 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biokemistri, Vol. 16, No. 2, December, 2004, pp. 112-120 Time-course analysis of the accumulation of phenols in tomato seedlings infected with Potato Virus Xand Tobacco mosaic virus Olusegun S. BALOGUN1 and Tohru TERAOKA2 1Department of Crop Protection, Faculty of Agriculture, University of Ilorin,
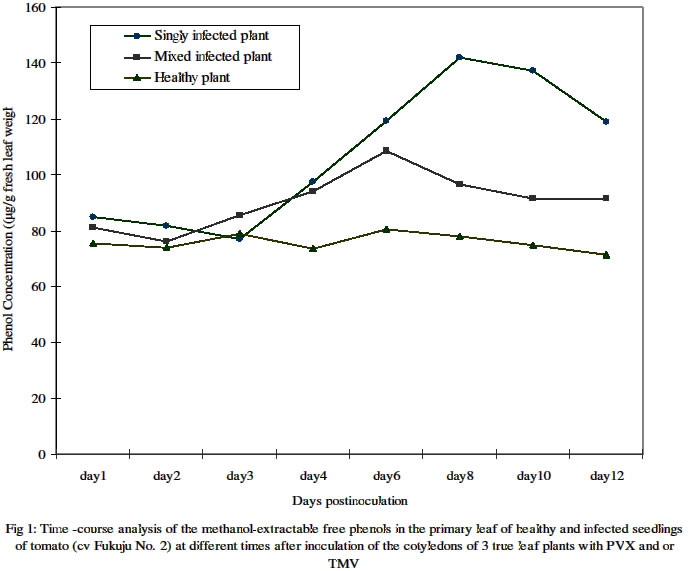
PMB 1515, Ilorin, Nigeria. Received 19 May 2004 Code Number: bk04028 ABSTRACT The cotyledons of 3 –true leaf potted seedlings of a common Japanese tomato (i.e. cv. Fukuju No. 2) were mock inoculated with buffer only or singly and doubly with potato Virus X(PVX) and/or an attenuated strain (L11A) of tobacco mosaic virus (TMV-L11A) in greenhouse experiments. Time-course analysis of the methanol-extractable free, and saponifiable ester-bound phenol contents, as determined by the Folin-Ciolcalteau method, showed significantly higher accumulation in the primary leaves during the first 3 days in both mock and viral inoculated plants compared to the uninoculated controls. Whereas, however, the level in the healthy plants remained at about the same level thereafter, that in the infected plants continued to increase steadily peaking between 6 and 10 days postinoculation (dpi). Single infection induced significantly more free phenol accumulation than mixed infection at the peak period. The mean daily level of ester- bound cell wall phenols was about 12 – 20 times lower than that of the free phenols. It increased steadily in infected plants doubling its initial concentration as at 12 dpi but remained relatively unchanged in the healthy plants in the course of development. In contrast to the trend for methanol extractable free phenol accumulation, mixed infected plants had higher contents of saponifiable phenols than singly infected ones. Thin layer chromatography on silica gel G of the extracts from samples obtained at 12 dpi revealed at least 5 distinct bands (spots) that were common to both healthy and infected plants. However, the appearance of some extra bands that were highly florescent under long wave (320 nm) ultraviolet light on non florescent background silica gel, also indicating their phenolic nature, were limited to infected plants only. This indicates that viral infection not only affected the quantity but may also have altered the type of phenol components of the infected tomato plants. Key words: Tomato cv. Fukuju No. 2, mock and viral inoculations, phenolic components INTRODUCTION The infection, singly or doubly with potato virus X (PVX) and strains of tobacco mosaic virus (TMV) in cultivar Fukuju No. 2 tomato plant is known to involve alterations in the protein and carbohydrate (sugar and starch) metabolism of the host plant (1). The patterns of disease manifestation and dynamics of viral accumulation in such plants have also been reported as involving several folds enhancement of PVX concentration during the acute stage of disease and consequent manifestation of a synergistic disease i.e. extensive mosaic, and distortion coupled with considerable necrosis of the systemically infected leaves. Whereas single infection with PVX (O strain) produced moderate chlorotic mottling during the acute stage and some growth inhibition later on, TMV-L11A (an attenuated strain) inoculated alone induced very mild mosaic only during the chronic stage of disease (2, 3, 4). Host-pathogen interactions leading to growth inhibition and distortion of development often lead to an increase in the activity of certain enzymes, particularly polyphenol oxidases and decreased or increased activity of plant growth regulators. Furthermore, the deposition of phenols and often lignin is a well-known plant response to fungal attack, and it has been suggested that this response can be important