 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biokemistri, Vol. 17, No. 2, Dec, 2005, pp. 171-178 Molecular Identification and Population Dynamics of the major Malaria Vectors in a Rainforest zone of Nigeria Isaac O.OYEWOLE1*,Adejoke C. IBIDAPO2,Adedayo O. ODUOLA3,4, Judith B. OBANSA4,and Samson T. AWOLOLA4 1Department of
Basic and Applied SciencesBabcockUniversity,
Ilishan Remo, Nigeria Received 9 July 2005 Code Number: bk05024 Abstract Adult female mosquito vectors were collected from three villages in a typical rain forest area of Nigeria where no information exists on the major malaria vectors associated with human malaria. Sampling was carried out between January 2004 and January 2005 using pyrethrum and Human landing catch (HLC) techniques. A total catch of 2010 mosquitoes was recorded out of which 1800 were morphologically identified as female Anopheline mosquitoes. Further identification of the Anopheline species using the morphological keys revealed that 1399 (77.7%) belonged to the Anopheles gambiae s.l. and 401 (22.3%) to Anopheles funestus. A PCR based test on the Anopheles gambiae group identified 636 (45.5%) as Anopheles gambiae s.s and 763 (54.5%) as Anopheles arabiensis respectively. The cocktail PCR-assayon the total Anopheles funestus group showed 307 (76.6%), to be Anopheles funestus s.s and 94 (23.4%) to be Anopheles leesoni. The total number of Anopheles gambiae collected across the 3 villages was significantly higher (P<0.05) than the number of Anopheles funestus caught within the same period. However, there was a seasonal difference in the population of Anopheline species collected in which the wet season collections constitute 45.4% An.gambiae s.l and 17.7% An.funestus while the dry season population constitutes 32.3% An.gambiae s.l and 4.5% An.funestus. The dry seasoncollections were predominantly An. arabiensis producing 23.9% of the total catch in.The overall number of Anopheles mosquitoes collected in the wet season was significantly higher than that of the dry season (P<0.01). Generally, low sporozoite rates were recorded in all the communities and this may be an indication that transmission in this area is less intense. This study provides information on mosquito ecology, genetic and molecular techniques for identification of species complexes which are important strategies for planning malaria control programmes. Key words: An. gambiae, An arabiensis, An.funestus, An. lessoni, PCR, ELISA, Nigeria. INTRODUCTION Malaria is a life threatening disease, transmitted by the Anopheles mosquitoes. The disease affects more than 200 million people in the world over. It is the major problem of the developing countries especially those in the tropical Africa where 90 percent of the reported death have been attributed to malaria infection 1,2 . Malaria is a vector-borne human disease and Anopheles mosquitoes are the implicated agents of malaria parasite transmission3. In sub-Sahara Africa, the main malaria vectors that play major roles in transmission of the disease belongs to the group of the Anopheles gambiae Giles and An. funestus Giles. Although other (secondary) vectors such as An. moucheti and An. nili have been reported in some parts of Africa 4. These groups comprise of diverse species whose distribution and composition vary from one geographical location to another being greatly influenced by the prevailing climatic cnoditions5. The An. gambiae complex comprises of seven morphologically indistinguishable species while An. funestus group consists of about nine morphologically identical species especially at the adult stage6, 7,8. Identification of the members within the same morphologically identical taxon to species level is an essential component of epidemiological study of malaria. Since this will help to incriminate the responsible vector species as an important step in formulating strategic control programme. However, cryptic within the taxon are difficult to identify using conventional morphological characters