if at all, in the impairment of lipoperoxidation. Furthermore, in the present investigation, anthocyanin was precipitated with neutral lead acetate. The water soluble naturally occurring antioxidant that could be present in the extract before the precipitation stage are ascorbic acid and catechol, that is, if they occur naturally in Hibiscus rosasinensis. However, we have not come across any information in the literature that ascorbic acid and catechol coprecipitates with flavonoids when lead acetate is employed as the precipitating agent. As is evident from the table rats in group 1 that were not treated with CCl4 and anthocyanin had higher MDA levels than those in groups 3 and 4 that were treated with CCl4 and anthocyanin. This is an indication that although CCl4 at a dose of 30mg/kg causes lipid peroxidation (table 1, group 2) it does not appear to have done so sufficiently in the presence of the partially purified anthocyanin extract (table 1, groups 3 and 4). In vivo, lipid peroxidation and attendant malondialdehyde production can be caused by enzymatic and nonenzymatic free radical generating events in the absence of toxic substances (15). Hence, lipid peroxidation in group 1 rats must have arisen from both natural and ethanol initiated processes (see section on animal treatment for the reason for ethanol inclusion). In group 2, lipid peroxidation is attributable to both natural and CCl4 – initiated events. In groups 3 and 4 lipoperoxidation is due to natural events and events associated with the metabolism of the toxic substances, ethanol and CCl4. It is evident from the table that the plasma MDA levels in groups 3 and 4 are 0.17 and 0.35% respectively lower than that of group 1 as well as 49 and 60% lower than that of group 2 due to the anthocyanin treatment. The mechanism by which H. rosasinensis anthocyanin and/or its anthocyanidin (probably generated in vivo) effect this antilipoperoxidative action is yet to be elucidated. Nevertheless Figure 1c shows that H. rosasinensis anthocyanidin, cyanidin (41) is highly hydroxylated. The same is true of its glycosylated derivative, the anthocyanin. They share this structural feature with alkyl gallate – another highly hydroxylated phenolic compound with effective antioxidant function (6). Their hydroxyl groups probably play crucial role in the lipoprotective effect. Furthermore, catechol (o-dihydroxybenzene) is a very powerful reducing agent (42). Catechol-like structure is evident in alkylgallate (Fig. 1b) in terms of hydroxyl group distribution. If this feature is the basis of the antioxidant action of alkylgallates it may also confer this ability to H. rosasinensis anthocyanin/anthocyanidin since the same feature is present in the latter (Figs. 1b and 1c). This strikes us as probable basis of the influence of pH on the degree of reduction of CCl4 – induced lipoperoxidation (Table 1) reported in this paper. The present study was designed to show whether the hepatoprotective action of H. rosasinensis petal anthocyanin demonstrated previously (30, 31) was due to antioxidant effect and not a case of plasma enzyme inhibition. The results presented here show that CCl4-induced lipid peroxidation in rats was impaired by H. rosasinensis anthocyanin extract. So the hepatoprotective action of H rosasinensis petal anthocyanin extract demonstrated previously (30, 31) is evidently attributable to its antioxidant action. REFERENCES
The following images related to this document are available:Photo images[bk03007f1.jpg] [bk03007t1.jpg] |
| |||||||||

{kind=link}
{kind=link}