 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
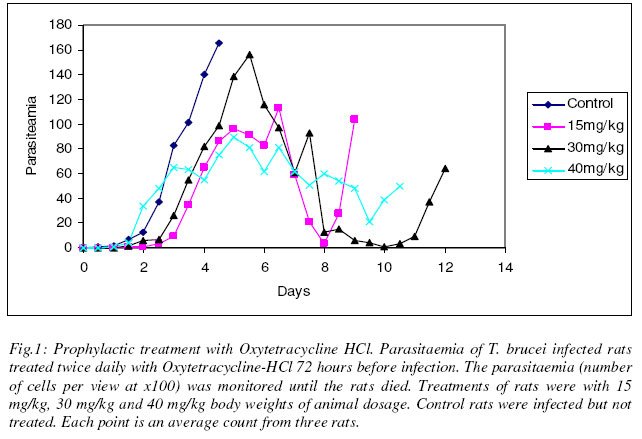
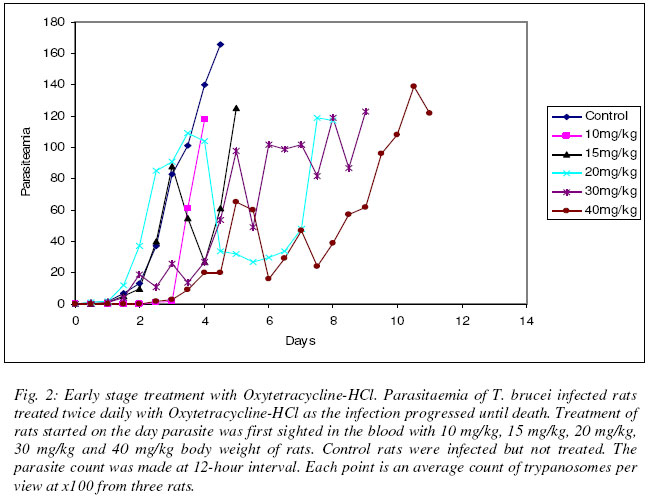
Justine T. EKANEM* and Iyabo S. ADENIRAN Department of Biochemistry, University of Ilorin, Ilorin, Nigeria Received 20 October 2003 Code Number: bk03022 Abstract We tested the antimicrobial drug Oxytetracycline-HCl against the infections of the protozoan parasite Trypanosoma brucei which causes the disease African Trypanosomosis. Oxytetracycline-HCl was used to treat rats infected with the disease at early and late stages of infections. Prophylactic administration was also carried out. The results showed that prophylactic treatment only did not inhibit the proliferation of the parasite but with continued administration kept parasitaemia low. Both prophylactic and early stage treatment of the infection extended the lifespan of the animals. Oxytetracycline-HCl was ineffective for the treatment of the late stage of infection. Oxytetracycline with its iron chelating property could be a useful inhibitor of ribonucleotide reductase, a cell cycle regulated enzyme that in turn regulates cell proliferation. Our experiment suggests that oxytetracycline-HCl could be a good management therapy for the African trypanosomosis if administered at the early stage of infection. Key words: oxytetracycline, sleeping sickness, management INTRODUCTION African trypanosomosis, the human sleeping sickness is a great threat to more than 60 million people in 36 countries of sub-Saharan Africa (WHO, 1998). The disease is caused by Trypanosoma brucei, a protozoan parasite transmitted from host to host by the blood sucking tsetse fly (Seebeck et al., 1999). Trypanosomes are a group of unicellular haemoflagellates of the genus Trypanosoma (WHO, 1986). The parasite lives and multiply in the host were it is able to establish its population through binary cell division (Vickerman, 1985; Ekanem, 2001). During the course of the infection there is a constant rising and falling parasitaemia resulting from the generation of sub-populations with antigenically different variant-specific glycoproteins (VSGs) at the cell surface (Rudenko et al., 1990). This property helps trypanosomes to evade the immune system, shield other proteins e.g. permeases on the surface from immune attack and inhibit phagocytosis (Rudenko et al., 1990). Due to this constant change in variable surface glycoproteins, termed antigenic variation, it has become quite difficult to develop vaccines against the disease trypanosomosis (Gray and Luckins, 1978; WHO, 1998). Most of the very limited drugs currently in use to combat it are either too expensive or does not satisfy even the minimal standards set today for drug safety and efficiency (Seebeck et al., 1999). Iron is essential to replication and growth (Romeo et al., 2001) thus all organisms have an ambiguous relationship with it (Hill, 1983; Halliwell and Gutteridge, 1990), a relationship pathogens and their hosts also share (Cabantchik et al., 1999). This dependence of pathogens on their hosts’ supply of iron makes them susceptible to the potential bacteriostatic effects of host iron limitation or chelation (Romeo et al., 2001). Much evidence abound to indicate that manipulation of the host iron status does lead to modulation of the proliferation and virulence of bacteria and protozoa (Richardson and Ponka, 1997; Mabeza et al., 1999). The essentiality of iron cannot be overlooked due to its role as prosthetic group in a variety of enzymes, electron transport proteins required for energy metabolism, for the biosynthesis of deoxyribonucleotides required for DNA synthesis and repair (Aisen et al., 1999), cell viability and proliferation (Richardson and Ponka, 1997). It is important in the G1 phase of the cell cycle and having its role in DNA synthesis liked to the activity of ribonucleotide reductase (RNR), an iron-reqiuring enzyme responsible for the synthesis of deoxyribonucleotides (Thelander and Reichard, 1979; Nuffer and Cheng 1987; Reichard, 1993; Jordan and Reichard, 1998). Ribonucleotide reductase plays a central role in DNA replication, cell division and