because of the morphological overlap between them. The need to avoid misidentification between species for proper clarification of their epidemiological role in disease transmission has led to the development of molecular assays for species identification 9,10,11,12. The present study is aimed at using molecular assays to identify the major Anopheline species attracted to man in order to provide information on the mosquito population dynamics and malaria transmission in a forest ecotype. MATERIALS AND METHODS Study Area Three communities in Ikenne and Remo north local government areas of Ogun state were selected for the study. These communities lie in the forest area of Nigeria and are about 150km from Lagos. The prevailing tropical climate is characterized with rainy season from April to October (with a short break in July) and dry season between November and March. Population in each community is between 600 and 8,000 whose major occupation is farming. Their dwellings are either traditional houses (partially or totally plastered with mud) often roofed with woven palm leaves or modern houses (partially or totally plastered with cement) roofed with iron sheets. Domestic animals such as goats, dogs, chicken and pigs are commonly sheltered inside the dwellings or in nearby sheds. Akaka is located on the hill, surrounded by valley with slow moving river that runs around the town. Ilara-Remo is located on lowland, it shares boundary with Akaka-Remo, Ilisan-Remo, Ode-Remo and Irolu communities. The only source of water here is through the stream located proximally to Akaka-Remo town but much further from Ilara-Remo itself. Ijesa-isu is a smaller settlement located along Ago Iwoye road the major through fare to the neighbouring towns. Specimen collection and preservation Adult female Anopheline vectors were collected in all the three villages twice a month between January 2004 and 2005. Indoor and outdoor collections were made using pyrethrum and Human Landing Catches (HLC) with the aid of aspirator and searchlight following the standard procedure13. Samples were preserved dry over silica gel in eppendorf tubes prior to identification. Larvae were surveyed to locate the Anopheline breeding sites. Specimen identification Binocular microscope was used for morphological identification with the aid of identification manual 6,7. Molecular Identification Polymerase chain reaction (PCR) technique of Scot et al.12 with the minor modifications as detailed in Van Rensburg et al.14 was used to identify species of An. gambiae while An. funestus Giles was identify to species level using the multiplex PCR technique of Koekemoer et al.15 Elisa Tests Elisa assay was used to investigate the source of the blood meal 16 while the modified method of Wirtz et al.17 was used as earlier outlined by Burkot et al.18 to determine circumsporozoite protein of Plasmodium falciparum, P. Ovale and P. malariae. Statistical analysis Chi-square tests were used to compare the species population, composition, biting activities and seasonal variation in the study communities. RESULTS Morphological identification of Mosquitoes collected A total of 2010 female mosquitoes were collected from the 3 villages within a period of one year and 1800 of these were identified as female anopheline mosquitoes; consisting of 1399 (77.7%) Anopheles gambiae s.l and 401 (22.3%) Anopheles funestus. The remaining 210 mosquitoes caught were mainly culicine. The overall number of Anopheles gambiae caught was significantly higher (χ2 = 120.84; P ±< 0.05; df =1) than the number of Anopheles funestus collected.
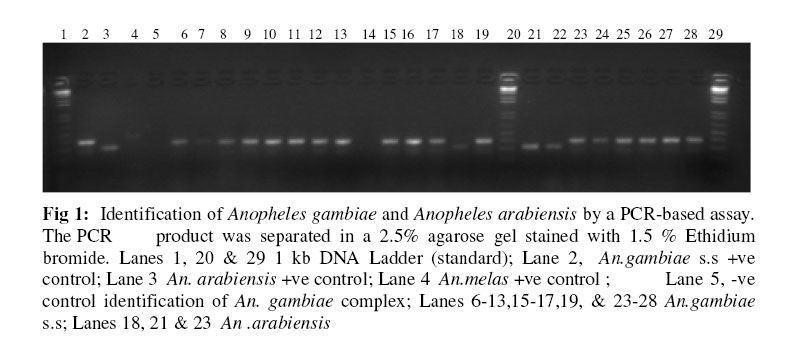
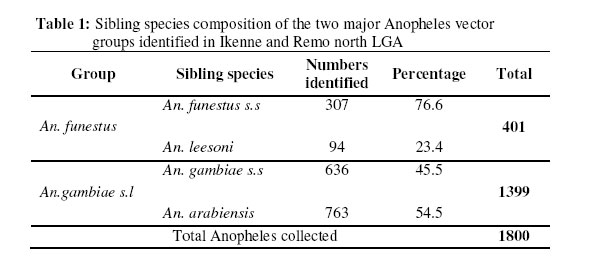
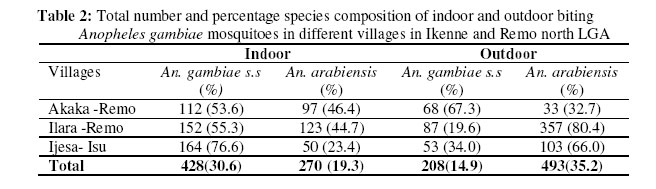
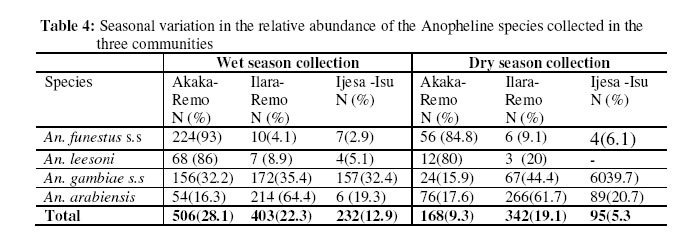
