 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biokemistri, Vol. 17, No. 1, June, 2005, pp. 19-25
In vitro effects of aqueous extracts of Zanthoxylum macrophylla roots on adenosine triphosphatases from human erythrocytes of different genotypes Iheanyichukwu ELEKWA1, Michael O. MONANU 2* and Emmanuel O. ANOSIKE2 1Department of Biochemistry, School of Biological Sciences, AbiaStateUniversity,
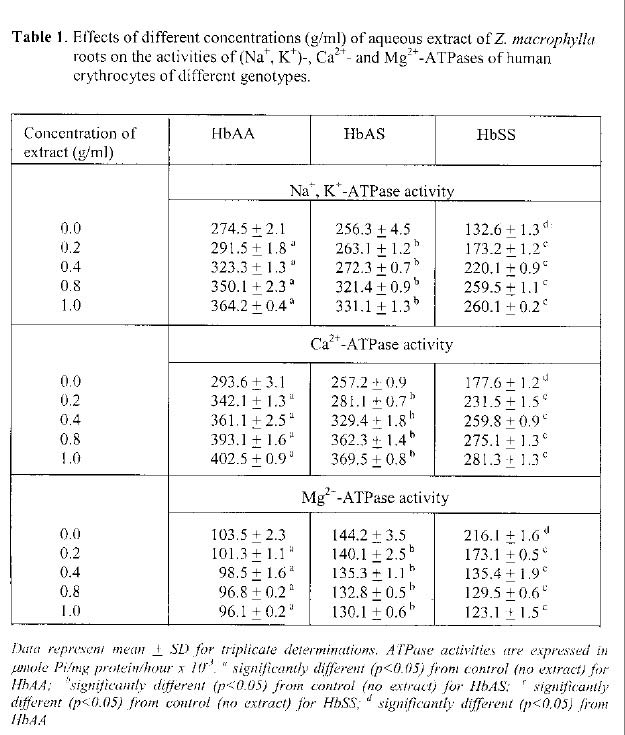
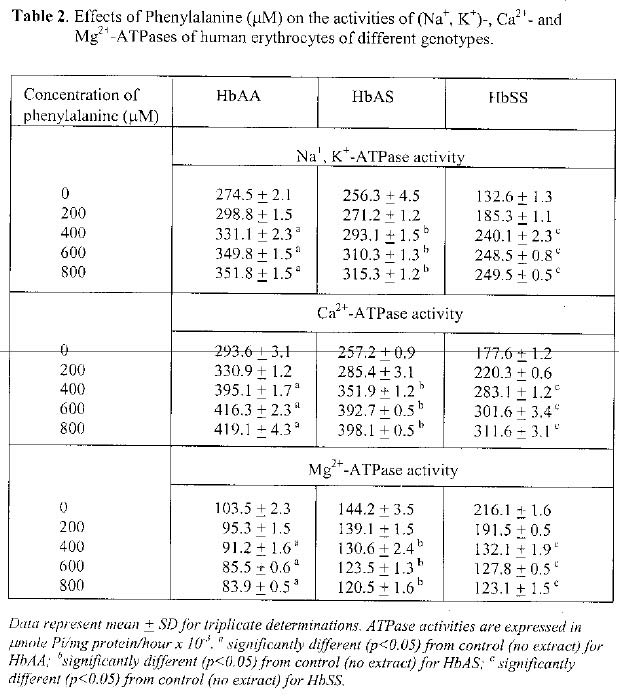
Uturu, Okigwe Received 27 February 2004 Code Number: bk05004 ABSTRACT The in vitro effects of aqueous extracts of Zanthoxylum macrophylla roots on the activities of three ATPases (Na+, K+- ; Ca2+- ; and Mg2+-ATPases) were investigated in membrane preparations from human erythrocytes of HbAA, HbAS and HbSS bloods. The activities of these membrane-bound enzymes were determined in the presence of varying concentrations of the plant extract. The results indicated that for Na+, K+- and Ca2+-ATPases, a dose-dependent activation of the three enzymes by the aqueous plant extract occurred. The trend for the different genotypes was HbSS>HbAS>HbAA. The activations were significant (p<0.05) when compared with activities obtained in the absence of the plant extract. Phenylalanine, a known anti-sickling agent, at varying concentrations (200-800µM) produced similar activating effect. In contrast, the extract inhibited the Mg2+-ATPase activity in a dose-dependent manner with a trend of HbSS>HbAS>HbAA. The inhibitions seen were significant (p<0.05) in comparison with activities in the absence of the plant extract. Phenylalanine produced similar effect for Mg2+-ATPase. These findings in this study suggest that differences in the ATPases of the different genotypes may account for the reports of other workers on the involvement of ion transport as a factor in the physiological manifestation of sickle cell disease. The findings are also significant to an understanding of the molecular basis for the use of the aqueous extract of Z. macrophylla roots in the management of sickle cell disease.
