 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
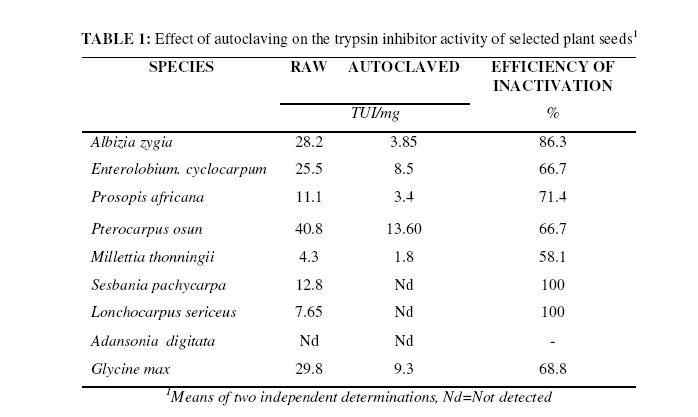
Biokemistri, Vol. 18, No. 1, June, 2006, pp. 21-24 Efficiency of inactivation of trypsin inhibitory activity in some selected tropical plant seeds by autoclaving Ikechukwu E. EZEAGU Nutrition Unit, Department of Medical Biochemistry, University of Nigeria, EnuguCampus, Nigeria Email: ikezeagu@yahoo.co.uk , Tel: 0803-3949541 Received February 24, 2005 Code Number: bk06004 Abstract Trypsin inhibitor (TI) levels in the crop seeds varied between 0.0 in Adansonia digitata and 40.8 TIU/mg in Pterocarpus osun. Efficiency of inactivation of TI by autoclaving ranged from 58.1% in Millettia thonningii to 100% in Sesbania pachycarpa and Lonchocarpus. sericeus. It is concluded that the effect of heat treatment on TI varies with plant species and could be dependent on the structural formula of each TI. Keywords:autoclaving, efficiency, trypsin inhibitor, tropical, seeds INTRODUCTION The shortages and current high prices of traditional staples have stimulated the search for newer and unconventional sources. On going research in this laboratory is exploring the possibility of nutritional exploitation of lesser-known biomass as alternatives in livestock feeding so as to spare more of the conventional food articles for human consumption (1). Previous studies have indicated that quite a large number of unconventional plant seeds could be good sources of nutrients and may have the potential of broadening the present narrow food base in the tropical regions if given research attention (2,3,4). However, occurrence of antinutrients in plant food is a notable disadvantage for their utilization if unprocessed (5). Consequently, plant foods have been traditionally processed for human consumption in various ways such that toxicity is largely eliminated. For example, phytohaemagglutinins and trypsin inhibitors (TIs) are heat labile and as such are readily destroyed by treatment of plant material with moist heat or autoclaving, dry heat being less effective. Levels of oxalates, tannin, nitrate/nitrite and various other antinutrients in food may be reduced by soaking and/or boiling in water, toasting and by fermentation (6,7,8). This study seeks to determine the efficiency of autoclaving process in the inactivation of trypsin inhibitory activity in some crop seeds which have been previously described4,9. Materials and Methods Treatment of Samples About 1kg of the matured fruits were procured from IITA-ICRAF Arboretum, International Institute for Tropical Agriculture, Ibaden, Nigeria. The seeds were manually separated from the pods. A soy bean sample (TGX 1660-15F), obtained from IITA, Ibadan, was included for comparison.Samples (500g each) were ground to flour using a Wiley Mill with the 1 mm mesh sieve and stored in plastic bags at –4 oC until analysis. About 200g of each seed flour was put in small beakers and subjected to autoclaving for 20 minutes under steam pressure of 1kg/cm2 at a temperature of 105 o C. The flours were cooled and dried at 30 o C in the oven Determination of trypsin inhibitor activity Trypsin inhibitor (TI) activity was determined by the method of Kakade et al. (10) using benzoyl-DL-arginine-p-nitroanilide (BAPNA) as substrate. 1.0g of samples was extracted with 50ml of distilled water for 3 hrs with shaking. 2.0ml diluted (1:33) extract was mixed with trypsin solution and incubated at 37 oC for 10mins, followed by the addition of 2.0ml substrate solution. Exactly 10mins after, the reaction was stopped by injecting 1.0ml of 30% acetic acid solution and corresponding blanks (acetic acid added before enzyme) were run concurrently. The mixture was filtered through a Whatman No. 2 paper and the absorbance read at 410nm. The reaction was also run in the absence of inhibitors by replacing the sample with 2.0ml distilled water. The results were expressed as the number of trypsin units inhibited (TUI) per milligram of dry sample. One trypsin unit (TU) was arbitrarily defined as an increase of 0.01 absorbance units at 410nm in 10mins for 10ml of reaction mixture under conditions described. Results and Discussion Table 1 compares the levels of TI both in the raw and autoclaved seed samples. Adansonia digitata seeds exhibited no TI activity, which is a nutritional advantage. Highest level of TI (40.8 TIU/mg) occurred in Pterocarpus osun. Low levels occurred in Millettia thonningii and Lonchocarpus sericeus (4.3 and 7.65 TIU/mg respectively). While the levels of TI activities in Albizia zygia (28.2 TIU/mg) and Enterolobium cyclocarpum (25.5 TIU/mg) and soybean (Glycine max) (29.8 TIU/mg) were comparable, the efficiency of inactivation differed. TI activities were totally eliminated only in S. pachycarpa and L. sericeus seeds while, in others, only percentages of the initial levels were destroyed. In P. osun, 66.7% TI were eliminated leaving the residual level still high (13.6 TIU/mg). Marickar and Pattabiraman11 reported over 95% of activity destroyed in 15 minutes of 100 oC heat treatment, while Ramamani et al.12recorded an efficiency of about 95% by roasting in soybean While most literature dealing with protease inhibitors indicate that they are heat labile, some workers have contradicting reports. Nitsan13, testing field bean, found that growth rate and weight gain per unit of food eaten where both reduced in rats fed autoclaved field beans and concluded that trypsin and chymotrypsin inhibitors were not destroyed by heat treatment, and thus confirming an earlier report of Sohonie and Ambe14. Tuber et al.15 reported that crystalline trypsin inhibitor from lima beans is exceptionally heat stable in contrast to the crystalline inhibitor which Kunitz16 isolated from soybean which is completely destroyed on short atmospheric boiling. In general, purified trypsin inhibitors may be resistant to heat17,18. Since trypsin inhibitor is a biologically active protein molecule, heat may change its native conformation, depending on microenvironment of the molecule. Prior hydration by soaking in water for 12-24 hours may make heat treatment more effective. It should however be noted that excessive heat treatment can, of course, reduce the nutritive value of these seeds owing to destruction of amino acids19,20. References
© 2006 Nigerian Society for Experimental Biology. The following images related to this document are available:Photo images[bk06004t1.jpg] |
| |||||||||

{kind=link}