 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biokemistri, Vol. 15, No.2, Dec, 2003, pp. 57-66 Prevalence of Haemoglobinopathy and Malaria Diseases in the Population of Old Aguata Division, Anambra State, Nigeria Peter N. UZOEGWU 1 and A. E. ONWURAH 2, 1Department
of Biochemistry, Tropical Diseases Research Unit, Sickle Cell Laboratory, University of Nigeria,
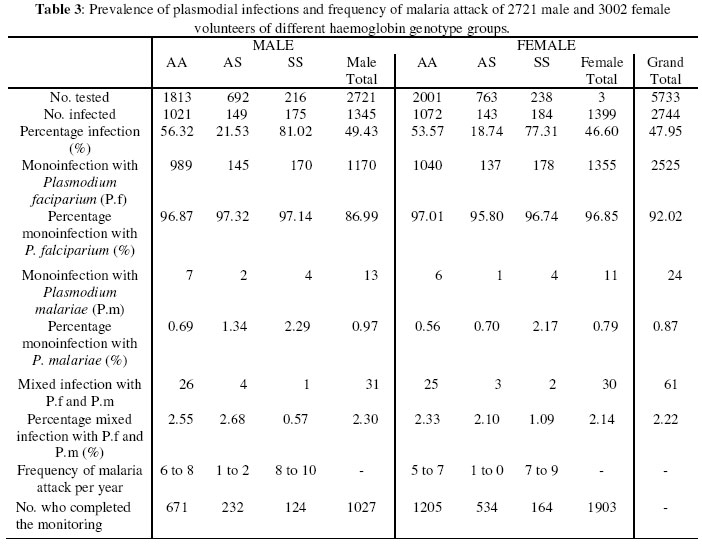
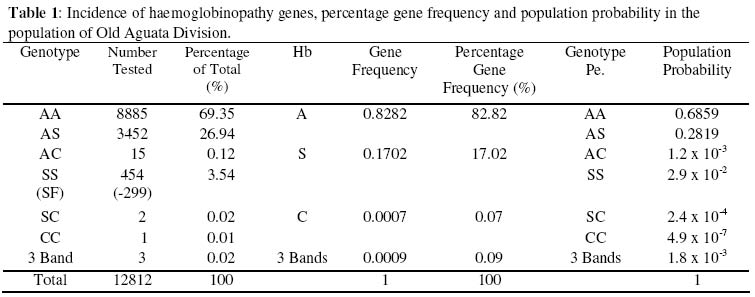
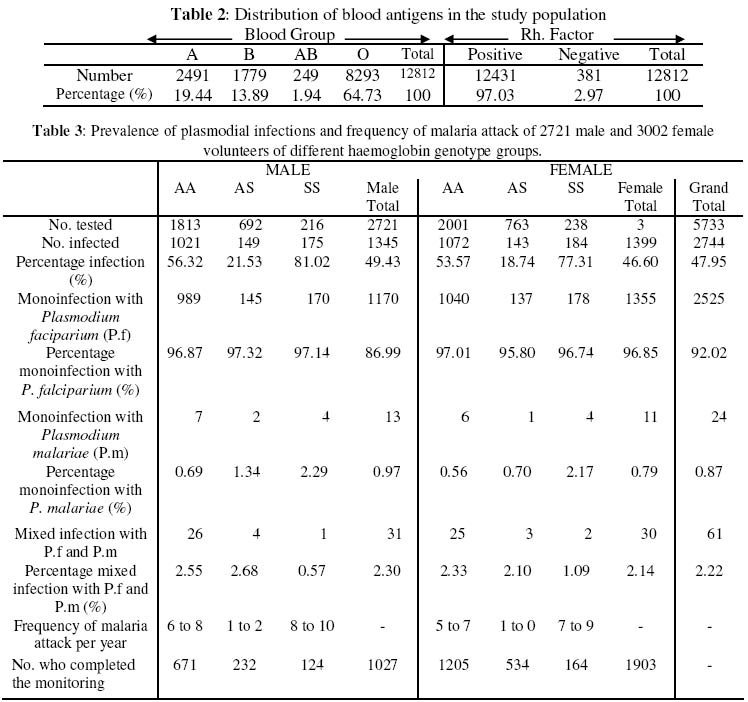
Nsukka Received 6 August 2003 Code Number: bk03021 Abstract The prevalence of haemoglobinophy and malaria were determined in 12812 inhabitants (5227 males and 7585 females) of the twenty-one (21) autonomous communities of Old Aguata Division, Anambra State, Nigeria. The intention was not only to reveal their most current incidences but also to adduce the most probable reason(s) for the widespread myths peddled about sickle cell disease (SCD) in the area. Results revealed high incidences of sickle cell anaemia (3.54%) and sickle gene trait (26.94%), low incidences of sickle haemoglobin C (0.02%) and haemoglobin CC (0.01%) diseases and an average high prevalence of malaria (47.95%). Most of the malaria attack was caused by Plasmodium falciparum (P.f) infection. The possession of over 20% of persisting concentration of foetal haemoglobin (HbF) by 65.86% (299/454) of identified sickle cell anaemia (SCA) subjects and the striking limited knowledge of SCD and its method of control by the inhabitants of the communities are inconsonance with the high incidence of SCA in the population. The low percentage HbC gene frequency (0.07%) computed from the data could explain the rarity of HbSC and HbCC diseases in the population. Their probable frequencies of occurrence are of the order of one in every 20,000 and 600,000 persons respectively while that of SCA is of the order of one in every 200 persons. Further results showed that dominant homozygotes (HbAA) were more susceptible to plasmodial parasite infection than sickle heterozygotes (HbAS) while recessive homnozygotes (HbSS) were most vulnerable to malaria than the other two members of genotypic groups. Furthermore, random administration of sub-standard and under-dose of anti-malaria drugs by patent medicine dealers operating in the area is suspected to be the main factor responsible for the emergence of the observed anti-malaria drug-resistant strains of P.f which is not in consonance with high prevalence of malaria in this area. Key words:Sickle Cell Disease, Malaria, Sub-standard Anti-malaria Drug INTRODUCTION Sickle cell disease and malaria are the worst tropical diseases most prevalent in Nigeria in particular and Africa in general. The two disease states manifest similar symptoms of severe haemolytic anaemia, fever, weakness and high infant mortality rate. Sickle cell disease is a genetic disorder arising from a point mutation in the β-globin gene (1). The mutation, GAG to GTG, leads to the displacement of an acidic amino acid (glutamic acid) by a neutral amino acid residue (valine) at the sixth position of the amino acid sequence of the β-globin chain to form sickle haemoglobin (HbS). The formation of another abnormal haemoglobin C is achieved by the displacement of the sixth amino acid residue, glutamic acid, in the native β-globin chain by the basic amino acid residue, lysine. At low oxygen tension, such as is usually found in tissues during plasmodial or other infections, HbS could polymerise into higher order aggregates (2) which rigidifies and distorts the biconcave egg-shape of normal red blood cells (RBCs) into sickle shape. The resultant effect of the formation of rigid sickled red blood cells is their aggregation in tissues or organs to block the microvasculature which could cause tissue or organ damage, intractable pains or even death (3). Malaria, on the other hand, is caused by an intracellular protozoan parasite, known as Plasmodium. They are usually injected into individuals by female anopheles mosquitoes during their blood meal. The most lethal form of malaria is caused by Plasmodium falciparum (P.f.). It is known that SCD is widely spread in the high malaria endemic areas of Nigeria (4). Many Nigerians are known not to have understood clearly the generation, presentations