INTRODUCTION
The unidirectional active transport of ions such as Na+, K+ and Ca2+, has been reported to play major roles in maintaining the stability of the erythrocyte membranes (1, 2). In deed, the presence of three different adenosine triphosphatases (Na+, K+-; Ca2+-ATPase; and Mg2+-ATPase in human erythrocyte membranes had been reported by various workers (3, 4, 5). The determinations of the activity levels of these ATPases in different human genotypes showed that (Na+, K+- and Ca2+-ATPases) were significantly lower in HbSS erythrocytes, while Mg2+-ATPase was significantly higher than for HbAA erythrocytes (5, 6). The role of Ca2+ in preserving the low permeability of the red cell to K+ and Na+, thereby maintaining the normal low rate of cation leakage from the cell had been reported (7). In addition, elevated intracellular Ca2+ had been suggested to destroy the normal cell shape and plasticity leading to increase K+ leakage hence inhibiting Na+, K+-pump (7). Indeed, it had been suggested that sickling of red cells could be reversed if the excess Ca2+ in the red cells is pumped out (8). There are reports that pharmacological agents that are capable of altering the permeability of cell membranes could be useful in the management of sickling, a major manifestation in sickle cell disease (9, 10). A number of plant products, some of which had long been used in trado-medical practice, have shown promising returns as possible agents for the amelioration of sickling of erythrocytes (13, 14). One of such, Zanthoxylum macrophylla (formerly Fagara zanthoxyloides) gained attention following the report by El-Said et al (13) that aqueous extracts of the roots of Z. macrophylla, in addition to having anti-microbial activity, also preserved the colour of the blood within its zone of anti-microbial action. Further work by Sofowora and Isaac-Sodeye (14) showed that the crude aqueous extract of Z. macrophylla roots reverted metabisulphite-induced sickling of HbSS and crenated HbAA erythrocytes to normal configuration. The plausible agent for this effect was shown to be 2-hydroxymethylbenzoic acid (15). A recent report (6) indicated that the aqueous extract of Garcinia kola, another trado-medical plant, commonly consumed in Nigeria, activated Na+, K+- and Ca2+-ATPases of HbAA, HbAS and HbSS erythrocyte preparations. Inhibition was observed for Mg2+-ATPase in the three genotypes. In both instances, greatest effects were observed for the HbSS genotype. The present study aims at investigating the possible effects of aqueous extract of Z. macrophylla roots on ATPases of human erythrocytes of different genotypes in view of the earlier suggestions that the control of ion flux in erythrocytes, possibly by these ion ‘pumps’, could play a role in maintaining membrane stability as well as cell volume/shape. The study also aims at providing further support, at the molecular level, on earlier reports on the action of the extract in the sickling process. MATERIALS AND METHODS Adenosine triphosphate (sodium salt) was purchased from Riedel De Haen, AG Germany, while heparin was from Sigma Chemical Co., St. Louis, U. S. A. All other reagents were of the purest grades commercially available. The Z. macrophylla root samples were purchased from the herbal line at Ogbete main market, Enugu, and were authenticated at the National Root Crops Research Institute, Umudike. Blood collection The different blood samples were collected, by veni-puncture into lithium heparinized sterile tubes, from consenting volunteers who visited the University of Port Harcourt Teaching Hospital, University of Nigeria Teaching Hospital, Enugu and the MedicalCenters of the University of Port Harcourt and University of Nigeria, Nsukka campuses. The genotypes of the samples were determined using standard electrophoresis procedure, and the samples stored at 4°C in the refrigerator. They were used within 24 hours of collection. Preparation of Z. macrophylla root extract Following washing with distilled water and air-drying, the roots were ground into fine powder using an electric grinder. The powder was then extracted in distilled water (100g / 250 ml) for 24 hours before the resultant mixture was filtered through a Whatman #1 filter paper. The filtrate was stored at 4°C and was the stock solution for the various determinations of the effect of the plant extract in this study. Preparation of erythrocyte membrane This followed the method described by Hamlyn and Duffy (16). Briefly, the whole blood was centrifuged at 5, 000 g for 10 minutes and the resultant precipitate washed with 0.15 M NaCl (pH 7.4). This ‘washing’ process was repeated thrice, the final precipitate lysed by swirling in 5 mM NaH2PO4.2H2O (pH 7.7), and then centrifuged at 5,000 g for 10 minutes. The resultant precipitate was ‘washed’ with 10 mM Tris-HCl (pH 7.7) and suspended in 3 ml distilled water. The isolated membranes were stored at 4o C and used within 12 hours of collection of blood samples. Enzyme assays The assay of the enzyme activities followed the procedure of Hesketh et al (17) and monitored the inorganic phosphate released from ATP. Enzyme activities were expressed as µmole Pi/mg protein/hour x 10-3. Na+, K+-ATPase: For this enzyme, the reaction mixture contained 0.5 ml each of 0.35 M NaCl, 17.5 mM KCl, 21.0 mM MgCl2,10 mM Tris-HCl (pH 7.4), and 8.0 mM ATP-Na2. The reaction was started by the addition of 0.2 ml of erythrocyte membrane preparation and the mixture incubated at 37°C for 1 hour. The reaction was terminated by addition of 0.8 ml of ice-cold 10% (w/v) trichloroacetic acid and the resultant mixture stood for 20 minutes at 4°C. It was then centrifuged at 4, 000g for 5 minutes using a bench-top centrifuge. The concentration of phosphate in 1 ml of the supernatant was measured by the method described by Fiske and Subbarow (18). For this, 1.0 ml of 2.5% ammonium molybdate was added and after 10 minutes, the addition of 0.1 ml of 2% ascorbic acid followed. The mixture was kept at room temperature for 20 minutes for colour development. The absorbance of the final mixture was measured at 725 nm using SP 6-200 spectrophotometer. Ca2+-ATPase: The reaction mixture contained 0.5 ml each of 21 mM MgCl2, 17.5 mM CaCl2, 10 mM Tris-HCl (pH 7.4) and 8.0 mM ATP-Na2. The reaction was initiated and processed as described for Na+, K+-ATPase assay. Mg2+-ATPase: The reaction mixture contained 0.5 ml each of 21 mM MgCl2, 10 mM Tris-HCl (pH 7.4) and 8.0 mM ATP-Na2. The initiation and processing of the reaction followed the procedure described for Na+, K+-ATPase assay. Protein determination The protein concentrations of the membrane preparations were carried out by the method of Lowry et al (19), using bovine serum albumin as standard protein. Effects of the aqueous extract on the ATPases of erythrocytes from the various genotypes The reaction tubes, in addition to the contents reported above for the various ATPases, contained 0.5 ml of different concentrations of the aqueous plant extract to produce the desired final concentrations in the total reaction mixture of 1 ml. The reaction was further processed as described for the Na+, K+-ATPase assay. Statistical analysis The student t-test was applied at 5% confidence level. RESULTS The levels of Na+, K+- and Ca2+-ATPases, in the absence and presence pf varying concentrations of the aqueous extract of Z. macrophylla roots are shown in Table 1. For Na+, K+-ATPase, the enzyme activities (µmole Pi/mg protein/hour x 10-3) in the absence of the plant extract were 274.5 + 2.1 (HbAA), 256.3 + 4.5 (HbAS) and 132.6 + 1.3 (HbSS). The values for HbSS genotype was significantly (p<0.05) lower than the HbAA genotype. The activities in the presence of plant extract showed increases with increasing concentration of the extract. A statistically significant increase (p<0.05) in comparison with activities in the absence of the plant extract was noted for all genotypes. For Ca2+-ATPase, enzyme activities in the absence of plant extract were 293.6 + 3.1 (HbAA), 257.2 + 0.9 (HbAS) and 177.6 + 1.2 (HbSS). The activity for HbSS genotype when compared with HbAA genotype was significantly lower (p<0.05). In the presence of varying concentrations of the plant extract, the activities showed dose-dependent increases. The