 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
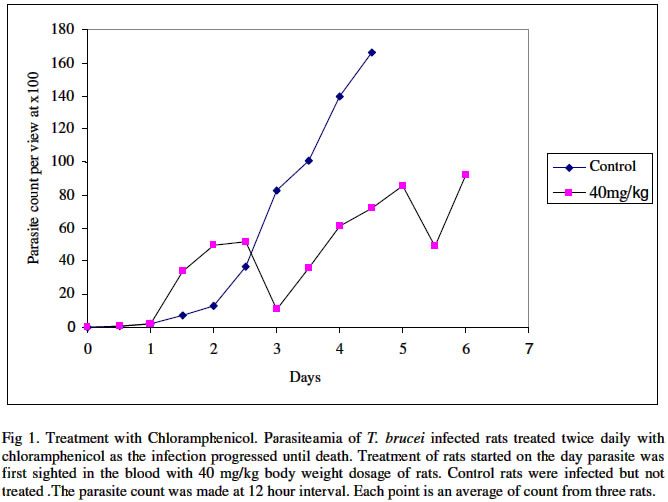
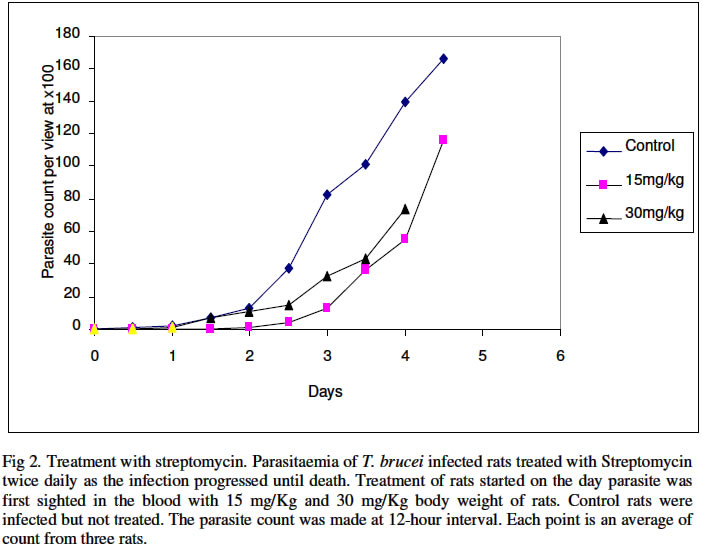
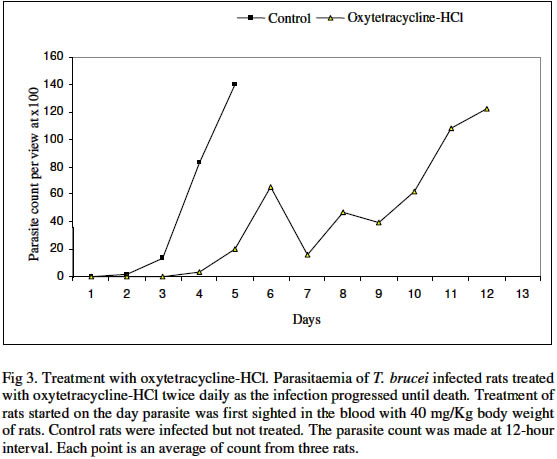
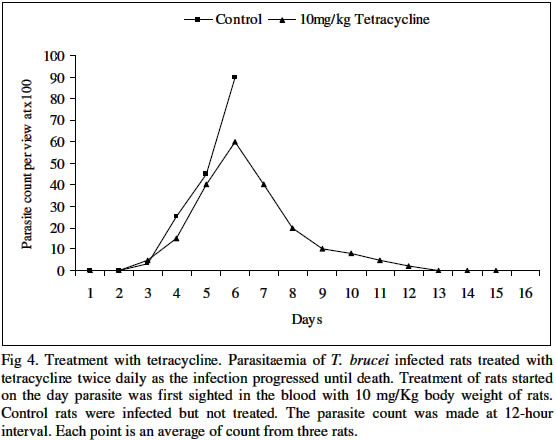
Biokemistri, Vol. 16, No. 2, December, 2004, pp. 56-63 Iron chelation excludes protein synthesis inhibition in the tetracycline management of African trypanosomosis Justine T. EKANEM*, Titilayo O. JOHNSON, Iyabo S. ADENIRAN and Valeelat OKEOLA Department of Biochemistry, University of Ilorin, Ilorin, Nigeria. Received 12 December 2003 Code Number: bk04020 ABSTRACT Ribonucleotide reductase, an iron requiring enzyme necessary in the production of deoxyribonucleotides required for replication in cell division and proliferation is induced during the S phase of the cell cycle. We have compared the trypanocidal properties of four antibiotics that show bactericidal activities by destabilizing ribosome-mRNA complex to inhibit protein synthesis. Tetracycline and oxytetracycline that have iron chelating properties extended the lifespan of trypanosome infected rats from 6 and 5 days of control to 15 and 12 days respectively while chloramphenicol and streptomycin that have no iron chelating properties could not extend the lifespan of infected rats. We confirm our earlier report that iron chelation plays a prominent role in the tetracycline management of African trypanosomosis. Key words: Tetracycline, iron chelation, T. brucei, growth inhibition INTRODUCTION Trypanosoma brucei, the causative agent of African trypanosomosis divides and proliferates by binary fission in the process of establishing an infection in the host. Parasitaemia has been shown to correlate with the severity of the disease1. A major requirement of rapidly dividing cells including trypanosomes is the intracellular availability of unbound iron required by ribonucleotide reductase2,3 in the synthesis of deoxyribonucleotides (dNTPs) from ribonucleotides (NTPs). RNR is responsible for the production of all the four dNTPs required for replication, a necessary step in cell division and proliferation4-7. Iron chelation inhibits ribonucleotide reductase8-10. The enzyme has been suggested as an enzyme of promise for controlling African trypanosomosis11. Ribonucleotide reductase in itself is a cell cycle dependent enzyme12. Its activity peaks at the synthetic (S) phase of the cell cycle and falls at other phases. The protein of the enzyme is synthesized from its gene by induction during the S phase12. Many antibiotics with bactericidal properties exert their effects by inhibiting the induction of proteins or specific enzymes from their genes13. We have earlier reported that tetracycline14,15 and oxytetracycline16 can be useful in the clinical management of African trypanosomosis based on earlier reports that these antibiotics have iron chelating properties11. They are however well documented protein synthesis inhibitors that destabilize ribosome-mRNA complex during protein synthesis13,18. Other antibiotics that also destabilize the complex are chloramphenicol and streptomycin13,18. They however do not have iron chelating properties18. In this report we have investigated the possibility of protein synthesis inhibition contributing to the possible management of African trypanosomosis by tetracycline. MATERIALS AND METHODS Federe strain of African trypanosomes, Trypanosoma brucei, was obtained from the Veterinary and Livestock