Molecular characterization of sibling species within the two major vector groups Two polymerase chain reaction assays: The Anopheles gambiae species specific PCR and the cocktail PCR assay were employed to distinguish the cryptic species present within the Anopheles gambiae and Anopheles funestus group respectively (Figures 1 & 2). Of the total Anopheles gambiae collected, 636 (45.5%) were identified as Anopheles gambiae s.s. while 763 (54.5%) were Anopheles arabiensis. Also, the groups of Anopheles funestus sampled showed that 307(76.6%) were Anopheles funestus s.s and 94 (23.4%) were Anopheles leesoni (Table 1). The Anopheles arabiensis population was significantly higher than the population of its sympatric counterpart, Anopheles gambiae s.s. (χ2 = 89.1, df =2, P ±< 0.05. Meanwhile, no significant statistical difference was observed between the populations of Anopheles funestus s.s and Anopheles leesoni (χ2 = 3.7, df =2, P > 0.05). Indoor / Outdoor biting activities of the cryptic species The total number of Anopheles gambiae s.s collected both indoors and outdoors was 428 (67.3%) and 208 (32.7%) respectively (Table 2). The observed differences in the proportion of the Anopheles gambiae collected showed that the total number of Anopheles gambiae s.s collected indoors was significantly higher when compared with those collected outdoors. (χ2 = 10.3; df =2, P±< 0.05) and this reached the maximum in July/August. The differences in the indoor and outdoor collections showed that the outdoor population of the Anopheles arabiensis species was significantly higher when compared with the population collected indoors. (χ2 =107.9; df =2, P ±<0.05) and this also reached the peak in June/July. However, the indoor population of Anopheles funestus s.s was not statistically different (χ2 = 0.3, df =2, P > 0.05) from those collected outdoors but higher number of indoor collection was recorded in June. The pre- bed collections showed that the population of An. leesoni was significantly higher when compared with other species (P<0.01). Abundance of the mosquito vector species at the sampling sites The population of the species collected in the three communities as shown in Tables 2 & 3 showed that the proportion of Anopheles gambiae s.l collected in Ilara-Remo community was not significantly different when compared with numbers collected at Akaka-Remo and Ijesa Isu (χ2 = 1.3; df =2, P > 0.05). Also the differences in the population of Anopheles funestus collected in the three sites were not statistically significant ((χ2 = 0.12; df =2, P > 0.05). However, the total number of Anopheles gambiae collected across the 3 villages was significantly higher (P<0.05) than the number of Anopheles funestus caught within the same period. The density of the four species identified in the sampled communities varied according to the season of the year (Table 4) .The wet season collections constitute 45.4% An.gambiae s.l and 17.7% An.funestus while the dry season population constitutes 32.3% An.gambiae s.l and 4.5% An.funestus. The dry seasoncollections were predominantly An. arabiensis producing 23.9% of the total catch in.The overall number of Anopheles mosquitoes collected in the wet season was significantly higher than that of the dry season (P<0.01). ELISA Tests for blood meal sources showed that >70% of An. arabiensis fed on other animals apart from man which was significantly higher than that observed for An.gambiae s.s and An. funestus group (P<0.05). However, An. funestus s.s were predominantly positive for human blood meal. Sporozoite rates showed that only 1.2(2/150) and 1.0(2/80) of An.gambiae s.l and An.funestus group respectively tested positive for ELISA test and this showed no significant difference between the wet and dry season collections (P>0.05). DISCUSSION Previous studies had reported about thirty-three species of Anopheles mosquitoes in Nigeria 19, 6. The major vectors of malaria were members of the An. gambiae s.l and An.funestus group while the secondary vectors were An.nili, An.moucheti and An.rivulorum which also have a wide distribution throughout Nigeria20, 6,7. More recent studies have also documented the species composition, distribution and transmission dynamics of some of the