 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
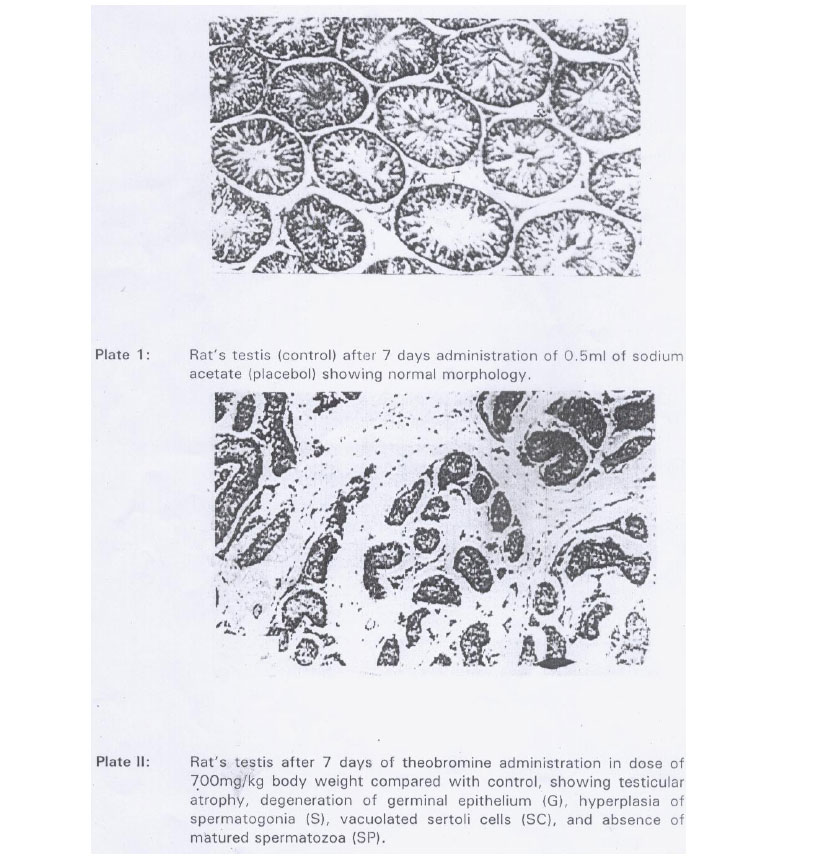
Biokemistri, Vol. 17, No. 2, Dec, 2005, pp. 123-128 Theobromine induced seminiferous tubular lesion with elevated serum testosterone levels in male Wistar rats Mbeh U. ETENG*, Eyong U. EYONG, G. O. IFERE and N. CHUKWUEMEKA Department of Biochemistry, College of Medical Sciences,
University of Calabar, Calabar, Nigeria Received 2 March 2004 Code Number: bk05017 Abstract Concentrations of serum testosterone, albumin, transferrin, total serum protein and testicular histology of male albino wistar rats were evaluated following administration of 700mg/kg body weight theobromine in 0.5ml 14% sodium acetate for seven days. Theobromine administration produced significant increase in serum testosterone concentration (P < 0.05) and significant decreases in serum albumin and total serum protein (P < 0.05) when compared with the controls. The decrease in serum transferrin was not significant (P > 0.05) compared with the control and there were seminiferous tubular lesions. The results suggest in part, structural damage on the tubules which may affect its function but the extent to which its function is impaired is not known. Key words: Theobromine, Seminiferous tubular lesion, Serum testosterone and proteins Introduction Theobromine, a methylxanthine alkaloid is widely enjoyed in coffee and cocoa beverages and as an ingredient in many prescriptions and over the counter medications such as stimulants, analgesics and diuretics1. The testes have been reported to be the target sites of theobromine toxicity2. The alkaloid is known to move freely between the plasma and testes due to the fenestrated nature of the testes capillaries, which permit free passage of large molecules3. Purine and Pyrimidine base nucleotides are the components of the deoxyribonucleotide (DNA). As a purine base analogue, theobromine may affect the reproductive function of the testes, viz production of spermatozoa and biochemically, the biosynthesis of testosterone and the levels of some serum transport proteins. This study was designed to investigate the effect of theobromine exposure on serum testosterone, albumin, transferrin, total serum protein and testicular histology in male albino wistar rats. MATERIALS AND METHODS Theobromine Pure theobromine (anhydrous white powder) was purchased from British drug House Chemical Company, England and used for the study. A theobromine stock solution was prepared by dissolving 22.5g anhydrous theobromine powder in 100ml of sodium acetate and the dose used for study obtained from this stock by appropriate dilution. Animals Sixteen male albino rats of the wistar strain were obtained from the disease-free stock of our departmental animal house and reared on a popular commercial stock diet, Pfizer rat chow (Pfizer Livestock Feeds, Aba, Nigeria) until they were used for the experiment. The rats were randomly assigned into two groups (Control and test) of eight animals each. The animals were housed in Perspex cages with plastic bottom grid and steel top and kept under adequate ventilation at room temperature and relative humidity 26 ± 2°c and 46% respectively. Food and water were provided ad libitum. Chemical compound administration Animals in the test group were gavaged with 700mg/kg body weight of theobromine in 14% sodium acetate, while the control was gavaged with 0.5ml of sodium acetate solution (placebo). Chemical compound administration continued daily between the hours of 08:00 and 09:00am each day4. Blood sample collection Twenty-four hours after the last dose was administered, the animals were anaesthetized in chloroform vapour, quickly brought out of the jar and dissected. Whole blood collected by cardiac puncture from each animal into clean, dry centrifuge tubes, were allowed to stand for about 1hour to clot, and further centrifuged at 10,000 rpm for 5 minutes using an MSE table top centrifuge4. Serum was separated from the clot with Pasteur pipette into sterile screw cap samples tubes and used for the assay of serum testosterone and albumin, total serum