for determining non-host resistance (15,16). A mycorrhizal strain of Laccaria laccata capable of inducing accumulation of osmiophilic materials, which histochemical tests revealed to be primarily phenolic, in the cortical cells of the primary roots of seedlings of Douglas-fir (Pseudotsuga menziensii) has been found (17). Series of tests led to the conclusion that phenol accumulation rather than antibiosis was the basis for the protection of such primary roots from lethal root rot by Fusarium oxysporum. Campbell and Ellis (6) reported enhanced phenolic metabolism, as indicated by tissue lignification and accumulation of specific methanol-extractable compounds in suspension cultured cells of a soft wood species, Pinus banksiama Lamb., as early as 12 h post elicitation by the ectomycorrhizal fungus Thelephora terrestris. Incompatible plant-virus interactions that result in hypersensitive response are also characterized by lignin synthesis. The plant forms necrotic lesions containing highly lignified cells around the pathogen (12). In compatible plant-pathogen interaction, increase in phenol concentration is often lacking (5). This, however, is not always the case as indicated by observations while working on stem rust infected, resistant and susceptible near-isogenic lines of wheat (14). Significant increase in phenol accumulation in wheat plants infected with the wheat streak mosaic potyvirus (WSMV) compared to the healthy controls has also been reported (9). Phenolic compounds produced by plants are formed through phenylpropanoid metabolism. However, since free phenols can be cytotoxic in the cytoplasm, plants sequester these compounds in the vacuole or deposit them in or on the cell wall. Once the phenolic acids or cinnamyl alcohols reach the cell wall, they may be either esterified or ether-linked to the cell wall polysaccharides or hemicelluloses, or be polymerized into lignin (10,11). Exhibition of biochemical response to viral infections vary for different plant systems. The objectives of this study, therefore, were to determine whether singly or doubly infected leaves contain different quantities and types of free and bound phenols as compared to leaves of mock inoculated or non inoculated, healthy tomato plants, at different times after inoculation. MATERIALS AND METHODS Plant propagation, virus inoculation and sample collection Seedlings of cv. Fukuju no.2 (a common Japanese cultivar) tomato, Lycopersicon esculentum Mill., were raised under greenhouse conditions in 1-litre capacity earthen pots that were arranged following completely randomized design, with each containing 4 plants. The two cotyledons of seedlings at the 2 to 3 true-leaf stage were inoculated singly by rubbing with a suspension of 0.2 mg of any of O strain of potato virus X (PVX), or L11A (attenuated strain) of tobacco mosaic virus (TMV) per ml of phosphate buffer (pH 7.0). Mixed inoculation was by combination of equal quantity of PVX and the TMV strain simultaneously applied. Plants that were mock inoculated with buffer only and those left uninoculated at all served as healthy controls. Leaves were dusted lightly with carborundum prior inoculations and washed with running water immediately afterwards. One of the two primary leaves of individual seedlings was collected thereafter, following pre-determined patterns, for phenol assay. Leaf samples were collected at the same time every morning to minimize the effects of diurnal variations. Whole leaves (without the petiole) were severed with a scalpel, weighed, wrapped in aluminum foil and frozen in liquid nitrogen. The samples were kept at –70oC when not assayed immediately. Extraction of free and bound cell wall phenols Free phenols were extracted from leaf samples of both healthy and infected plants essentially according