malaria species in southwestern Nigeria21, 22,23. In the present study four major species of malaria were identified. Two of these species, An. arabiensis and An. gambiae s.s which were members of An.gambiae s.l occurred in sympatry at Ilara-Remo, Idagolu and Ijes-isu. The sympatric occurrence of these species had been earlier reported7. These two species were also found to dominate the total collections in the study communities. The abundance of these species had been reported in the southwestern Nigeria and in humid forest zones elsewhere24, 5,4,23. This has been found to be connected with certain climatic factors most especially the total annual precipitation which appears to influence the range and relative abundance of these species5. An. arabiensis dominate the dry season collections while An. gambiae s.s was more abundant during the wet season. The seasonal variation in number and composition of species which occurred in sympatry had been reported25. Members of the An.funestus group identified in this study were An.funestus s.s and An. leesoni while those reported in Africa so far include An.rivulorum Lesson, An. parensis Gillies, An. funestus Giles and An.leesoni which all occurred in sympatry6, 7,26. Most of these studies only implicated An.funestus as a major vector except that of Wilkes et al.27 which showed An. rivulorum as one of the minor vectors. In the present study, we observed that An. leesoni were predominant in outdoor collections which probably supports the earlier findings where An. leesoni was being described as an exophilic and zoophilic species 28,29,23. Minimal seasonal variations in population, distribution and composition of Anopheline mosquitoes were observed in all the communities. The wet season collection was almost three times higher than that of the dry season. Large number of An.funestus species collected in Akaka could be due to the location of the village and its proximity to a slow–moving stream. The breeding site of these species was confirmed from the results obtained from the larvae collected from the edge of the stream, where most of the emerged adults (70%) were An. funestus. Previous studies had also reported a predominant number of An, funestus in villages located in high altitude with proximity to slow-moving stream30. Although in the present study, there was a drastic reduction in the dry season population of these species which coincided with reduction in the volume of the stream and disappearance of temporary pool of water around the bank. This observation may not agree with the earlier reports that An.funestus adapts to a variety of habitat6,7, or are more common in the dry season than in the wet season31. The results of the blood meal analysis also agreed with the previous reports which showed that An. gambiae and An. funestus s.s were highly anthropophagic30. In the present study a large number of these species were collected from the human baits. An arabiensis Patton and An. leesoni were both zoophagic since the blood meal analysis showed an active feeding on cattle outside the house 32,33. Low sporozoite rates observed may indicate low intensity of malaria transmission in this area 30. The correct identification of the vector species of Anopheline mosquitoes, their abundance, distribution and composition as well as the malaria transmission dynamics in different localities is germane to multidirectional approach required to roll back malaria in Nigeria. Acknowledgements We gratefully acknowledged the efforts of our field assistants during the field activities. This study was partly supported by the grant MIM project A30026 through the UNICEF/UNDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases (TDR) awarded to S.T.A. REFERENCES
© 2005 Nigerian Society for Experimental Biology. The following images related to this document are available:Photo images[bk05024f2.jpg] [bk05024f1.jpg] [bk05024t4.jpg] [bk05024t2.jpg] [bk05024t1.jpg] [bk05024t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}