and the management of SCD perhaps because of the observed high level of ignorance of the disorder among Nigerians (5). This ignorance might have contributed to the gross misconception of the presentations of SCD among Nigerians, at least before 1980 that SCD is caused by evil spirits witches or demons (5). Sickle cell disease and malaria are well spread in Nigeria but their correct prevalences are not definitively known. Although Nigeria was reported to have the highest incidence of SCA in Africa and the world in general (6) more than three decades ago, only limited information on the incidences of haemoglobinopathies and their traits are available from some localised surveys carried out many year ago, among the Igbos of Eastern Nigeria, (6-12) and other ethnic groups in Nigerian (13-15). These workers reported different incidence rates for different parts of Nigeria and adduced different incidence rates observed. Since no current information is available on the incidences of different SCDs and their traits in the Old Aguata Division and since a lot of mysterious infant deaths were verbally reported by the people of the area, it became necessary that the twenty-one (21) autonomous communities that make up the Old Aguata Division be randomly surveyed for the distribution of the haemoglobinopathy genes and traits and for the prevalence of malaria in the area. The information obtained would not only be useful to the Nigerian government in the planning of her much discussed health-for-all in the year 2010, but also be of use in the enlightenment of the people of the area about SCD, its prevention, control and management. MATERIALS AND METHODS Study Area The survey was carried out between 1992 and 1996, in most of the twenty-one (21) autonomous communities that make up the Old Aguata Division, now divided into three local government areas, namely Aguata, Orumba North and Orumba South, for administrative convenience. The Old Aguata Division located about twenty kilometres east of Awka in AnambraState, has a population of about 1.5 million. The inhabitants are mainly farmers and traders. However, the autonomous communities have high concentrations of primary, secondary and two tertiary institutions. Although the inhabitants had limited knowledge of SCD, like other Nigerians, the “incurable disease” that presented frequent anaemia was described as “Oya Ota Obara” (the disease that consumes blood), perhaps because it causes frequent anaemia. Furthermore, because the disease was incurable, it was misconceived to be caused by witches and wizards or charms prepared by imaginary enemies. The misconception is strongly believed since the disease presents severe anaemia which isbelieved to be caused by witches sucking the blood of the patients. Selection of Subjects Enlightenment lectures on sickle cell and malaria diseases were first delivered for one hour to the assembly of inhabitants of the respective communities in order to create the awareness of the diseases. The participants were mainly students and members of staff of primary, secondary and tertiary institutions located at the communities as well as other residents of the respective communities. After each lecture, volunteers who would want to know their genotype, blood group and Rhesus factor and/or take part in the bimonthly plasmodial infection monitoring were selected. Their residents/classes in schools were identified for easy access. Proforma for their personal data and previous knowledge of the diseases were completed by or for each individual participant. A total of 12812 subjects 5227 males and 7585 females of comparable age range, between 6 and 43 years were selected for the tests at the beginning of the study. Blood Sample Collection Blood samples (0.5ml) were drawn by venipuncture, from the volunteers using 2ml volume styrile syringe and needle (21G x 1½”) (Terumo Leuven Belgium). The blood samples were gently poured into 2ml volume polysterene tubes containing 20µl 2% (w/v) ethylenediamine tetracetate (EDTA) Disodium (Sigma) as an anticoagulant. The samples were then carried in a cool compartment to the University of Nigeria for analysis. The tests were either done that day or the blood samples stored at 4oC in a refrigerator till the following day. Preparation of