 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biokemistri, Vol. 16, No.1, June, 2004, pp.49-55
Assessment of biological value of Terminalia catappa seed meal-based diet in rats Nasir O. MUHAMMAD* and Oyelola B. OLOYEDE Department of Biochemistry, University of Ilorin, Ilorin, Nigeria.
Received 23 November 2003 Code Number: bk04008 Abstract The effects of defatted Terminalia catappa seed meal on the growth performance and carcass of rats were investigated. Twelve weaning albino rats (Rattus norvegicus) with an average weight of 24.0 ± 3.4g were maintained on diets composed of defatted T. catappa seed meal (Tc meal) and soybean meal (control) for six weeks. The weights of the rats were monitored on weekly basis, at the same period of the day and before being served the (weighing) day’s feed. The organs and carcasses of the rats were weighed after they were sacrificed and disemboweled, and the chemical compositions of the carcasses were also determined. The body weights of the rats maintained on defatted Tc meal were significantly reduced to one-third (1/3) of the weights of the control animal (p<0.05). The organ to body weight ratio of the Tc meal rats were significantly higher (p<0.05) than that of the control. In the carcass of rats fed Tc meal diet, ether extract, crude protein and ash contents were significantly reduced (p<0.05) when compared with the control. However, the crude fibre and the nitrogen free extract (NFE) were significantly higher in the Tc meal rats than that of the control (p<0.05). It can be deduced that the defatted Terminalia catappa seed meal could cause depression in growth rate, enlargement of rat tissues with adverse effects on carcass of rats. Key words: Terminalia catappa, defatted meal, growth depression, carcasses INTRODUCTION
Of all the food nutrients, protein requirements are fundamental, without which life would be impossible.1 Soybean has been found to be superior to other plant protein supplements.2 Soybean is however, presently not sufficiently produced to meet the demand of its use as plant protein ingredient in both livestock and human feeds. This limitation coupled with the high cost of animal source of protein (which are of course of better quality than plant sources) poses a challenge for a search for alternative source of plant protein for human and animal consumption. Furthermore, the lack of adequate information on the chemical and biological values of many plant materials of potential value as human and animal food has limited utilization of such materials, particularly protein and oil rich seeds; such should therefore be exploited. Among such alternatives is the seed of Terminalia catappa. Terminalia catappa is a medium size tree whose branches form layers of canopy. The leaves of this tree have been reported to have medicinal values3,4. The fruit is edible, fleshy, green (unripe) and yellow or red (when ripe). The exocarp is relatively thin and smooth, and the endocarp is hard. When cracked, a kernel is obtained and this can be consumed as well. The kernel of T. catappa has aphrodisiac activity and may be useful in the treatment of certain form of sexual inadequacies such as premature ejaculation.5 The seed is very rich in protein (19 – 22%) and oil (50 – 52 %).6 The amino acid and mineral profiles of this seed has equally been documented.7 Information on the biological value of T. catappa seed consumption is still very scanty in literature. This study therefore aimed at investigating its effect on the growth performance and carcass of rats. MATERIALS AND METHODS Materials The ripe fruits of Terminalia catappa were picked from the premises of the University of Ilorin, Ilorin, Nigeria and authenticated at the Deparment of Plant Biology of the University. The albino rats were inbred in the Department of Biochemistry, University of Ilorin, Ilorin, Nigeria. Processing of Seed mealThe riped fruits were oven-dried at 60oC and then cracked to remove the seeds, using 125mm Bench vice, FUKUNG Brand made in The People’s Republic of China. The seeds were then milled using a manual grinding machine and the resulting meal was subsequently defatted with Soxleht extractor using petroleum ether (40oC – 60oC) as the extracting solvent. The extracted meal was autoclaved at 121oC and 15kgcm2 for 30 minutes and was subsequently used in compounding rat feed using defatted soybean meal as control8. The formulated diets are shown in Table 1. Table 1: Composition of Diet