proliferation by providing the deoxyribonucleotide (dNTP) precursors required (Nocentini, 1996; Ingram and Kinnaird, 1999). It is also the only enzyme along the cell division cycle that requires iron not as part of a prosthetic group but as part of the free iron pool (Thelander et al., 1983; Cooper et al., 1996). The role of iron in RNR activity has been elucidated through various studies by iron chelators to alter cellular iron status (Hoyes et al., 1992, Nyholm et al., 1993; Cooper et al., 1996) thus confirming the link between RNR activity and iron transport into proliferating cells (Chitamber et al., 2000). The essentiality of iron in RNR has been proved by studies which showed that when iron delivery is limited, the tyrosyl radical signal diminishes and the R2 subunit exists as a functionally inactive apo-R2 protein ( Nyholm et al., 1993). The structure of Ribonucleotide reductase has been described and classed into three different classes I (a and b), II and III (Chabes et al., 2000). Ribonucleotide reductase has been reported present in T. brucei (Hofer et al., 1997; 1998). Trypanosome RNR has binding site for iron (Hofer et al., 1997, Dormeyer et al., 1997). Unlike mammalian cells, bloodstream forms of T. brucei have a low iron content (Schell et al., 1991) which is attributed to its lack of cytochromes and like all living organisms trypanosomes require iron for growth (Schell et al., 1991; Steverding, 1998). The delivery of iron into bloodstream forms of trypanosome is mediated by host transferrin that is taken up via a unique receptor (Fast et al., 1999). The growth of trypanosomes has been successfully inhibited by iron chelation (Breidbach et al., 2002). The enzyme has also been reported as a possible target for drug discovery against sleeping sickness (Ekanem, 2001). Oxytetracycline belongs to a class of antibiotics of naturally occurring tetracyclines and has found much extensive use in combating both plant (McManus et al., 2002) and animal infections (Lents et al., 2002). The tetracyclines, oxytetracycline inclusive have been found to have strong iron chelating properties (Grenier et al., 2000). Oxytetracycline-HCl could be a possible drug against sleeping sickness more so when a related drug tetracycline-HCl has been reported as a possible cheap drug against the disease, African trypanosomosis (Ekanem et al., 2002; Johnson and Ekanem, 2003). In this report we have investigated the effect of oxytetracycline-HCl on T. brucei infected rats at early and late stages of infection. We have also assessed the effect of prophylactic treatment of the drug against the disease. MATERIALS AND METHODS Parasite Federe strain of African trypanosomes, Trypanosoma brucei was obtained from the Veterinary and Livestock Studies Department, Nigerian Institute for Trypanosoma Research (VLS – NITR), Vom near Jos in Plateau State of Nigeria. The parasite was injected into the rats intraperitoneally and maintained through repeated passages into other rats. Experimental animalsAlbino rats (Rattus novergicus species) of an average weight of 230g were used for the experiment. They were obtained from the Animal house of the Department of Biochemistry, University of Ilorin, Ilorin. The animals were kept under standard laboratory conditions and they were maintained on growers mash obtained from Bendel Feed and Flour Mills Ltd., Km 100 Benin-Auchi Road, Ewu, Edo State, Nigeria. ReagentsResearch grade reagents were used for the experiments. NaCl was a product of BDH Chemical Limited, Poole, England. Injectable form of Oxytetracycline-HCl 50mg/ml was a product of Hamexmedica Limited, Teriot Street, London, U.K. Administration of oxytetracycline-HClOxytetracycline-HCl was injected intraperitoneally into the animals twice daily at various concentrations taking the weights of the rats into consideration. Inoculation of parasitesThe tail of an infected rat was cleaned with a damp cloth and the tip of the tail was cut with a clean pair of scissors. Then the blood was extruded into 0.5ml normal saline solution. The extruded blood and saline solution was swirled to mix and then drawn into a 1ml syringe. A drop