 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biokemistri, Vol. 16, No. 2, December, 2004, pp. 72-78
Inhibitory activity of seed extract from Picralima nitida, (Staph) on β-D-glucosidase Enomfon J. AKPAN1 and Ime B. UMOH2 1. Department of Biochemistry, Faculty of Basic Medical
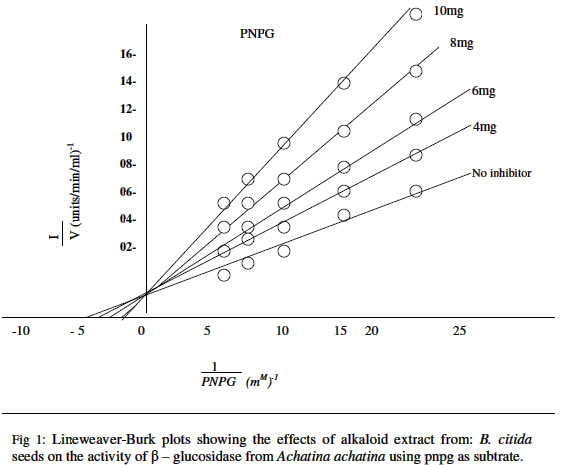
Sciences, University of Uyo, Nigeria. Received 15 October 2003 Code Number: bk04022 ABSTRACT The rate of hydrolysis of p- nitrophenyl β-D-glucopyranoside (PNPG) by β-D glucosidase (β-D-glucoside glucohydrolase, E.C. 2.1.21) from African giant snail (Achatina achatina) has been studied in the presence of seed extracts from Picralima nitida (Staph) Th&H.D. A at a substrate concentration range of 0.04-0.2mM there was no product (P- nitrophenol) inhibition of the enzyme. The Km and Vmax values for PNPG were calculated from Line Weaver-Burk plots and the values obtained were 0.214mM and 125-nmol/mg protein/min respectively. Β-glucosidase from Achatina achatina degraded PNPG faster than, that from such as Aspergillus niger Humicola grisea and Geotricum. spp The alkaloid extracts from the mature seeds of Picralima nitida inhibited the enzyme in a competitive manner with inhibitor equilibrium constant (ki) values of 0.263, 0.157.0.125 and 0.10mg/1 for alkaloid (inhibitor) concentrations of 4,6,8 and 10mg./l respectively. The biochemical implication of this study is that since the seeds extract showed inhibition of enzyme activity, the exploitation of this potential could be of immense value in the current search for new therapeutically effective drugs with inhibitory effects against β-glucosidae which is implicated in HIV infectivity of cells. Key words: β-D-glucosidase, P. nitida, inhibition, HIV infectivity INTRODUCTION Medicinal plants are known to contain drug bases such as alkaloids, saponins, tannins, cardiac glycosides, and polyphenols. More than 30% of commercial drugs are derived from active plant extracts (1). There are claims by herbalists in recent times that certain ailments e.g. cancer, malaria, AIDS, viral and some bacterial infections which have defied Western medicine can be cured with local herbs. In Nigeria, records indicate that about 4.5% of the people are infected with AIDS Virus (2). Drugs currently in use have cumbersome dosage regime unbearable and potentially fatal adverse effects, and are generally beyond the reach of the common man. Perhaps the most important treasure of the plant kingdom, which is awaiting discovery, is their medicinal components and their use in health and diseases. It has been reported that some of the world’s AIDS patients are being treated using traditional herbal medicines (3). β-D-glucosidase, a hydrolytic enzyme of the AIDS Virus is implicated in the amplification of this disease. Montefiori et al (4) have shown that the full pathogenic potential of HIV-in vitro is realized only if the viral envelope proteins are N- glycosylated and the carbohydrate moieties are subsequently processed correctly. The pathogenic mechanisms affected by protein glycosylation include virus infectivity and cytopathicity. Once HIV-1 envelope proteins are glycosylated, the carbohydrate moieties must be processed by action that includes the activities of glucosidase in order to elicit a full pathogenic response. In view of these findings inhibitors of glycoprotein processing that act at the level of lipid-linked oligosaccharide precursor synthesis, glucosidase1 and lipid-linked oligosaccharide precursor synthesis, glucosidase 1 activity and mannosidase 1 activity may have potentials as anti-HIV-1 therapeutic agents. The Apocynaceae plant picralima nitida (staph) th & H. Durand is widely but sparsely distributed in Ghana and other West African countries where the plant seeds are alleged to be used for various medicinal purposes. In Ghana the seed preparations are commonly used as an analgesic (5). Clinquart (6) first isolated akuammine the major alkaloid of the plant. Akuammicine akuammidine, akuammigine, pseudo-akuammigine, picraline and picralicine have also been isolated from the mature seed of P. nitida (7,9,10). Also isolated is a triterpenoidal saponin (β-amyrin) from the stem bark. The technical aspects of cultivating large numbers of organisms potentially pathogenic, pose formidable challenges to third world laboratories. This paper investigated an alternative “surrogate” model for assessing anti-infective herbs, seeds, and fractions using biochemical model key enzymes from the African giant snail (the surrogate) with kinetic parameters similar to enzymes from human immune deficiency virus, (HIV) β-D glucosidase. There is no previous report on the inhibitory effects of extracts of the seeds of P. nitida on β-D- glucosidase. This prompted the authors to probe for possible