to the method of Campbell and Ellis (6). Fresh tomato leaf samples that were taken at various times after inoculation, weighed individually and then frozen were powdered in liquid nitrogen using mortar and pestle. The powder was scooped into 5-ml polystyrene tubes with cap. Extraction was carried out with 2 vol. of 50% methanol for 1.5 h at 80oC in a water bath. The mixture was centrifuged for 5 min at 3,000 rpm and the supernatant used for the Folin-Ciocalteau assays. From the remaining pellets, phenolic acids esterified to the cell wall by ester linkages were saponified, according to the method of Funk and Brodelius (7), with 0.5 M NaOH in the ratio of 1 g of original sample: 4 ml for 24 h at room temperature. The mixtures were neutralized with one-quarter vol of 2 M HC1, centrifuged, and the supernatants also used for Folin-Ciocalteau assays. Determination of total phenols by the Folin-Ciocalteau assay Phenolic content of the methanol or NaOH extracts, described above, was determined by the method of Julkunen-Tiitto (8). Fifty microlitre (50μl) of extract was diluted to 1 ml with distilled water, and mixed with 0.5 ml of 2 M Folin-Ciocalteau reagent (Sigma) and 2.5 ml of 20% Na2CO3. The mixture was allowed to stand for 20 min at room temperature, and then the absorbance of the samples was measured at 725 nm using the Hitachi 1100U ultra Spectrophotometer. The phenol concentration was determined from a standard curve prepared with p-Coumaric acid (sigma). Two separate time-course experiments were carried out with samples replicated 4 times on each occasion. Thin layer chromatographic analysis of phenol extractsThin layer chromatographic analysis of the methanol extractable phenols was carried out using 1-butanol: acetic acid: water (at ratio 5:1:2) as the solvent system (6). Methanol extracts (5 ml) were taken to dryness under vacuum and then re-dissolved in 500μl of 50% methanol. A 10 μl aliquot of the redissolved extract was fractionated by TLC on a 10 cm long silica gel G layer using 1-butanol: acetic acid: water (at ratio 5:1:2) as the solvent system. After development of the chromatogram, phenolic compounds were visualized as short (254 nm) and long (320 nm) wave UV light absorbing spots on the florescent background or as florescent spots on the non florescent background and by their characteristic blue or blue-grey colour after Folin-Ciocalteau spray followed by exposure to ammonia fumes. The chromatogram was photographed under ultraviolet light at 320 nm wavelength. RESULTS Accumulation of free phenols Analysis of the methanol-extractable free phenols in plants under the various treatments from 1 dpi through 4 dpi, and in alternative days from 6 dpi through 12 dpi showed alterations in the accumulation of these compounds with age of infection (Table 1). The concentrations for the first 3 days showed that increased accumulation was significantly elicited by both viral and mock inoculations compared to non-inoculation. From the 4th day onward, however, infected plants whether singly or doubly inoculated, had significantly higher concentration (P=0.05), than any of the healthy plants. As shown in Figure 1, the average concentration per day in singly or mixed infected and healthy plants also differed considerably over time. Table 1: Comparative concentrations of accumulated methanol- extractable free phenolics in the primary leaf of seedlings of tomato cv Fukuju no 2, at different times after mock, single or double inoculation of the cotyledons with PVX and /or TMV.
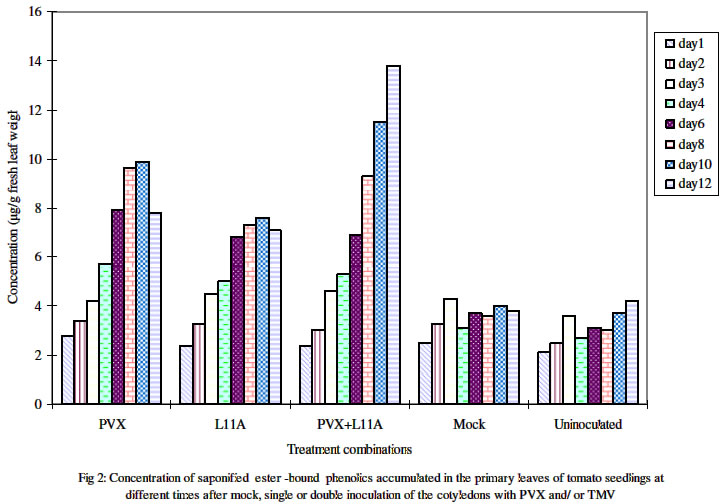
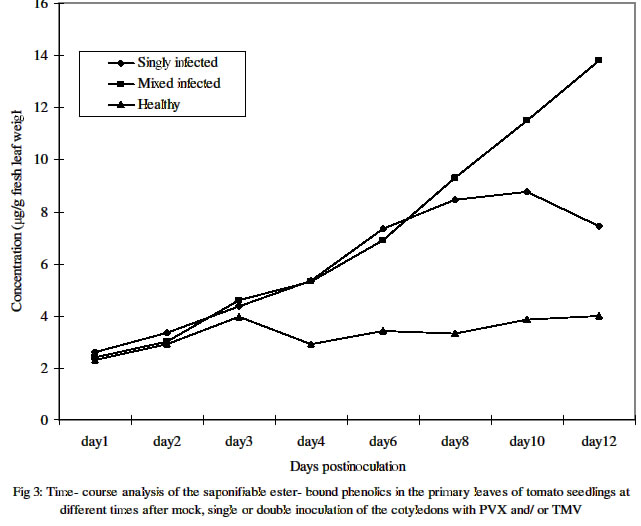