protein and transferrin, testes from the right side of both the control and test animals were removed and immediately fixed in 10% buffered formalin solution after washing with physiological saline for histological studies6. Serum testosterone assay The serum samples were analyzed in triplicates for testosterone level by the enzyme linked immunosorbent assay (ELISA) method of Engrall and Perlman5, using the micro well testosterone EIA Kit obtained from syntron Bioresearch Inc. England. In principle, testosterone present in the sample competes with enzyme labeled testosterone for binding with anti- testosterone antibody immobilized on a micro well surface. The amount of labeled testosterone that form conjugates will decrease in proportion to the concentration of the unlabeled testosterone in serum sample. The unbound sample and conjugate are then removed by washing and colour developed. The intensity of the colour measured at 450nm reflects the amount of bound enzyme testosterone conjugate and is inversely proportional to the concentration of testosterone within the dynamic range of the assay. The absorbance was read with micro well reader, percentage transmittance calculated and testosterone concentration in sample obtained from the standard testosterone calibration curve and expressed in ng/ml. The assay sensitivity is reported as 0.85ng/ml. Assay of serum albumin in total serum protein To assay for the level of albumin in the serum, globins were precipitated from serum proteins using diethyl ether, leaving albumin in the aqueous phase. The serum albumin was then assayed by the biuret method of Doumas et al6. In principle, copper ions in biuret reagent reacted with peptide bonds of the protein to give a violet coloured complex in an alkaline medium. The colour intensity read at 540nm from a digital spectro photometer (spectro uv-vis RS, labo med, Inc, Canada) was compared with an albumin protein standard. Similarly, for total Serum protein, the biuret method was employed but whole serum, (not treated with the protein precipitant, diethyl ether) was used. Assay of serum transferrin Serum transferrin was determined as total iron building capacity (TIBC) using the mg (CO3)2 method in combination with international committee for standardization in haematology, ICSH7. The value of TIBC in Ug/100ml serum was divided by a factor 1.45 to obtain the level of transferrin in mg/dl of serum8. Histological studies Following fixation of the right side testes from both control and test animals, tissue sections were processed according to the method of Druny and Wallington9. the tissue sections embedded in paraffin wax were further cut into ribbon slices of about 5.0µm in thickness. The slices were stained with Hematoxylin and Eosin, and then mounted onto a light microscope (magnification X 160) slides using DPX for histological examination. Statistical analysis Pair-wise comparison of the data was done using the Student’s t test10. Values were regarded significant at P < 0.05. RESULTS Effects of theobromine on Serum Testosterone, Transferrin, Albumin and total protein levels The effect of theobromine on serum testosterone, transferring albumin and total serum protein levels of wistar rats are summarized in table 1. Administration theobromine produced a significant increase in testosterone levels relative to the control group (P<0.05), but both serum albumin and total protein levels were significantly reduced in the treatment group compared to the control (P <0.05). The decrease in the mean ± SEM values of serum transferrin were not significant compared to the controls (P>0.05). Histological Observation In the test group (testes from rats administered theobromine in dose 700mg/kg body weight for 7 days), no detectable histological changes were found in the leydig cells. However, there were some testicular lesions affecting the seminiferous tubular cells. These ranged from hypertrophy of the seminiferous tubules, degeneration and necrosis of germinal epithelium hyperplasia of spermatogonia, focal necrotic changes of spermatids and spermatocytes. Sertoli cells