Studies Department, Nigeria Institute for Trypanosomiasis Research (VLS – NITR), Vom near Jos in PlateauState of Nigeria. The parasite was injected into the rats intraperitoneally and maintained by repeated passages into other rats. Antimicrobial drugsFour antimicrobial drugs were used to treat the experimental rats after infection. Injectable form of oxytetracycline HCl was a product of Haemexmedical Limited, Teviot Street, London, U.K, streptomycin sulphate B.P was a product of RIKA Pharma Gmbh, Hamburg, Germany and chloramphenicol B.P was a product of Greenfield International Ltd. London- England. Tetracycline was a product of Sigma Chemical Company, England. Inoculation of parasiteThe tail of an infected rat was cleaned with a damp cloth and tip of the tail was cut with a clean pair of scissors. Then the blood was extruded into about 0.5ml normal saline. The extruded blood and saline solution was swirled to mix and drawn into a 1ml syringe. A drop of the solution was placed on a microscope slide and observed under the light microscope to ascertain the presence of the parasite in the solution. Inoculation into the peritoneal cavity of an uninfected rat was carried out when parasite suspension contained 3 or 4 trypanosomes per view at x100 magnification. Parasite count Parasitaemia was determined by counting the number of trypanosomes per view under the light microscope at x100 magnification from a thin blood smear obtained from the tip of the tail of an infected rat. The Parasitaemia was recorded at 12 hour interval until the rats died. Administration of drugsTetracycline, oxytetracycline-HCl, streptomycin and chloramphenicol were all injected intraperitoneally into the animals twice daily at various concentrations as recommended by manufacturers taking the body weights of the rats into consideration. The administration started on the first day of sighting parasite in the blood and was continued until the rats died. Tetracycline: 50 mg of tetracycline mixed with a small quantity of water was made up to 100ml. Volumes corresponding to 10 mg/Kg rat weight were administered to the rats. Chloramphenicol: 500mg of streptomycin was dissolved in 10ml of distilled water. Volumes corresponding to 40mg/kg body weights were administered to the rats. Streptomycin: 5mg of streptomycin was dissolved in 10ml of distilled water. Volumes corresponding to 15mg/kg and 30mg/kg body weight of the animals were administered to the rats. Oxytetracycline-HCl: The injectable solution was obtained. The solution contained 50mg of oxytetracycline-HCl per ml. Volumes corresponding to 40mg/Kg body weights were administered to the rats. Control: Control rats (n=3) were infected with the parasite Trypanosoma brucei and allowed to develop without treatment. The parasitaemia was recorded until the death of the animals. Another group of three uninfected rats was also treated with the antimicrobial drugs for 21 days without infection to ascertain survival of treated animal in presence of the drugs. Treatment with the Drugs The parasitaemia of untreated rats increased infinitely and the rats died on day 5. The uninfected rats survived the treatments for 21 days. The treatment of Trypanosoma brucei infected rats immediately parasite was first sighted in the blood with chloramphenicol is shown in Fig 1. Chloramphenicol when administered at 40mg/kg body weight did not extend the life span of the infected rats considerably