 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biokemistri, Vol. 18, No. 1, June, 2006, pp. 53-61 Behavior of manganese ion in basic medium: consequence for the measurement of enzymatic activities using nitrophenyl derivatives Sébastien NIAMKE1,3*,Lucien Patrice KOUAME2, Edmond Ahipoh DUE2and Soumaila DABONNE3 1.
Laboratoire de Biotechnologies, Unité de Formation et de Recherche en
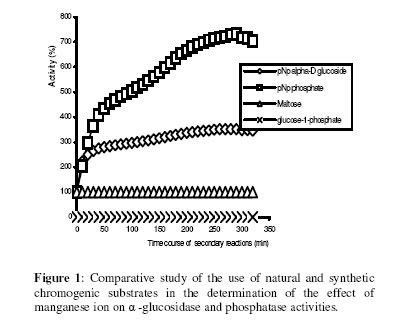
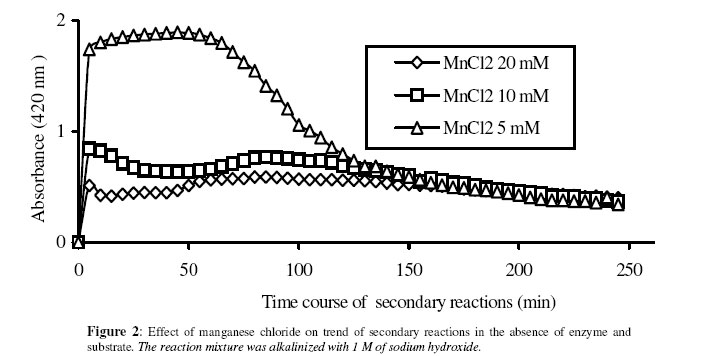
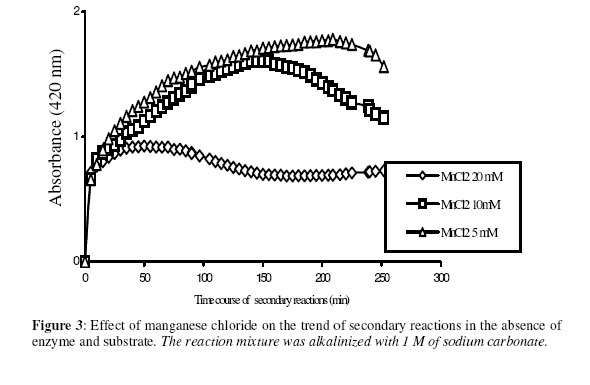
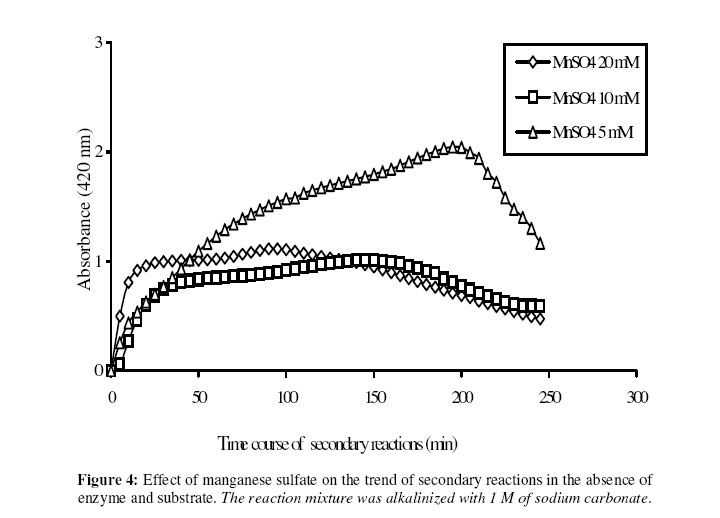
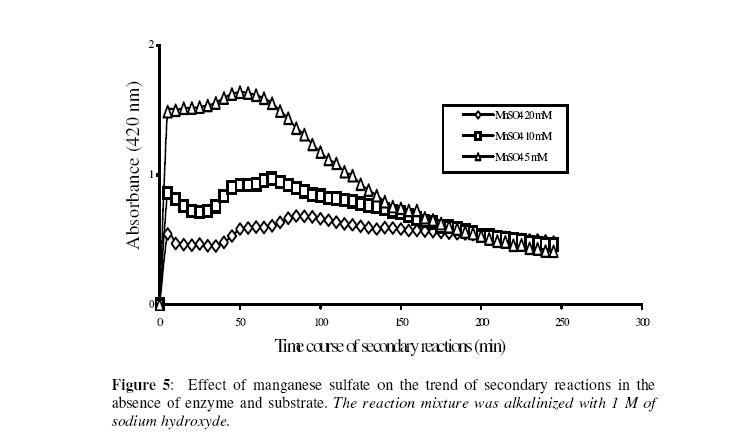
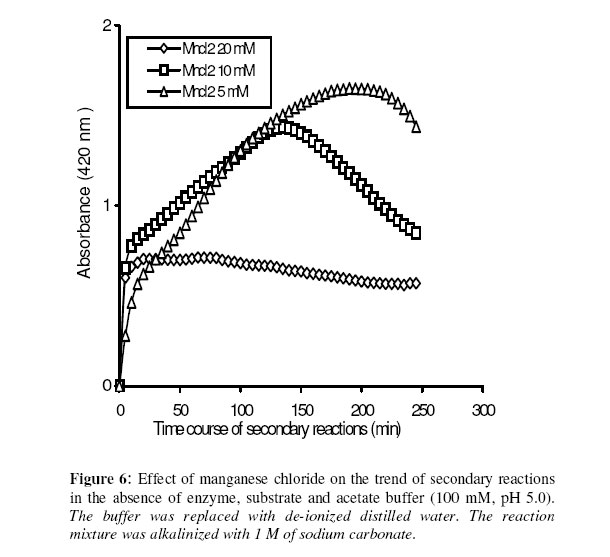
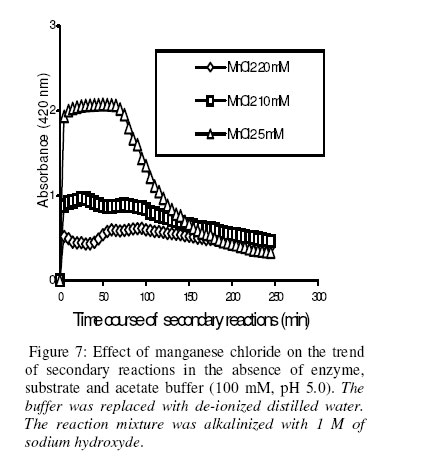
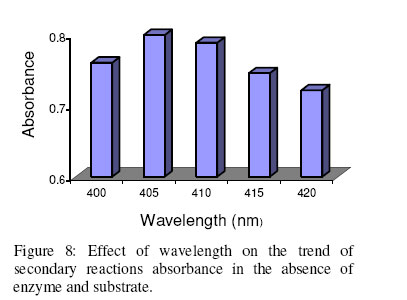
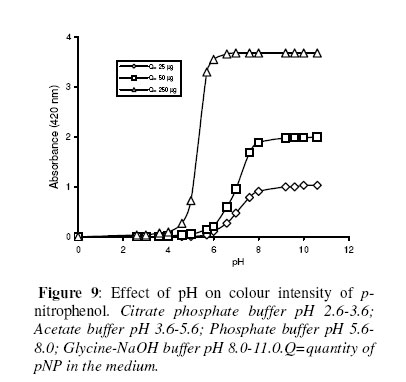
Biosciences de l’Université de Cocody-Abidjan, 22 BP 582 Abidjan 22, Côte d’Ivoire. Received January 25, 2006 Code Number: bk06009Abstract The use of nitrophenyl derivatives in the determination of the influence of manganese ion on catalytic activities of glycosidases and phosphatases remains a problem. In fact, in this study, adding manganese chloride or manganese sulfate to sodium hydroxyde or sodium carbonate in aqueous solution yielded secondary reactions. These secondary reactions affected the absorbance for wavelengths varying between 400-420 nm. This chemical behavior did not allow reliable results in determining the influence of manganese ion on glycosidase and phosphatase activities, when nitrophenyl derivatives were used as substrates. This study showed that this problem could be avoided when natural substrates such as disaccharides (glycosidases) and phosphorylglycosides (phosphatases) were used in enzymatic tests. Keywords:phosphatase, glycosidase, manganese ion, nitrophenyl derivatives, basic medium, enzymatic determination. INTRODUCTION To assay exoglycosidase and phosphatase activities, researchers generally use synthetic chromophoric substrates, such as (o- or p-) nitrophenyl-phosphate and (o- or p-) nitrophenyl-glycoside1-3. The enzymatic hydrolysis is stopped by adding sodium carbonate or sodium hydroxide to the reaction mixture. The (p- or o-) nitrophenol generated, becomes yellow. The intensity of the colour is determined with a spectrophotometer at wavelengths ranging between 400 and 420 nm4-7. This quantification method of glycosidase and phosphatase activities allows determination the influence of chemicals on enzymatic activities: first, the enzyme is incubated with the chemical for a while, and then the substrate is added to this medium to quantify the enzymatic activity. The reaction is stopped as previously. Among chemicals having their influences studied on aforementioned enzyme catalytic activities, manganese chloride and manganese sulfate are generally listed8-9. These two compounds generate a manganese ion in the medium. Studying the influence of catalytic activities of two purifiedglycosidases oftermite Macrotermes subhyalinusunder the same conditions using p-nitrophenyl-α-D-glucopyranoside as substrate, Kouamé10 noticed a change in the reaction mixture after addition of sodium carbonate (2%, w/v) to stop the reaction. In order to understand the chemical behavior, and to identify the products responsible for the secondary reactions initiated by this ion on catalytic activities of enzymes hydrolyzing nitrophenyl derivatives, this study was undertaken. MATERIALS AND METHODS Chemicals and enzymes Chromogenic substrates (p-nitrophenyl- α-D-glucopyranoside and p-nitrophenyl-phosphate) and maltose were obtained from Sigma-Aldrich and glucose 6-phosphate, from Merck. All other chemicals and reagents were of analytical grade. The enzymes (α-glucosidase and phosphatase) used in this work were from cockroach (Periplaneta americana). This insect was collected in the campus of the University of Cocody-Abidjan (Côte d’Ivoire), and the enzymes were purified according to the method of Kouamé11. Enzyme assays p-Nitrophenyl-α-D-glucosidase and p-nitrophenyl-phosphatase activities were measured at 25°C for 10 min in reaction mixture containing 250 µl of 100 mM acetate buffer (pH 5.0), 75 µl of p-nitrophenyl-α-D-glucopyranoside (or p-nitrophenyl-phosphate) (5 mM) and 50 µl of enzyme preparation. The reaction was stopped by adding 2 ml of sodium carbonate (1 M). The release of p-nitrophenoxide anion (pNP) was quantified at 420 nm using a spectrophotometer (Spectronic Genesis 5). The calibration curves (pNP) were carried out under the same conditions of pH and temperature7. Maltase activity was measured under the same conditions as previously, but with a reaction mixture containing 75 µl of maltose (33.4 mM). The reaction was stopped by heating the medium at 100°C for 5 min. The glucose released was determined by the glucose oxidase-peroxidase method12. The peroxidase activity was measured at 436 nm. Glucose-6-phosphatase activity was measured as described previously, using 75 µl of glucose-6-phosphate (33.4 mM). The reaction was stopped by adding to the reaction mixture 2 ml of trichloacetic acid 2 % (w/v). The phosphate released was quantified by the method of Taussky and Shorr13, using molybdate and vanadate. Phosphatase activity was measured at 410 nm. Effect of manganese ion on α-glucosidase and phosphatase activities The effect of manganese ion on α-glucosidase activities of cockroach periplaneta americana toward synthetic chromogenic substrate (p-nitrophenyl-α-D-glucopyranoside, 5 mM), and natural substrate (maltose, 33.4 mM) was determined at 25°C by pre-incubation of 50 µl of enzyme preparation with manganese ion in 100 mM acetate buffer (pH 5.0) for 10 min. Then, 75 µl of substrate was added to the reaction mixture (final volume of 250 µl). The incubation was performed at 25°C for 10 min, and the products released in each case by α-glucosidase activity were determined as previously. The measurement of the absorbance was performed using a spectrophotometer just after addition of sodium hydroxide (1 M) or sodium carbonate (1 M) for p-nitrophenyl-glucosidase activity, and 30 min after addition of glucose oxidase-peroxidase preparation for phosphatase activity. Concerning the effect of manganese ion on phosphatase activities of cockroachperiplaneta americana, synthetic chromogenic substrate (p-nitrophenyl-phosphate, 5 mM) and natural substrate (glucose-6-phosphate, 33.4 mM) were used, and activities were measured as described previously. Absorbances were measured with a spectrophotometer just after addition of sodium hydroxide (1 M) or sodium carbonate (1 M) for p- nitrophenyl-phosphatase activity, and after addition of molybdate and vanadate for glucose-6-phosphatase activity. Effect of manganese ion on the trend of secondary reactions in the absence of enzyme and substrate In a spectrophotometer cell, it was successively added 125 µl of 100 mM acetate buffer (pH 5.0); prepared in de-ionized distilled water, 125 µl of manganese chloride