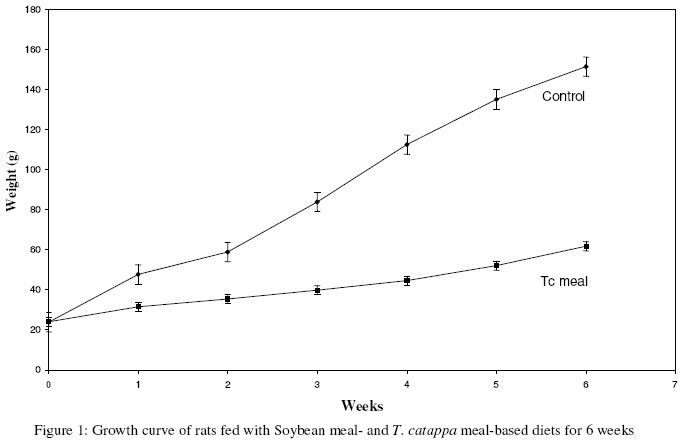
* Soybean meal-based diet The proximate analysis of the composed diet was carried out using the method described by AOAC.9 Animals and DietsTwelve male, 3-weeks old weaning albino rats of average weight 24.0 ± 3.4g were divided into two groups of six (6) animals each. The rats were maintained on normal rat chow, allowed to acclimatized for a week, fasted for 24 hours, and then placed on the different diets. The first group of rats (the control) was placed on the defatted soybean meal-based diet while the second group of rats (Tc meal) was placed on defatted Terminalia catappa seed meal-based diet. The rats were housed in 33cm x 20.5cm x 19cm plastic metabolic cages and were fed with their respective diets and water ad libitum for six weeks at 25oC ± 1oC, 50% ± 5% humidity and, 12 hours light and 12 hours dark conditions. They were weighed weekly and at the end of the experimental feeding period, were sacrificed by anaesthetizing with (cotton wool soaked in) chloroform. They were then quickly dissected to excise the brain, liver, kidney, stomach and small intestine. The kidney was decapsulated and the stomach and intestine cleaned free of food. The organs and the carcasses were then weighed. The carcasses were dried in an oven at 80oC, and then analysed using the method of AOAC9. Statistical analysis was carried out using the students’ t-test.10 RESULTSTable 2 shows the proximate analysis of the formulated diets (test and control) on which the rats were maintained. There was no significant difference (p>0.05) in the various components of the diets. Rats maintained on the Tc meal-based diet had growth depression as shown by the body weights (Table 3) and growth pattern (Figure 1). The weights of rats fed with Tc meal were significantly reduced (p<0.05) to about one-third (1/3) of the control. The carcass weights of the experimental animals are shown in Table 3. The weights of the various organs and organ:body weight ratio of the rats are shown in Table 4. The weights of the organs of rats maintained on the Tc meal were significantly lower (p<0.05) than those of the control animals, however, there were significant increases (p<0.05) in the organ:body weight ratio of rats reared on the Tc meal diet compared to the control. The proximate composition of the carcasses of rats in both group are shown in Table 5. When compared with the control, the crude protein, ether extract and ash contents of the carcass of rats fed with the Tc meal diet were significantly reduced (p<0.05) while there were significant increases (p<0.05) in the crude fibre and nitrogen free extract (NFE) of the Tc meal rats. Table 2: Proximate Composition (%) of the Formulated Diets
Values are expressed as mean of three determinations ± SEM. Values are not significantly different (p>0.05) Table 3: Body and Carcass weights (g) of rats maintained on soybean meal-based- and T. catappa seed meal-based diets for six weeks
Values are expressed as mean of six determinations ± SEM. The values obtained from proximate analysis of the formulated diets (Table 2) compared favourably with each other, indicating that the rats in both groups were placed on the same amount of nutrients quantitatively and therefore whatever differences that are noticed might be due to the differences in the quality of the nutrients in these diets or their bioavailability when consumed. The growth rate depression (Fig. 1) and significant body weight reduction (Table 3) in the rats placed on the Tc meal diet is an Table 4: Weight of organs and Organ:Body weight ratio of rats maintained on soybean meal-based- and T. catappa seed meal-based diets for six weeks.
