 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
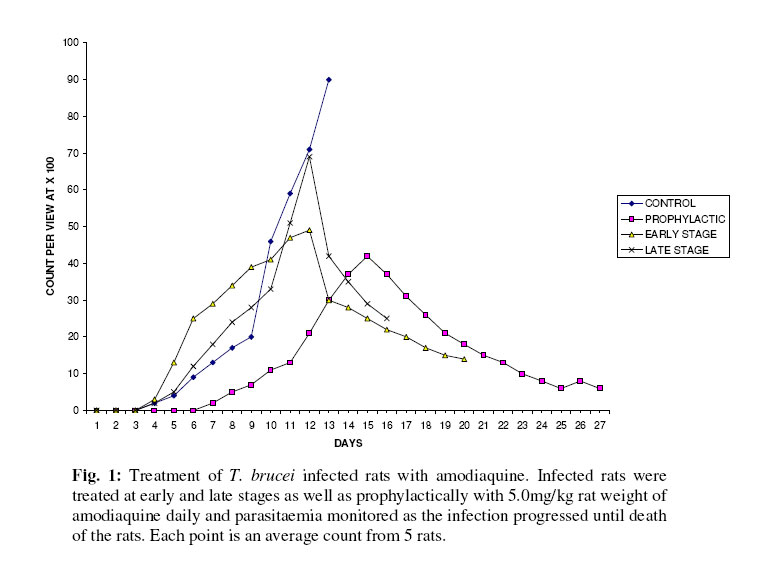
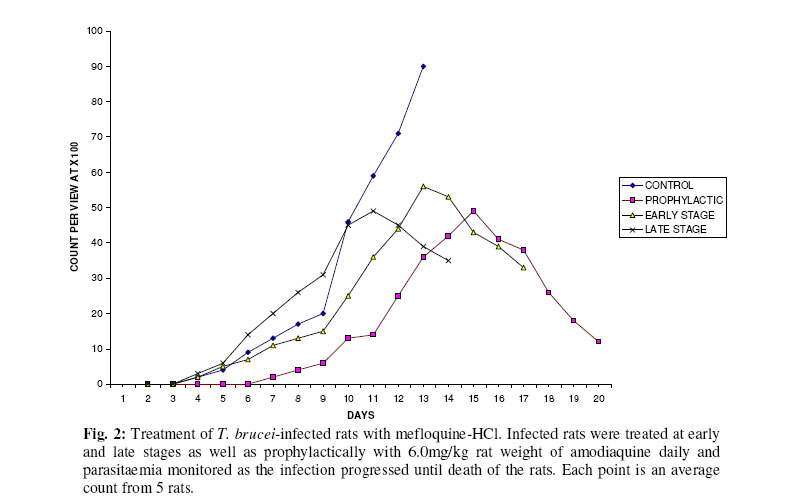
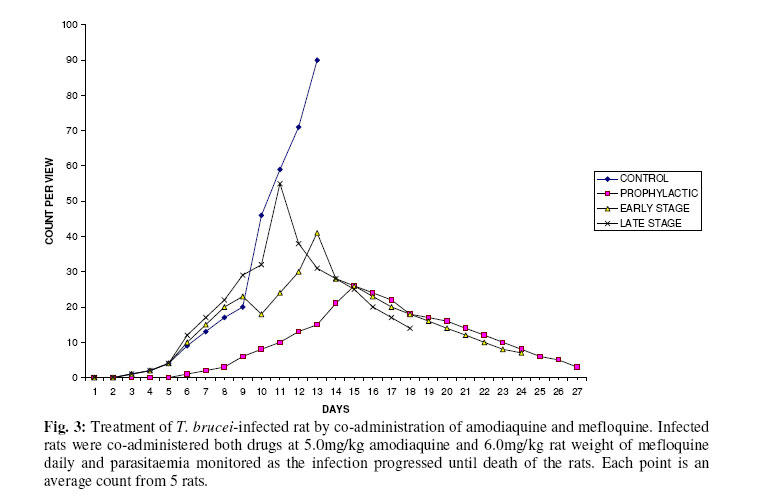
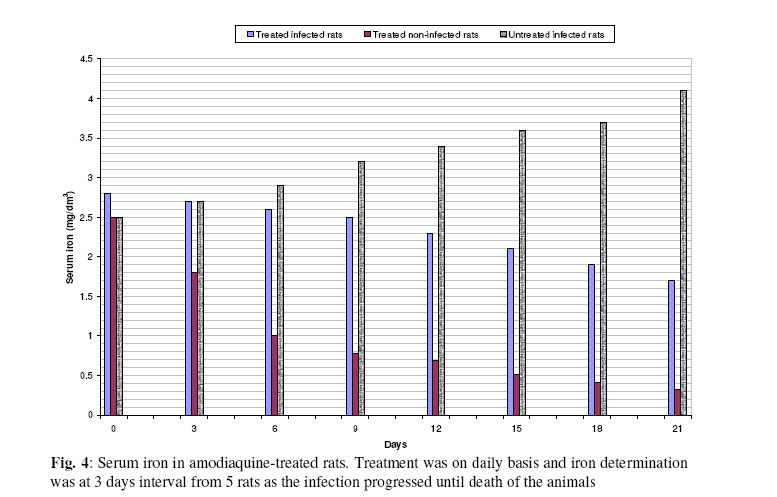
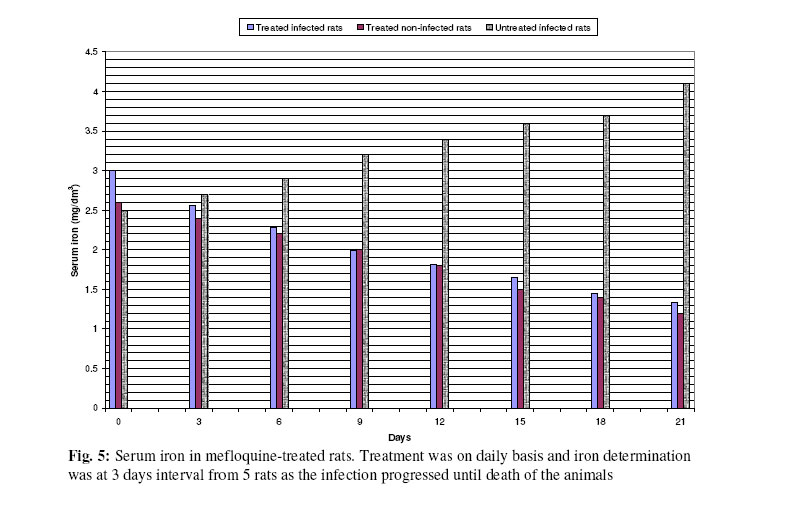
Biokemistri, Vol. 17, No. 2, Dec, 2005, pp. 115-121 Therapeutic properties and serum iron in T. brucei infected rats treated with amodiaquine and mefloquine Justine T. EKANEM*, Faoziyat A. SULAIMAN and Oluyomi ADEYEMI Trypanosomosis
Research Unit, Department of Biochemistry, University of Ilorin,
Ilorin, Nigeria Received 28 December 2004 Code Number: bk05016 Abstract Serum iron was monitored in Trypanosoma brucei-infected rats treated with Amodiaquine and mefloquine antimalarials as the infection progressed. The chemotherapeutic properties of the drugs against African sleeping sickness were also assessed. Results show gradual reduction in the levels of serum iron in the infected but treated rats as the infection progressed. Results also show that reduction in serum iron level is more pronounced in amodiaquine treated but uninfected rats than the mefloquine treated rats. Serum level in infected but not treated rats increased steadily from the day of infection. For prophylactic treatments of infected rat, amodiaquine extended the lifespan of the rats for 14 days while mefloquine extended it for 7 days. For early stage treatment, amodiaquine and mefloquine extended life span for 7 and 4 days respectively while late stage treatment the extensions were 2 and 1 day respectively. Results suggest that these antimalarials especially amodiaquine could be useful in the clinical management of African sleeping sickness and that this may be through reduction of blood iron level, a situation that can inhibit ribonucleotide reductase of the proliferating parasites. Key words: Amodiaquin, mefloquin, T. brucei, serum iron INTRODUCTION African trypanosomosis continues to be prevalent in man and domestic animals in Africa especially in the sub-Saharan region1 where it is estimated that about 300,000 new cases are reported annually2 in the face of the inadequacies of the available drugs3. The continuously changing epidemiology4 and the vast and uncontrollable animal reservoir hosts5 make it even more difficult for epidemiological control. The causative agent of the disease, Trypanosoma brucei requires iron for its growth6. The blood stream form of the parasite obtains iron through a well developed transferrin system7 from the host blood. Intracellularly, the iron-dependent, cell cycle related enzyme, ribonucleotide reductase plays a central role in cell division and proliferation8. This iron-requiring RNR is presented in T. brucei9,10 and it has been suggested that it can be exploited for the development of cheap antiparasite therapies including African trypanosomiasis11-14. Drugs of the quinine family have also been suggested to have iron-chelating properties15,16. In this report, we have assessed the trypanocidal properties of amodiaquine and mefloquine hydrochloride in T. brucei-infected rats and have also determined the levels of serum-iron in the treated rats. MATERIALS & METHODS Experimental animals Albino rats weighing approximately 260g were obtained from the animal house of the Department of Biochemistry, University of Ilorin, acclimatized for about 14 days and fed with animal feed obtained from Bendel Feeds and Flour Mills Ltd, Ewu, Edo State, Nigeria and water was provided ad libitum Parasite Strain Lafia strain of T. brucei was obtained from veterinary & livestock studies Department of Nigerian Institute for Trypanosomiasis Research, Vom, PlateauState of Nigeria. The parasite was maintained by repeated passages into other rats. 0.5ml of parasite suspension was inoculated into the peritoneal cavity of uninfected rat. The suspension contained 3 or 4 trypanosomes per view at x100 magnification. Animals were inoculated with the parasite and parasite count carried out on daily basin to monitor infection progress until the animals died. Experimental drugs and administration Amodiaquine HCl was a product of Pfizer Afique de l’Ouest-B.P.-Senegal while Mefloquine HCl was a product of Roches, USA. Amodiaquine was administered to rats intraperitoneally on daily basis for the period of the experiment at 5.0mg/kg rat weight while mefloqoiune was administered at 6.0mg/kg rat weight. Prophylactic treatment started 72 hours before infection and continued as the infection progressed until death. The early stage treatment started on the day parasite was first sighted in the blood while late stage treatment was commenced 10 days post-infection and respectively continued until the animals died. Control animals were either infected but not treated with drugs or uninfected but treated with drugs. Serum iron determination Iron was determined by the use of 1,10 phenanthroline as described by Faulkner et al17 Parasitaemia This was obtained by counting the number of parasites per view under light microscope at x100 magnification from a thin blood smear obtained from the tip of the tail of an infected rat. RESULTS Treatment with amodiaquine Prophylactic treatment shows the lowest parasitic load or peak compared with the control, early and late stage treatments. It was able to extend life span of infected rats by 14 days over control infected rats while early stage treatment extended life span by 7 days. Late stage treatment at high parasitaemia decreased from day 12 till day 16 when the rats died extending the lifespan by 4 days (Fig. 1). Treatment with mefloquine HCl In the prophylactic treatment, (Fig. 2) the life span of infected rats was extended from 13 days for control to 20 days (7 days). The parasitaemia remained low compared with control throughout the experiment. Early stage treatment extended life span by 4 days over the control while there was no significant difference between the late stage treatment and the control as the life span of the rats was extended by only one day. Treatment with drug combination Rats co-administered with mefloquine and Amodiaquine showed an extensively lowered parasitaemia compared to rats singly administered with the drugs and it extended life span by 14 days (Fig. 3). Similarly early and late stage treatments extended life span by 11 and 5 days