or manganese sulfate (5, 10 and 20 mM) and 2 ml of sodium carbonate (1 M) or sodium hydroxide (1 M). The absorbance was measured with a spectrophotometer at 420 nm. Are manganese chloride, manganese sulfate, sodium hydroxide and sodium carbonate, taken individually, able to change the absorbance at 420 nm? In order to answer this question, 2 ml of each of these chemical compounds were introduced in a spectrophotometer cell (Spectronic Genesic 5). Acetate buffer (100 mM, pH 5.0) was used as reference. The absorbances were also measured with the same spectrophotometer at 420 nm. Effect of wavelength on the absorbance measurement of secondary reactions in the absence of enzyme and substrate The same reaction mixture and conditions as those used in the study of the influence of manganese ion on the trend of secondary reactions in the absence of enzyme and substrate were used for this study. Wavelengths ranging from 400 to 420 nm were studied. The reaction time was 15 min. Effect of pH on the intensity of p-nitrophenol colour A defined amount (250 µg, 50 µg and 25 µg) of p-nitrophenol is dissolved in 2 ml of 100 mM acetate buffer (pH 2.6 to 11.0). The intensity of p-nitrophenol colour was determined at 420 nm with a spectrophotometer (Spectronic Genesis 5) against a reference (containing no nitrophenol). RESULTS AND DISCUSSION The use of natural substrates as maltose and glucose-6-phosphate in the determination of the effect of manganese ion on catalytic activities of α-glucosidase and phosphatase of cockroach Periplaneta americanashowed that this ion inhibited totally the phosphatase activity, while it had no effect on α-glucosidase (Figure 1). This result remained as such for more than 5 hours after the first absorbance readings, and addition of glucose oxidase-peroxidase solution (α-glucosidase) or vanadate-molybdate reagent (phosphatase). This was expressed by a constante percentage for the activity for more than 5 hours (Figure 1). When synthetic chromogenic substrates such as p-nitrophenyl-phosphate and p-nitrophenyl-α-D-glucopyranoside were used in the place of the above mentioned natural substrates, the determination of the effect of this ion on these enzymes remained a problem. In fact, after incubation of the reaction mixture, followed by the addition of sodium carbonate to stop the enzymatic reaction, a rapid increase of absorbance with time was observed at 420 nm after the first reading. Curves obtained from chemical behavior as a function of time were difficult to interpret (Figure 1). In the first 10 minutes, we obtained an increasing percentage for the activity, varying from 100-200, showing that manganese ion led to an activation effect on α-glucosidase and phosphatase activities. These results are the opposite of those obtained with natural substrates and suggest a direct participation of secondary reactions. In order to learn about the products responsible for this chemical behavior, many tests were performed. To check the involvement or not of the substrate (p-nitrophenyl-phosphate or p- nitrophenyl-α-D-glucopyranoside) and the enzyme (phosphatase or α-glucosidase) in the initiation and the trend of secondary reactions, they were not added to the reaction mixture. They were replaced by acetate buffer (100 mM, pH 5.0). The alkalinization of the medium has been realized either with 1 M of sodium hydroxide or sodium carbonate (Figures 2 and 3). In the two cases of alkalinization, we noticed