inhibitory effects of the alkaloids on the enzyme. MATERIALS AND METHODS OrganismA number of snails (Achatina achatina), bought from the local market, were maintained in the laboratory (22-25°C) in a perforated metal box. The animals were fed on fresh paw-paw (Carica papaya) leaves. The box was moistened with tap water at regular intervals. ChemicalsP-Nitropheny1 β-D-glucopyranoside was obtained from koch-light laboratories, colnbrook, Burk. U. K. All other chemicals were of ANALAR grade. Collection and treatment of seed sampleThe mature fruits of Picralima nitida were collected from a bush in EkparakwaVillage in Oruk anam L.G.A., AkwaIbomState of Nigeria between the months of March and May 2001. The fruits were thoroughly washed, dried and seeds removed and sun-dried for 5 days. The dried seeds were then pulverized using an electric blender (Mx-302N, Japan). The powder obtained was sieved (0.2mm) and stored in a tightly closed dark glass container in a refrigerator. Extraction of total alkaloidal content of the seed5kg of the powdered seed was placed in a soxhlet and exhaustively extracted using 5 liters of redistilled methanol. The methanol extract was evaporated in vacuo using a rotary evaporator and dried to a constant weight. The residue was taken up in 3% HCI and filtered. Freshly prepared Mayer’s reagent was added drop wise to precipitate the alkaloid. The precipitated alkaloid was filtered and air-dried and the residue redissolved in a mixture of acetone: methanol: water (6:2:1). It was then filtered and evaporated to dryness, using a tarred dish. The total alkaloidal content was 0.6g. Extraction and purification of β -glucosidaseThe digestive tracts of the snail were removed immediately after de-shelling and stored frozen at – 15°C. When required, the thawed digestive tracts were gently pounded in a pre-cooled mortar till a homogeneous pulp was obtained. This was taken up in a minimal amount of Mclvaine buffer, pH 5.0 containing 0.1M-NaCl (11). The gut contents were then extracted in a 100ml round-bottomed flask with agitation and later centrifuged at 10,000xg for 30min at 4°C. The supernatants were stored frozen at –15°C until required. β-Glucosidase from the snail was obtained from the wall (crop, stomach and intestine) first by washing the contents four times with 50ml of 0.05M sodium acetate buffer (pH 5.0). About 6-8g (fresh weight) of the various sections of the washed gut wall was then homogenized manually using mortar and pestle in a minimum amount of the acetate buffer, and the homogenates were then centrifuged at 10,000 x g for 15 min at 4°C. The supernatant solutions containing the crude enzyme were mixed with twice their volume of cold acetone and left at 4°C for 30min. The precipitates obtained after centrifugation at 10,000 x g for 15min were separately dissolved in 0.05M-soium acetate buffer (pH 5.0). After centrifugation to remove undisclosed particles, the supernatant solutions were made up to 8ml with the above buffer and portions of these solutions were used for enzyme assay. Umezurike (12) reported that β-Glucosidase is completely precipitated from solution in the presence of 2 volumes of acetone. Protein determinationProtein content of the enzyme extract was determined by the method of Lowry et al (14) with bovine serum albumin as standard. Enzyme assayβ-D-glucosidase activity was assayed with P-nitrophenyl β-D-glucopyranoside (substrate) in 0.05M-sodium acetate acetic acid buffer (pH 5.0) as described by Berfneld (15). β-D-glucosidase activity was expressed as nMol of P-nitrophenol liberated/min per mg fresh weight of tissue. Michaelis-menten contants (km) and maximum velocities (V max) were calculated from Line weaver Burk plots of 1/v against 1/[s]. While Ki values were obtained from Dixon plots of 1/v against [I] (16). Inhibition studiesThe stock alkaloid standard of 200mg/1000cm3 in methanol was prepared. The following concentrations 2,4,6,8,10,12 mg/l were prepared from the stock by serial dilutions. β-glucosidase activity was again determined after adding various concentration range used was 0.04-0.2mM PNPG. RESULTS AND DISCUSSION The seed of P. nitida were confirmed quantitatively to be rich in alkaloids. The alkaloid concentration was 0.6g/5kg used for extraction. The enzyme was partially purified using acetone (12). The concentrations of protein before and after acetone precipitation are reported in Table1.Line weaver-Burk plots (17) of the results obtained when velocity was measured as a function of substrate concentration showed that the enzyme was not inhibited by P-nitrophenol (product) at a substrate concentration range of 0.04 – 0.2mM P-nitrophenyl β-D-glucopyranoside (substrate). TABLE: 1 Protein Contents of β-D glucosidase extract before and after acetone precipitation.