Haemoglobin Lysate About 0.3ml of the blood sample was centrifuged at 3000g for ten minutes with Hittich universal bench centrifuge, model 1200. Plasma was aspirated off while the precipitate (blood cell layer) was resuspended in equal volume of normal saline (0.85% NaCl, w/v) for washing. The washing was repeated three times and finally resuspended in equal volume of normal saline. The red blood cell suspension (40μl) was mixed with equal volume of distilled water to lyse the blood cells. The resulting haemoglobin lysate (the lysate) was used for haemoglobin genotype determination. Uncoagulated whole blood could also be diluted with equal volume of distilled water to form the lysate that could also be used for genotype determination. Electrophoresis of Haemolysate Haemolysate electrophoresis was performed according to the modified method of (16) as described below. Cellulose acetate strips were soaked in the working buffer containing 0.012M Tris (hydroxymethyl) aminomethane (Tris, Sigma), 0.0004M Ethylenediaminetetracetic acid (EDTA) Sodium Dihydrate, (Sigma) and 0.0015M borate (BDH), pH 8.9, (Tris-EDTA-Borate buffer). The working buffer was prepared from the stock buffer which contained 0.12M Tris, 0.004M EDTA and 0.015M Borate, pH 8.6, by making a one in ten dilution with distilled water. Haemoglobin lysate was spotted, in duplicate, at about 1cm from one end of well dried cellulose acetate strip. Haemolysates of reference AS and AC individuals were also spotted on the same acetate strip as controls. The two compartments of the electrophoretic tank contained the same working buffer. Whereas one compartment was connected to the anode of the power supply unit (Vokam Shandon), the other compartment was connected to the cathode. The cellulose acetate strip with the haemolysate spots was placed on the runs of the tank to form a bridge. The strip was held in place at the two ends by the buffer-dampened filter paper. A constant voltage of 150V (2mA Current) was maintained through the power supply unit. Adequate separation was achieved within five to seven (5-7) minutes. The haemoglobin spots were visualised directly as pink spots or otherwise stained with Ponseau S stain (0.2%, w/v in 5% acetic acid) and then destained with distilled water until the background of the paper became white. Foetal Haemoglobin Determination Foetal Haemoglobin of all the 454 sickle cell anaemia patients was determined by the alkali denaturation method as reported in (17). Blood Antigens Determination The blood group and Rhesus factor antigens of 12812 volunteers were determined by Coombs direct agglutination method as reported in (18) using commercially purchased ABD antisera (Anti A, Anti B and Anti D, Antisera, LDP, UK). A drop of whole uncoagulated blood was mixed thoroughly with a drop of respective antiserum and the mixture rocked gently for two (2) minutes. Agglutination indicated positive reaction while absence of agglutination indicated negative reaction. Diagnosis of Malaria Parasite Infection A total of 5723 volunteers, 2721 males between 14 and 26 years of age and 3002 females between 16 and 30 years of age and of mixed genotype groups (AA, AS and SS) were tested for plasmodial infections and then monitored for the frequency of malaria attack for one year. These volunteers were selected from the 12812 subjects who willingly submitted themselves for the genotype and malaria tests. The selected subjects, who showed no overt symptoms of malnutrition or any specific nutritional deficiency were clinically examined for malarial symptoms such as relapsing fever, headache, shivering, overt signs of anaemia, tiredness and loss of appetite by a team of medical officers working at the Government General Hospital Ekwulobia, Aguata local government area of Anambra State, one of the autonomous communities under survey. Malaria attack was confirmed by microscopical examination of Giemsa-stained thin and thick blood films at the Diagnostic Laboratory of the Department of Microbiology, University of Nigeria, Nsukka. Determination of Frequency of Malaria Attack The frequencies of malaria attack of the 2721 male and 3002 female volunteers were determined by a biweekly monitoring of plasmodial infection on them. The thin and thick blood films of the selected subjects were made every two weeks and microsopically examined for the presence of plasmodial parasites for a period of one year. Not all the volunteers who started the monitoring completed the one year monitoring. The numbers who completed the monitoring in each genotype group are shown in Table 3. RESULTS Incidence of Haemoglobin Genes The incidences of SCA and sickle cell trait in the population