of the solution was placed on a microscope slide and observed under the light microscope to ascertain the presence of the parasite in the solution. Inoculation into the peritoneal cavity of an uninfected rat was carried out when parasite suspension contained 3 or 4 trypanosomes per view at x100 magnification. Parasitaemia (parasite count)Parasitaemia was determined by counting the number of trypanosomes per view under light microscope at x100 magnification from a thin blood smear obtained from the tip of the tail of an infected rat. The parasitaemia was recorded at 12 hr interval until the rats died. Prophylactic treatmentThree groups of three rats were treated with the drug for 72 hours at 12 hr interval before trypanosome infection and as the infection progresses until the death of the animals. The dosages used were 15mg/kg, 30mg/kg and 40mg/kg body weight of the rats. Early stage treatmentFive groups of three rats were treated with 10mg/kg, 15mg/kg, 20mg/kg, 30mg/kg and 40mg/kg body weights of the rats from the time the parasite was first sighted in the blood until the death of the rats. Late stage treatmentThree groups of three rats were used and treated with 10mg/kg, 20mg/kg and 40mg/kg body weights of the rats 48 hours after the parasite was first sighted in the blood until the animals died. ControlFor each experiment control rats (n=3) were injected with the parasite and allowed to develop without treatment. The parasitaemia was recorded until the death of the animals. Another group of three rats was also treated with oxytetracycline-HCl at 40mg/kg body weight of animals for 14 days without infection. RESULT Prophylactic treatment Fig. 1 shows the result obtained when rats were treated with oxytetracycline-HCl 72 hours before infection. At 15mg/kg body weight, the life span of the animals were increased beyond day 5 at which the control animals died to day 10 and the animals lived longer than control by 5 days. The parasitaemia increased steady from the day of infection and peaked the morning of day 7 after which it reduced to 3 parasites per view on day 8 before the death of the animals on day 10 with increased parasitaemia as indicated by the count for day 9. At double the dosage i.e. 30mg/kg, the parasitaemia increased steadily from the day of infection and reached its peak by the morning of day 6 then it dropped until one parasite per view was observed on day 10. The rats died on the morning of day 13 compared with the control animals, their life span were extended by 8 days. With a dosage of 40mg/kg, the parasitaemia gave steady increase and peaked on day 5. The animals died on day 11 having lived 6 days longer than the control animals. The parasitaemia was relatively low throughout the duration of infection when compared with the control. Early stageThe results obtained when the infected rats were treated with oxytetracycline-HCl immediately parasite was sighted are shown in Fig. 2. At 10mg/kg, the parasitaemia increased steadily and the animals died on day 4. At 15mg/kg there was a steady increase in parasitaemia till it peaked on day 5 when the rats eventually died. Treatment at 20mg/kg showed steady increase in parasitaemia until day 4 which was followed by reduction till day 7 when the parasitaemia increased and it reached its peak before the death of the rats on day 8. The lifespan of the animals were thus extended by 3 days compared with the control animals. At 30mg/kg, the lifespan of the rats were increased by 4 days. The parasitaemia increased steadily and reached its peak on day 9 after which death occurred. At 40mg/kg dosage, the parasitaemia increased steadily and reached a peak on day 11. The animals died on day 12 having had their lifespan extended by 7 days compared with the control. Late stageFig. 