One primary leaf, i.e. the first true leaf was sampled daily from day 1 through day 12 post inoculation. Four different plants were assayed for individual treatments on any given day.Figures in the same column followed by the same letter are not significantly different at P= 0.05 using the New Duncan multiple range Test. Plants with the individual treatments varied in the days after inoculation at which the level of free phenols was highest. The highest levels in plants infected with PVX alone or PVX+L11A were at 8 dpi and 6 dpi respectively while that of plants with TMV-L11A alone was at 10 dpi. Meanwhile, the levels in the mock- inoculated and the uninoculated healthy controls were highest at 3 dpi and 8 dpi respectively. Table 2 shows the estimated average level of accumulated free phenols on any given day during the 12 days in which sampling and assays were carried out. It shows that plants with PVX alone (119 μg) followed by those with TMV-L11A alone (96 μg) had higher concentrations than the mixed infected (90.7μg) and the healthy plants. Accumulation of bound phenolsAnalysis of the saponifiable, ester- bound, cell wall phenols showed that the amount on any given day of assay was significantly lower than that of the free phenolics for any one treatment. As shown in Figures 2 and 3, the trend in healthy plants was similar to that of the free phenols concentration, which did not change much over the course of development. The trend in the infected plants was however that of steady increase over time before declining in singly infected plants. In mixed infected plants, the level increased dramatically from day 6 and did not decline throughout the duration of the experiment. At 12 dpi, the level was ca.14 μg/g of fresh leaf weight in doubly infected (PVX+L11A) plants as against 4.6 μg recorded for the same treatment at 3 dpi. The levels in plants singly infected with L11A or PVX at 3 dpi were 4.5 μg and 4.2 μg respectively. These however increased to 10 μg and 7.8 μg respectively at 10 dpi before declining at 12 dpi. The mean daily concentration in plants under the individual treatments is shown in Table 2. Unlike what obtained for free phenolics, mixed infected plants had significantly higher concentration than singly infected and healthy plants. Figure 3 shows the trends of the mean accumulation in singly (PVX alone and TMV-L11a alone) and doubly infected as well as in healthy plants (mock- and non-inoculated controls). Table 2: Mean daily concentration of methanol-extractable free, and saponifiable ester-bound phenolics in the primary leaf of cv Fukuju # 2 tomato seedlings over the first 12 days after inoculation of the cotyledons with PVX and TMV
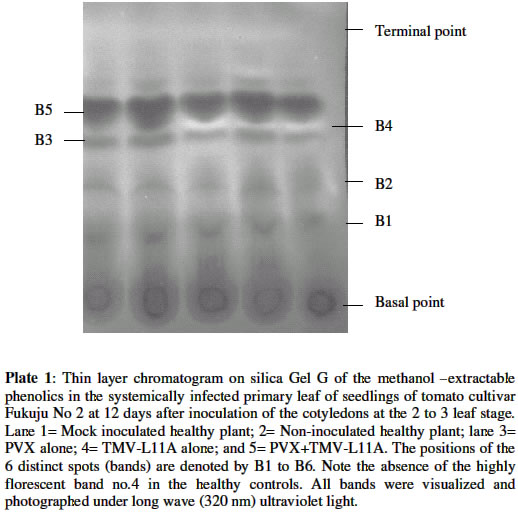
Each value is the average of concentrations obtained from assays carried out on samplescollected between 1dpi and 12 dpi. Four different plants were assayed for individual treatments on any given day.Figures in the same column followed by the same letter are not significantly different atP= 0.05 using the New Duncan Multiple Range Test. Chromatographic analysis of phenolic componentsPlate 1 is a thin layer chromatogram of the methanol-extractable metabolites in the systemically infected primary leaf of tomato sampled 12 days after mock or viral inoculation of the two cotyledons with PVX and/or TMV-L11A. Of the 6 distinct spots that are visible under short (254 nm) and long wave (320 nm) UV light, only Spots Nos 1, 2, 3, 5 and 6 are common to all the treatments. Spot No. 4, visible as an intense bluish-pinkish florescence was limited to infected plants only. Upon spraying with Folin–Ciocalteau reagent followed by exposure to ammonia fume, Spots Nos 1, 2, 3, 4, and 6 showed as blue- grey colour indicating that they were phenolic in nature. Spot No. 5, however, did not possess this attribute. In fact, it was visible to the naked eye as greenish coloration even before spraying. DISCUSSION