Values are expressed as mean of six determinations ± SEM. Table 5: Proximate Composition (% based on dry weight) of Carcass of rats maintained on soybean meal-based and T. catappa seed meal-based diets for six weeks.
Values are expressed as mean of six determinations ± SEM. indication that the defatted Tc meal does not support the growth of the animals. This is probably due to the high phytate, oxalate and tannin contents of the seed, which might have rendered unavailable the protein and mineral content of the seed.7 Phytate, an antinutrient, is a strong chelator of many divalent minerals such as Cu, Ca, Mg, Zn and Fe. The ability of phytate to chelate these essential minerals has been recognised as a potential threat in animal and human nutrition.11,12 Oxalate has also been reported to chelate with Ca2+ forming insoluble calcium oxalate.7 One of the nutritional effects attributed to tannins is interference with the absorption of calcium (Ca2+).13 Animals deficient in some minerals like Mg2+, Se2+ and Zn2+ have been documented to have impaired nucleic acid and protein synthesis.14,15 The deficiency or unavailability of these minerals would therefore lead to impairment in growth and development of the animals as observed in the present study. Additionally, phytates react directly with charged groups of proteins mediated by a positively charged mineral ion such as Ca2+ , due to their ionic nature, thus the resultant phytate-protein and phytate-mineral-protein complexes may also adversely influence protein digestion and bioavailability.16 Tannins have also been reported to decrease protein quality by decreasing digestibility and palatability.13 It is possible that these antinutritional factors inhibit trypsin and pepsin with their protein complexing ability.16 Trypsin inhibitors have been known to interfere with the physiological process of digestion through interference with the normal functioning of the pancreatic proteolytic enzymes in non-ruminants,17 leading to severe growth depression. It is possible that these antinutrients have interacted directly with the proteolytic enzymes leading to a corresponding reduction in the digestibility of the proteins in the diet.18, 19 Two of the animals on the Tc meal-based diet died before the termination of the experiment and this might be due to the possible toxicity of the Tc meal on the internal organs of the rats as shown by the signficant increase in the organ to body weight ratio on the rats maintained on the meal (Table 4). It has been reported that diets with high oxalate content could increase the risk of renal calcium absorption.20, 21 Tannins have also been reported to cause damage to the intestinal tract, toxic when absorbed from the gut and exhibit a possible carcinogenic effect.13 The results from the carcass evaluation of the rats on the Tc meal diet (Table 5) further corroborated the evidence that the high protein quality, inherent in the seed of Terminalia catappa had not been utilised to form body muscle as a result of possible chelation by the antinutrients. The significant reduction in ether extract of the carcass of rats placed on Tc meal diet (Table 5) further indicated that the lipid metabolism in these animals might have been adversely affected. This is because alteration in protein and lipid metabolism during deficiency of some minerals like Zn2+ and Mg2+ have been reported in literature.14, 15, 22, 23, 24 It may therefore be argued that reduction in the bioavailability of the mineral and protein contents of the Tc meal (due to its antinutritional factors), has led to the observed significant decrease in weight and depression in growth rate of the animals fed with the Tc meal-based diet. Furthermore, some of these antinutrients, especially phytates insoluble oxalates and condensed tannins, are heat stable, thus could withstand heat, harsh field conditions, seed/grain transportation, and storage environment, coupled with the fact that food processing alone may not prevent their potential adverse effects as some of them typically remained in the processed food.25, 26 Perhaps, the use of exogenous enzymes for phytate reduction in monogastric nutrition which is recently suggested for the development of thermostable phytases 27 or the use of fungi treatment to reduce antinutrients in foods, 28 would assist in the usage of this protein-rich seed in animal and human nutrition. REFERENCES
© 2004 Nigerian Society for Experimental Biology. The following images related to this document are available:Photo images[bk04008f1.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}