of Old Aguata Division were computed to be 3.54% and 29.94% respectively. However,
65.86% The percentage gene frequencies of HbA, S, C and three-band pattern, computed from the population probabilities of AA, AS, AC, SS and SC are 82.82%, 17.02%, 0.070% and 0.09% respectively (Table 1). Blood Antigen Distribution As shown in Table 2, blood group antigen, AB, exhibited the least percentage incidence of 1.94% while blood group antigen 0 (universal donor) manifested the highest incidence of 64.73%. The incidence of blood group A antigen was computed to give 19.44% while that of blood group B antigen was calculated to be 13.89%. Individuals without Rhesus factor antigen made up only 2.97% of the population while subjects positive for Rhesus factor antigen exhibited the high incidence of 97.03%. Plasmodial Parasite Infections The results of the parasitological examination for the presence of plasmodial parasites in 5723 volunteers (2721 males and 3002 females) of comparable age range between 14 and 30 years and of mixed genotype groups AA, AS and SS, displayed in Table 3, demonstrate that 49.43% and 46.60% of males and females respectively were infected with plasmodial parasites, giving an overall infection of 47.95%. Comparing the plasmodial parasite infection in different genotype groups, the results revealed that 56.32%, 21.3% and 82.02.02% of AA, AS and SS males respectively and 53.57%, 18.74% and 77.31% of AA, AS and SS females respectivelywere infected with plasmodial parasites. These results demonstrate that sickle cell anaemia subjects (HbSS) were most vulnerable to plasmodial parasite infection than the members of other genotype groups. Sickle heterozygotes (HbAS) were least susceptible. Furthermore, females were observed to be less susceptible to plasmodial parasite infection than males. Table 3 also shows that 96.87%, 97.32%, and 97.14% of all infected male AA, AS and SS subjects respectively and 97.01%, 95.80% and 96.85% of the infected female AA, AS and SS individuals respectively showed monoinfection with Plasmodium falciparum (P.f.). Similarly, only 0.69%, 1.34%, and 2.29T of the infected male AA, AS and SS respectively and 0.56%, 0.70% and 2.17% of the infected female AA, AS and SS subjects respectively displayed monoinfection, with Plasmodium malariae (P.m). Mixed-infections with P.f. and P.m were manifested in 2.55%, 2.68% and 0.57% male AA, AS and SS respectively and in 2.10%, 1.09% and 2.11% female AA, AS and SS subjects respectively. No infection with any other plasmodial specie was revealed in the population. By this result, Plasmodium falciparum is therefore the main cause of malaria attack in the study area. Frequency of Malaria Attack The frequencies of malaria attack as monitored revealed that AS in both male (1 to 2 times per year) and females (once or none per year) was least infected, when compared with the frequencies of other genotype groups. Male and female SCA patients showed the highest frequencies of 8 to 10 and 7 to 9 infections in a year respectively Clinical Examination Clinical examinations of infected individuals revealed that P. falciparum infection presented more severe and more serious symptoms than P. malariae infection which presented milder and slower but more progressively persisting symptoms. DISCUSSION The incidence of high SCA and sickle trait revealed among the population of Old Aguata Division of Anambra state are surprising and inconsistent with the lower incidences reported among Nigerians in earlier studies (9,10,13-15). However these results are in consonance with the incidences of SCA (3.81%) and sickle trait (29.01%) recently observed in the University of Nigeria, Nsukka community, predominantly inhabited by Igbos (19). This high incidence of SCA is surprising given that the survey was done at the period (1992-1996) when Nigerians were expected to have become more