The data obtained in the presence of 0.04 – 0.2mM PNPG are plotted as 1/v versus 1/[s] In Fig. 1. The Km value calculated from the graph in Fig. 1 was 0.214mM. The Vmax value for PNPG was 125 nmol/mg protein/min slope (Vmax/km) was 584. The Km value of 0.214mM for PNPG is however lower than 7.064mM reported for ortho-nitrophenyl-β-D glucopyranoside (ONPG) at a concentration range of 0.1-10.0m M (17) for β-glucosidase from Humicola grisea. It is also lower than 1.63mM (0.5-10.0mM) reported for cellobiose substrate (17). The lower apparent Michaelis-Menten constant (Km) indicates that the enzyme showed a higher affinity for the synthetic β-D-glucoside substrates than for the natural substrates such as cellobiose. This observation is consistent with a similar report by Dekker (19). The enzyme β-D-glucosidase is relatively unspecific, and its activity varies widely with different substrates such as salicin, cellobiose, etc. for example, if efficiency of substrate utilization is estimated on the basis of slope Vmax/Km ratio (20), the value of 584 obtained for PNPG indicates that this substrate is the best compared to 133.9 for PNPG and 26.1 for cellobiose (17). The Vmax value for PNPG was 125-nmol/mg protein/min using the substrate P-nitrophenyl β-D-glucopyranoside (PNPG). This value is however higher than 3.76-unit/mg protein/min for the substrate PNPG and 33.8 nmol/mg protein/min for cellobiose reported for β-D-glucosidase obtained from Aspergillus niger (19). Peralta et al (18)reported Vmax values of 26.1 and 33.96 units/mgprotein/min for cellobiose and ONPG as substrates for β-D-glucopyranoside from Humicola grisea. The result of this study indicates that β-D-glucopyranoside from Achatina achatina degrade PNPG faster than the same enzyme from other sources using other substrates. β-D-glucosidase activity was also assayed using various concentrations of the alkaloids. The results showed that the alkaloids inhibited the hydrolytic activity of the enzyme even at concentrations as low as 4mg. Alkaloids are reported to function at low concentrations (21,22). Line weaver-Burk plots showed that the inhibition was a competitive type and Ki values were 0.26mg/1 at inhibitor concentration of 4mg/1. Others were 0.17, 0.13 and 0.10mg/1 for inhibitor concentration of 6,8 and 10mg/1 respectively. Competitive inhibitions of β-glucosidase by glucose and other inhibitors have been reported (12,19,23) . The ability of P. nitida seed extract to inhibit the activity of β-glucosidase from Achatina achatina is a promising development in view of the implication of this enzyme in HIV-1 infectivity. The possibility of equivalence of enzyme from both sources (AIDS Virus and Surrogate) is enhanced by the broad nature of cellulases (both endo and exo glucanases) and glycosidase present in the cellulase- glucosidase complex (24). It is noteworthy that β-glucosidase with molecular weight of about 40,000D has been found in animal tissues (24). Since cellulose form a major source of carbohydrate for snails, it is not surprising that β-glucosidase which is always associated with cellulase, is found in large amounts in their gut contents. Β-glucosidase action appears to play a major role in providing the glucose used by this animal for metabolic processes (13). β-glucosidasee is also implicated in the processing of carbohydrate moieties during glucosylation by HIV-1 virus in order to elicit a full pathogenic response (4). Recent discoveries of active agents in plants (20) coupled with the results of this study have supported the idea that successive integration of traditional medicine into the health care delivery system particularly in the third world countries need be encouraged. However, the efficacy of such plant extracts must be scientifically established. A continuous search for substances active especially against AIDS virus with low toxicity would no doubt emerge in the near future. Further studies and applications of the seed extracts of P. nitida on human HIV may provide scientific basis for the use of these plant extracts as medicaments. REFERENCES
© 2004 Nigerian Society for Experimental Biology. The following images related to this document are available:Photo images[bk04022f1.jpg] |
| |||||||||

{kind=link}