3 shows the result of treatment with oxytetracycline-HCl 48 hours after parasite was first sighted in the blood. At 10mg/kg, the parasitaemia increased steadily and peaked on day 3. The rats died on day 5. At 20mg/kg dosage, the parasitaemia increased steadily and reached its peak by day 3 when death occurred. At double the dosage i.e. 40mg/kg dosage, the parasitaemia increased steadily till it peaked on day 5 when the animals died. There was no extension in the lifespan of all the animals used in the late stage treatment compared with the control. Oxytetracycline-HCl treatment of uninfected rats Uninfected rats that were treated with oxytetracycline-HCl for 14 days showed no abnormality and no considerable loss in weight at the end of the experiment. None of the experimental rats survived beyond 14 days. DISCUSSIONWhen trypanosome enters into a system, the parasite proliferates to establish its population in the infected host through binary cell division (Vickerman, 1985; Ekanem, 2001). They release their toxins into the mammalian system (Ekanem et al., 1996a,b) and eventually kill their host. Iron plays a pivotal role in the synthesis of DNA in the trypanosomal system through the enzyme ribonucleotide reductase (Dormeyer et al., 1997; Hofer et al., 1997). The deprivation of a biological system of iron brings about the inactivation of the ribonucleotide reductase enzyme (Cooper et al., 1996; Chabes et al., 2000). It has also been established previously that the deprivation of iron inhibits the growth of cells, even those of trypanosomes (Fast et al., 1999). Iron chelation has thus been adopted as an attractive and possible strategy for new antiparasite drugs and treatment of sleeping sickness (Ekanem, 2001; Breidbach et al., 2002). Grenier et al. (2000) reported iron chelating properties of oxytetracycline. Our study revealed that early stage treatment and prophylactics at high dosage of 30mg/kg and 40mg/kg extended the life span of the animals. In the case of the prophylactic treatment very low parasitaemia with almost total clearance of parasite in the system was observed. This is probably as a result of the drug chelating iron present within the system since it had been introduced into the host system long before parasite invasion of the system unlike during the early stage treatment whereby the drug was not administered until the parasite was sighted present in the blood. Also, results obtained showed that at high dosage of 30mg/kg and 40mg/kg body weight the parasitaemia was less than those recorded with the lower dosage of 10mg/kg, 15mg/kg and 20 mg/kg although the administration of 15 mg/kg dosage for prophylactic treatment brought about death a day before the animals treated with 30 mg/kg body weight dosage died (Figs. 1 & 2). This suggests that high dosage of oxytetracycline is considerably effective for removal of the parasite from the system. Oxytetracycline seems to have been recognized as a potential trypanocide or combined therapy trypanocide judging by the interest it aroused in some non-professionals who exploited it to combat the disease trypanosomosis. Some Maasai pastoralists in Kenya administered oxytetracycline with other trypanocides as soon as the disease was recognized (Rodericks et al., 2000) but not as prophylactics. Our study established that the use of oxytetracycline-HCl for prophylactics kept parasitaemia low and considerably extended the lifespan of infected animals. Severity of the disease correlates with the level of parasitaemia (Murray et al., 1988). The late stage of trypanosomosis is usually characterized by high parasitaemia, reduced packed cell volume, and pronounce haemolytic anemia (Murray et al., 1988) due to the lysing of the erythrocyte of the animals. The late stage treatment by oxytetracycline-HCl was not effective since death resulted at about the same time as the control (Fig. 3). Obviously, the toxic substances released by the parasites into the blood stream (Ekanem, 1989; Ekanem et al., 1986a,b) completely overwhelms the hosts’ system ultimately resulting in the death of the animals as the disease takes its toll on the animals. We suggest that oxytetracycline-HCl could be a useful drug in the management of African sleeping sickness since it extends the lifespan of infected rats when treated at early stage. REFERENCES
The following images related to this document are available:Photo images[bk03022f1.jpg] [bk03022f2.jpg] [bk03022f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}