increases for HbAA, HbAS and HbSS genotypes was significant (p<0.05) when compared with values in the absence of the plant extract. For Mg2+-ATPase, the enzyme activities in the absence of the plant extract was 103.5 + 2.3 (HbAA), 144.2 + 3.5 (HbAS) and 216.1 + 1.6 (HbSS). A significantly higher value (p<0.05) for HbSS genotype in comparison with HbAA genotype was obtained. A dose-dependent decrease in activity was obtained in the presence of varying concentrations of the plant extracts for this enzyme. The decreased obtained for the three genotypes were significant (p<0.05) when compared with values in the absence of the plant extract. The results of the effect of varying concentration of phenylalanine on the activities of the three ATPases are shown in Table 2. For Na+, K+-ATPase, significant increases (p<0.05) in activities for the three genotypes were obtained in the presence of 400, 600 and 800 µM concentrations when compared to the activities in the absence of phenylalanine. A similar trend was seen for Ca2+-ATPase activity. For Mg2+-ATPase, the activities for the three genotypes showed dose-dependent inhibition by phenylalanine, which were significantly different (p<0.05) when compared to the values in the absence of this amino acid. DISCUSSION As shown in Table 1, the levels of Na+, K+- and Ca2+-ATPases, in the absence of the aqueous extract of Z. macrophylla roots ,were significantly (p<0.05) lower in HbSS erythrocytes than HbAA and HbAS erythrocytes. For Mg2+-ATPase, the level was significantly (p<0.05) higher in HbSS than HbAA and HbAS erythrocytes. These findings agree with earlier report on the relative activities of the three ATPases in human erythrocytes of different genotypes (4, 8, 20). The presence of varying concentrations of the aqueous extract of Z. macrophylla roots had various effects on the ATPase activities. An apparent dose-dependent activation was observed for Na+, K+- and Ca2+-ATPases while an inhibition was noted for Mg2+-ATPase (Table 1). At 0.4 g/ml of the extract, percentage activations of 18%, 6% and 65% were noted for Na+, K+-ATPase from HbAA, HbAS and HbSS erythrocyte, respectively. At the same concentration of the extract, percentage activations obtained for Ca2+ -ATPase were 22%, 28% and 46%, for HbAA, HbAS and HbSS erythrocytes, respectively. The percentage inhibition seen for the effects of 0.4 g/ml of extract were 5%, 6% and 37% for HbAA, HbAS and HbSS erythrocytes, respectively. The trends seen in the present study were comparable to that reported for the aqueous extract of Garcinia kola (Henkel) seeds (6). However, the relative effects were higher for G. kola (Henkel) seed extracts when compared to Z. macrophylla roots, suggesting that G. kola (Henkel) seed extracts may have better actions on the possible regulation of the cellular effects performed by these ATPases. Furthermore, the activations/inhibitions for HbSS was remarkably higher than for HbAA and HbAS genotypes suggesting more ‘benefits’ of the effects on the ATPases for HbSS individuals. Phenylalanine, a known anti-sickling agent (12), at 400 µM, activated both Na+, K+- and Ca2+-ATPases, but inhibited the Mg2+-ATPase activity of HbAA, HbAS and HbSS erythrocytes membrane preparations (Table 2). The modulation of the activities of the three membrane-bound ATPases (increases for Na+, K+- and Ca2+-ATPases, and decrease for Mg2+-ATPase) by the plant extract noted in this study suggest vital roles for the plant extract in the maintenance of the shape/volume of erythrocytes. It further provides some clues at the molecular level on the possible roles of the ATPases in the sickling of erythrocytes, and of the aqueous extract of Z. macrophylla roots in reversing this sickling (14, 15) thereby supporting its use in the therapy of the sickle cell patients. ACKNOWLEDGEMENT The authors are grateful for the financial support from the Senate Research Committee, University of Port Harcourt. REFERENCES
© 2005 Nigerian Society for Experimental Biology The following images related to this document are available:Photo images[bk05004t1.jpg] [bk05004t2.jpg] |
| |||||||||

{kind=link}
{kind=link}