aware of the disease and hence applied effective control measures to prevent the reproduction of children with SCA. Sickle cell education was seriously shared among many Nigerian community organisations and members as from mid 1980s which could have made Nigerians more aware of the disease and consequently led to the reduction of the incidence of sickle cell disease. One, therefore, could have expected a lower incidence of SCA at this period of study. This expectation could be supported by the fact that miseducation about SCA that occurred in the 1970s among black Americans was responsible for higher prevalence of SCD which became lower in the 1990s when the black Americans became more aware of the disease (20). However, in Nigeria, particularly in the rural populations, the knowledge of SCD was still shallow and limited during this period of study (4). This fact was confirmed by the information extracted from personal interviews of the volunteers and from their completed proforma, showing that 71% of the people of the study area had limited knowledge of SCA. As a result of this ignorance implied above, a lot of misinformation generated therein, could have been responsible for the wrong ascription of he high infant mortality rate caused by SCA to evil spirits, witches and demons or to deities revenging the guilts of affected family members. Misinformation about the disease could have as well led to wrong diagnosis and misapplication of SCA management methods which, in turn, could have precipitated high infant mortality rate reported by many residents of the area. These factors could have lowered the incidence of SCA in this area. However, the high incidence of sickle trait coupled with the revealed gross ignorance of the disease among the people of the area, many of whom are rural dwellers, could have led to many haemoglobin-mismatched marriages with its attendant risk of reproducing children with SCA. This factor could also explain the high incidence of SCA observed during this study. Other factors such as persisting high concentration of foetal
haemoglobin (HbF) could invariably influence the prevalence of SCA in a population. There
are conflicting opinions about the level of influence persisting HbF concentration
could have on the severity of SCA crisis. Some scientists reported that HbF
concentration below 20% did not affect the severity of SCA crises (17,21, 22)
while others observed that HbF concentration could work synergistically with
5’-modulator to determine intracellular polymerisation of HbS which reduces
the severity of sickle cell syndrome (23) or high HbF concentration was responsible
for many more mild features of SCA observed in the Eastern Province of Saudi
Arabia while the more severe crises in the patients from the Western province
was consistent with low HbF level (24). The high persisting level of HbF (18-26%)
observed in 65.86% ( The high incidence of sickle gene carriers (HbAS) identified in this community unschooled in sickle cell disease genetic counselling, could also, most probably, contribute to this high incidence of SCA observed in this community. The high incidence of SCA and malaria in this community appears paradoxical, in that one would have expected lower incidence precipitated by persistent malarial-induced high infant mortality among SCA patients. Malaria and pneumonia are the worst diseases which trigger off severe haemolytic crisis and anaemia in Nigeria (5). Severe anaemia is the main cause of death among sicklers in the East and West Africa (25) and therefore prompt treatment of malaria (26) and other diseases such as acute chest syndrome (27) are very useful in the management of SCA. Although the high prevalence of malaria revealed in this study (47.95%) is lower than that reported (54.91%) for Ghanaian school children (28) both studies agreed that P.f. is the main cause of malaria. The results shown in Table 2 indicate that sickle heterozygotes are 2.7 times less susceptible to plasmodial infections than dominant homozygotes while the persons with SCA are 4 times more susceptible to malaria than sickle heterozygotes. These results are not surprising given that plasmodial infections of SCA persons trigger off a concentration-dependent polymerisation of HbS in RBC (29). Following