The total methanol-extractable phenols as well as wall bound phenolic compounds were more concentrated in infected plants than the healthy ones with increasing time following inoculation. Mock inoculation induced the same initial amount of upsurge as viral inoculation between 1 and 3 days postinoculation. Thereafter, however whereas the level in healthy plants stabilized that in the infected plants generally continued to rise. This showed that while increased accumulation of phenols may be triggered to some extent by minor injury, as witnessed in mock -inoculated plants, pathogenic infection, in this case viral, was evidently more important than injury in mediating this response in the tomato cultivar used in this study. Single infection with PVX or TMV-L11A at the peak period (i.e. between 6 and 10 days postinoculation) generally induced significantly more free phenols than mixed infection in the plants. This kind of response correlates with the relative mildness of the disease- inducing capabilities of the single infections compared to the mixed infection on the host tomato plants. In fact, as earlier reported (3), despite considerably high accumulation of the viruses both in the inoculated and systemically infected leaves, TMV-L11A induced no visible symptom at this stage of infection while only a very mild chlorotic mottling was visible in Fukuju No. 2 tomato plants singly infected with PVX. That increased accumulation of phenolic compounds was witnessed at all, however, suggests an initial attempt by the host to ward off whatever agent was responsible for its elicitation. Higher plants generally produce a great variety of secondary phenolic compounds, the role of which in the metabolism of the plant itself has not been adequately explained. Nevertheless, the deposition of phenols and often lignin is a well-known plant response to fungal attack, and it has been suggested that this response can be important for determining non-host resistance (15,16). Lignin deposition is correlated with an increased activity of enzymes in the phenylpropanoid pathway (13). A more or less steady increase in the level of the saponifiable wall bound i.e. deposited phenolics portion over time was observed in both mixed infected and singly infected plants. The highest level (almost twice the amount in singly infected plants) was found in mixed infected plants at 12 dpi. At this time, visual observation revealed some characteristic necrosis of the systemically infected uppermost leaves in the mixed infected plants. The increased compartmentalization of phenols in infected plants could be a failed attempt at resistance on the part of the host as it could be a consequence of successful invasion on the part of the virus. On the other hand, as reported previously (10,11), since free phenolics can be cytotoxic in the cytoplasm, plants sequester these compounds in the vacuole or deposit them in or on the cell wall where they may be either ester or ether linked to the cell wall polysaccharides or hemicelluloses, or be polymerized into lignin hence reducing their toxicity to the host plant. Thin layer chromatographic analysis revealed the presence of some phenolic spots in infected plants, which were absent in non-infected ones. This showed that in addition to the quantity accumulated, the type of the free phenols induced by viral infection was also altered. Kofalvi and Nassuth (9) had observed such alterations in wheat singly infected with wheat streak mosaic potyvirus (WSMV) in which significant increase in phenolics accumulation in infected wheat plants, compared to the healthy controls, was also recorded. REFERENCES
© 2004 Nigerian Society for Experimental Biology. The following images related to this document are available:Photo images[bk04028p1.jpg] [bk04028f1.jpg] [bk04028f2.jpg] [bk04028f3.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}