that secondary reactions increased with time. This means that substrate and enzyme were neither the initiating components nor responsible for this chemical behavior. However, it was showed in all cases that, the lower the manganese chloride concentration in the reaction mixture, the higher the absorbance values (Figures 2 and 3). When the medium became more basic, for instance, by adding 1 M of sodium carbonate, the two curves obtained from absorbances of reaction mixtures containing respectively 5 and 10 mM of manganese chloride showed a similar trend (Figure 3). These curves were bell-type with an ascending portion extended over many hours. After reaching maximum absorbances of reaction mixtures, these absorbances quickly fell down. The maximal absorbances obtained with the two curves (i.e. with 5 mM and 10 mM of manganese chloride) were 1.8 (for 5 mM) and 1.6 (for 10 mM) at 220 min and 150 min of reaction, respectively. For the reaction mixture containing 20 mM of manganese chloride, a different trend of curve was obtained comparatively to those described above (Figure 3). Absorbances rapidly increased during the first 10 min of reaction and the maximum absorbance was observed at the 40 th minute with a value of 0.9. Then it stabilized for 20 min; after which, we observed a slow decrease and then an increase. Absorbances obtained were lower than those obtained with reaction mixtures containing 5 mM and 10 mM of manganese chloride (Figure 3). When the alkalinization of the medium is realized with 1 M of sodium hydroxide, the reactions increased faster than those alkalinized with 1 M sodium carbonate. Maximum absorbances were quickly reached. When the medium concentration in manganese chloride was 5 mM, a bell-type curve was obtained as previously (alkalinization of the medium with 1 M sodium carbonate). The maximum absorbance was 1.9. It was reached at the 60 th min (Figure 2). However, when the reaction mixture concentrations were 10 mM and 20 mM, the curves had the same trend. They were characterized by two peaks. The curve obtained from the reaction mixture containing 20 mM of manganese chloride showed its first maximum absorbance at the 10th minute and the second at 70th minute; the same as the reaction mixture containing 10 mM of manganese chloride. Above 150th min, all absorbances were more or less identical. When manganese chloride was replaced by manganese sulfate, and the experiment was conducted under the same conditions, a change of absorbance was noticed. In each case of alkalinization of the reactional medium (1 M sodium hydroxide or 1 M sodium carbonate), more or less the same trend was obtained (Figures 4 and 5). These results suggested that manganese ion, sodium hydroxide, sodium carbonate and acetate buffer were probably responsible for the initiation and the increase of these secondary reactions. When acetate buffer (100 mM, pH 5.0) was replaced by de-ionized distilled water (Figure 6), an increase of absorbance was observed. The curves obtained were similar to those of figures 4 and 5. These results allowed us to infer that compounds favouring the triggering and the trend of secondary reactions might be manganese ion and sodium hydroxide or carbonate. Acetate buffer (100 mM, pH 5.0) was not involved in the initiation and the trend of these reactions. The absorbances of manganese chloride, manganese sulfate, sodium carbonate and sodium hydroxide solutions were separately measured at 