respectively over the control. Serum Fe in infected rats It was observed (Figs. 4 & and 5) that over a period of 21 days, the serum iron in infected and uninfected rats kept decreasing with daily administration of both drugs. With the uninfected rats, the decreasing trend was more pronounced in amodiaquine-treated rats (Fig. 4) than the Mefloquine-treated (Fig. 5). The treated infected rats showed gradual rise in serum iron as the infection progressed. DISCUSSION Iron is an essential nutrient for most bacteria and has been identified as an important factor in the establishment of infections18,19. The effect of trypanosome infection on rats leads to erythrocyte lyses and anemia20. Ribonucleotide reductase is an iron-requiring enzyme which plays a central role in DNA replication, cell division cycle and proliferation. This enzyme provides the deoxyribonucleotide (dNTP) precursors in DNA replication of the cell division cycle. It is the only enzyme along the cell division cycle that requires iron not as part of a prosthetic group but as part of the free iron pool21,22. The recombinant ribonucleotide reductase enzyme of trypanosomes contain iron-tyrosyl free radical centre9,10 thus suggesting that the native enzyme could be subjected to regulation through iron deprivation. Various reports11-14 have indicated that iron-chelation could be a potent means of killing African trypanosomes in vitro and in vivo and the prime suspect as target is the enzyme ribonucleotide reductase (RNR). It has been reported that blood stream forms of trypanosomes increase their transferring uptake from hosts by approximately 3 folds in iron depletion7,24. Prolonged iron depletion can therefore inhibit ribonucleotide reductase activity leading to cell death. The eventual death of these rats could be due to advanced haemolysis20. It could also be due to toxic extra-cellular factors released by the parasite25-29. The reduced parasitaemia and extension in life span observed in Figs. 1,.2 and 3 are possibly due to reduction in blood iron levels (Figs. 4 & and 5) as a result of the administration of the drugs. Parasite load in the blood has a relationship with observable pathogenic effects20. Amodiaquine is more potent in the reduction of the blood iron level (Fig. 4) and probably explains its ability to extend the life span of infected rat much more than mefloquine (Fig. 5). Combination of amodiaquine with mefloquine is not explorable since it gives no additional days to the extended life span shown by amodiaquine alone. It is believed that the infected rats still died after treatment with amodiaquine and a lowered parasitaemia because of extracellular factors that may have been released by trypanosomes into the blood stream25-29. We suggest that amodiaquine could be a useful agent in the management of African trypanosomosis and could be even more useful in prophylactic circumstances. REFERENCES
© 2005 Nigerian Society for Experimental Biology. The following images related to this document are available:Photo images[bk05016f1.jpg] [bk05016f5.jpg] [bk05016f4.jpg] [bk05016f2.jpg] [bk05016f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}