were also vacuolated (plate 2). DISCUSSION Thebromine administration of 700mg/kg body weight on daily basis for 7 days increased serum testosterone levels but decreased serum transferrin, total serum protein, serum albumin levels and induced testicular lesions, observed mainly on the seminiferous tubules which may adversely affect its function. The histological aspect of this study agrees with the previous reports of Soffieti et. al.2 in rabbits, a different rodent specie from the one used in this study. Although the pathogenesis of theobromine induced testicular damage still remains unclear, we are led on account of the observed histological changes including congestion and haemorrhage (plat 2) to suggest that vascular damage may play a role in the development of testicular toxicity. Secondly, according to Levi11 and Kimmel et. al.12, caffeine and therobromine via its sympathetic nerve supply induce vasoconstriction of placental and abdominal organs including the gonads leading to nutrient deprivation in the affected organs. These effects may therefore retard germ cell development and differentiation into spermatozoa. Several factors explain the observed rise in serum testosterone concentration. First theobromine induces accumulation of CAMP13 which is known to trigger three responses viz: changes in the state of phosphorylation of specific proteins, induction of protein synthesis and stimulation of hybrid synthesis. These processes bring about the transport of cholesterol through the mitochondria membranes and further transformation into steroid products in the endoplasmic reticulum 14, 15, 16. However, these require intact leydig cells. It is therefore reasonable that the intake leydig cells retained their metabolic competence to synthesize testosterone hence the increase in serum testosterone levels observed in this study. Besides this, other non-gondal tissues like the adrenal gland synthesize and secrete weak androgens, which may be converted to testosterone. In fact the major cAMP sensitive proteins of the adrenal cortex have been documented to be those associated with steroids biosynthesis17. These cells come under direct influence of cAMP whose accumulation is elicited by theobromine13. Total serum protein, albumin and transferrin all decreased. Albumin along with transcortin serves as a transport vehicle for testosterone since the steroid is not soluble in aqueous medium of the blood plasma18. The increase in testosterone observed here would have equally resulted in an increase in serum albumin but this is not so from the results of this study. Serum albumin in conjunction with other clinical indices is used as test to evaluate liver function disorder but in this research it is evaluated in regards to its transport role in connection with testosterone. Baron, documented that three factors may lead to a decrease in plasma total protein and albumin concentrations viz liver damage, impaired intake of protein and toxic destruction of the protein19. It is probable that the third factor may account for the observed decrease of serum albumin and total serum protein in this study. Conclusion We concluded that theobromine administration produce significant increase in serum testosterone and a decrease in serum albumin, total serum protein and transferrin. It also produced histological lesions on the seminiferous tubules. The extent to which structural damage on the seminiferous tubule may affect its function is not known, but on account of the fact that theobromine is a caffeine metabolite responsible for 50% of caffeine toxicity and caffeine intake is a dose dependent risk factor in infertility, feotal loss and growth impairment,1, 20 work is on going in our laboratory to assess the effect of theobromine on spermatogeneis. ACKNOWLEGEMENT The authors wish to acknowledge the funding of this research by the University of Calabar Senate research grant. REFERENCES
© 2005 Nigerian Society for Experimental Biology. The following images related to this document are available:Photo images[bk05017t1.jpg] [bk05017f1.jpg] |
| |||||||||

{kind=link}
{kind=link}