the HbS polymerisation, the RBCs containing them will assume sickle shape, become fragile and abnormal and are consequently destroyed by the immune cells (30,31). Subsequently, haemolytic anaemia sets in to provoke sickle cell crisis. On the other hand, plasmodial infection of AS-RBCs is not as serious. Only HbS is polymerised on infection leaving HbA intact. This event could also stimulate the immune cells to destroy the RBC containing the HbS and the plasmodial parasites. The parasitised HbA-RBC survive. This fact makes dominant homozygotes more susceptible to malaria than sickle heterozygotes. Although the high incidence of HbC gene (6%) was reported among the Yorubas of the Western States of Nigeria, (6), the rarity of the gene among the Igbos of Eastern Nigeria (10,12) was confirmed by this study from its low ultimate equilibrium percentage gene frequency of 0.07%. This rarity is suspected to be responsible for the very low incidences of HbCC (0.01%) and HbSC (0.02%) diseases observed in this study. It is not yet understood why the incidence of HbC gene is highest in Northern Ghana and Upper Volta (20%), higher in the Western States (6%) than Eastern States of Nigeria where the gene was formerly thought to be non-existence (8). However, one may implicate very recent intertribal marriages between the Yorubas and the Igbos or between Nigerians and Ghanaian which were formerly rare but intensified by urbanisation. Intertribal marriage could give rise to gene mixing. One may also suspect easier mutation on a β-globin gene that could result to haemoglobin C in Upper Volta, Ghana or the WesternStates, than in the Eastern States of Nigeria. The three-haemoglobin-band pattern reported in this study could be similar to those earlier reported by some workers (6, 12). But their identities were not known. It is proposed here that the two mutant haemoglobin bands may represent two different mutant haemoglobin entities other than HbS and HbC but with similar electrophoretic mobilities or the two haemoglobins may respectively be HbS and HbC. The results of the blood group antigen test (Table 2) confirmed the report that the incidence of blood antigen O predominate those of all other blood antigens while group AB exhibited the least incidence among Igbos of eastern States of Nigeria (4). The low incidence of Rhesus negative antigen observed in this population (2.97%) is a welcome discovery in view of the fact that low Rhesus negative incidence among females in the community could permit only very limited Rhesus-mismatched marriages among couples, which could generate less haemolytic disease of the newborn (erythroblastosis foetalis) associated with such marriages. In conclusion, this study has indeed not only revealed that SCA and malaria with common symptom of severe haemolytic anaemia, are highly prevalent in the communities of Old Aguata towns but also corroborated the high prevalence of malaria, SCA and its trait in the eastern State of Nigeria. The intense plasmodial infestation, attributable to drug-resistant strains of Plasmodium falciparum, probably the result of malarial drug abuse, was expected to reduce the incidence of SCA at the age range studied, but was found to the contrary. We attribute this to high incidence of sickle carriers, high persisting levels of HbF, identified in many of the sickle cell subjects in this population and some other genetic and environmental factors, for the high incidence of SCA revealed in this study. ACKNOWLEDGEMENT We are grateful to Dr. Simon Umego, Government GeneralHospital, Ekwulobia, Aguata local government area and Mr. Alphonsus Nneji, Microbiology Department of The University of Nigeria, Nsukka, for the Clinical and Parasitological diagnoses of malaria respectively. The study was partly supported with ICGEB, Trieste grant. No. 96/018. The wonderful assistance of Prof. A. Falarchi, Director, ICGEB, Trieste, Italy is hereby acknowledged. We are also very grateful to Prof. Michael O. Eze, Department of Chemistry, of the University of Winnipeg, Canada, for his assistance in arranging and securing the grant. REFERENCES
The following images related to this document are available:Photo images[bk03021t3.jpg] [bk03021t1.jpg] [bk03021t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}