420 nm. No change of absorbance was noticed in each case; meaning that each compound, taken individually in an aqueous medium, was not able to initiate or to trigger the secondary reactions. However, when manganese ion was added to a basic medium, these reactions were yielded (Figure 7). A trend of absorbances was observed regardless of wavelengths chosen between 400-420 nm. After 15 min of reaction, the highest absorbance was observed at 405 nm (Figure 8). This result indicated that the absorbances at 420 nm were under-estimated compared to what they should be if they were measured at 405 nm. To avoid the initiation of these secondary reactions, the reaction mixture should not be alkalinized with sodium hydroxide or sodium carbonate. The enzymatic reaction containing manganese ion should be stopped by using other methods. One of the classical means is the heating of the reaction mixture. However, the heat treatment of the reaction mixture presents two major drawbacks: a possible hydrolysis of the substrate and the change of the maximum wavelength of p-nitrophenol7. To avoid such situations, the other alternative was to measure the absorbances at defined periods of time, without heating the reaction mixture, and by using either ELISA plate reader or spectrophotometers equipped with a regulated water bath. However, when the reaction mixture was not alkalinized and the quantity of p-nitrophenol was low in the medium, at pH 5.0, the absorbances were also low (Figure 9). This situation is therefore not helpful if we have an enzyme which is inhibited by manganese ion. As a matter of fact, a slight inhibition could be considered as a strong inhibition because the enzymatic activity was low. Above pH 6.0, the absorbances increased rapidly to be more or less stabilized at pH 8.0. However, when p-nitrophenol quantity was high in the reaction mixture, at pH above 4.0, absorbances were high (Figure 9). Another alkalinization of the reaction mixture was not necessary. Itt is the case with enzymes which have their optimum pH in alkaline medium. The major problem encountered concerns glycosidases where the majority of enzymes are acidic biocatalysers. To solve these problems, the use of natural substrates (disaccharides for glycosidases and phosphorylglycosides for phosphatases) was necessary and important in the study of the influence of manganese ion on the glycosidase and phosphatase catalytic activities. CONCLUSION The study showed that when a manganese ion was in basic medium, it induced reactions that increased absorbances at wavelengths between 400 and 420 nm. This chemical behavior greatly interfered with the influence of this ion on the catalytic activities of enzymes hydrolyzing nitrophenyl derivatives. Thus, these secondary reactions allowed the manganese ion to be a strong activator for glycosidases and monoesterases. To avoid this unfortunate situation, the use of natural substrates is advised in this study. ACKNOWLEDGMENT The authors are grateful to Professor Bernard Colas of the University of Nantes (France), Unité de Recherche sur la Biocatalyse, CNRS-UMR 6204, for his assistance. REFERENCES
© 2006 Nigerian Society for Experimental Biology. The following images related to this document are available:Photo images[bk06009f4.jpg] [bk06009f9.jpg] [bk06009f5.jpg] [bk06009f7.jpg] [bk06009f2.jpg] [bk06009f3.jpg] [bk06009f6.jpg] [bk06009f1.jpg] [bk06009f8.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}