more than the control group since the animals died on day 6 which is a day after death was recorded for the control group. A lower parasitaemia was however observed during the duration of the infection before day 6 that the animals died. The results obtained for the treatment of infected rats when parasite was first sighted in the rats using streptomycin are shown in Fig. 2. At 15mg/kg dosage, steady increase in parasitaemia was recorded and death ultimately on day 5 as in the control. At double the dosage, 30 mg/Kg body weight, there was steady increase in parasitaemia until day 4 when the rats died. Thus, there was no extension in the life span of the animals beyond that of the control. Parasitaemia was generally lower than those of the control in both cases. Oxytetracycline-HCl extended the lifespan from 5 to 12 days as seen in Fig. 3 and tetracycline did not only extend the lifespan to 15 days but also cleared parasitaemia by the 13th day (Fig. 4). On invasion of a mammalian system, trypanosomes proliferate rapidly through binary cell division to establish its population in the infected host19. Toxins are released into the mammalian system by the trypanosomes20 and this eventually kills their host. Generally, trypanosomosis lowers the resistance of infected host to other diseases and worse still, it leads to sub-optimal immune responses after vaccination21,22. Iron plays a pivotal role in the synthesis of DNA in trypanosomal system through the enzyme ribonucleotide reductase23,24. If a biological system is starved of iron, the enzyme ribonucleotide reductase becomes inactivated25-27. Previous studies have revealed that growth of cells even those of trypanosomes can be successfully inhibited by depriving the cells of iron27. Consequently, iron-chelation has thus been adopted as an attractive and possible strategy for new anti-parasite drugs and treatment of sleeping sickness11,28. Ribonucleotide reductase is a cell cycle induced enzyme12 that is equally dependent on continuous availability of recyclable iron2. The antibiotics used in this study are all established protein synthesis inhibitors and in addition tetracycline and oxytetracycline are reported iron chelators17 that we have earlier reported as possible candidates for clinical management of African trypanosomosis14-15. Chloramphenicol and streptomycin do not chelate iron18. Chloramphenicol and streptomycin have been shown in this study to be ineffective against the protozoan parasite Trypanosoma brucei (Figs. 1 and 2). Their ineffectiveness could be attributed to their inability to chelate iron. In fact, chloramphenicol has an untoward effect of increasing serum iron29. Tetracycline and oxytetracycline notably extended the lifespan of T. brucei infected rats (Figs 3 and 4). The results suggest that protein synthesis inhibition contributes minimally to reduction in parasitaemia observed at the early stages of infection and do not play a part in the extension of the lifespan of infected rats as observed in the treatment with the tetracycline group. We confirm our earlier result that iron chelation plays a prominent role in the suggested tetracycline management of African sleeping sickness. REFERENCES
© 2004 Nigerian Society for Experimental Biology The following images related to this document are available:Photo images[bk04020f4.jpg] [bk04020f1.jpg] [bk04020f2.jpg] [bk04020f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}